
Abstract
During the Formative period by the Late-Holocene (ca. 3000–1500 BP), semi-sedentary and sedentary human occupations had emerged in the oases, salares, and riverine systems in the central depression (2400–1000 masl) of the Atacama Desert, northern Chile (19–25°S). This hyperarid core was marginally occupied during the post-Pleistocene and middle Holocene droughts. Settlement on these lower belts was accompanied by a rise in humidity, the introduction of Andean crops, flourishment of Prosopis spp. (algarrobo) forests, and increasing integration of domestic camelid caravans. Here, we explore lowland husbandry within risk-spreading strategies, focusing on silvopastoralism and endozoochory between camelids and algarrobos. Analysis of camelid coprolites from seven archeological sites located in the Pampa del Tamarugal, Loa River, and Salar de Atacama found intense grinding from camelid chewing and indicated a ruminal digestive system. Abundant macro and microremains in the form of tissues, phytoliths, crystals, cell structures, and others, were identified as Prosopis, Atriplex, Schoenoplectus, Distichlis, and Phragmites. We conclude that camelids were foraging for Prosopis, although the rather low number of entire seeds preserved in the coprolites leads us to think that these herbivores might not have been the main vectors for the spread and germination of algarrobos. More samples and interdisciplinary studies are needed to comprehend the complex socioecological web in the shaping of these forests and the management of the Atacama Desert landscapes.
Introduction
The Atacama Desert in northern Chile (19–25°S) is an extreme environment where people have lived since at least 13,000 BP. This deep history has been composed of several cycles of humidity and aridity, as well as different sociocultural solutions to learning and managing the diversity of these evolving landscapes (Gayo et al., 2019; Grosjean et al., 2005; Lima et al., 2016; Santoro et al., 2017; Uribe et al., 2020). Occupying a narrow strip of land to the west of the Andes that collides with the Pacific Ocean for thousands of kilometers, this continental territory has an average width of 180 km and an altitudinal difference ranging from 0 to 6000 m above sea level (masl). Its low humidity is caused by the Pacific Subtropical Anticyclone, cold Humboldt Current, and the Andes mountains that block the advection of humid air from the Amazon basin, despite some rainfall passing through in the summer months, nourishing streams and rivers, as well as bringing occasional flash floods. Moreover, the intense solar radiation and high evaporation rates affect the distribution of plant species and vegetation cover. This vegetation exhibits low diversity and varies across different ecological belts such as the puna (±6000–4500 masl), pre-puna (±4500–2400 masl), central depression (±2400–1000 masl), coastal cordillera (±1000–0 masl), and littoral zone (±0 masl). Nevertheless, traditional ecological knowledge shows a broader spectrum of categories, based on first-hand experience (García et al., 2020).
The central depression is characterized by extreme hyperarid conditions as local rainfall rarely occurs (<10 mm/year), and the availability of superficial water is spatially discontinuous. This landscape is referred to as “absolute desert,” where vegetation cover is practically non-existent (Arroyo et al., 1988) and thus bio productivity and resource predictability are low (Grosjean et al., 2007). Still, the localized outcropping of groundwater or discharge of surface runoff over the area has given rise to discrete drainage systems, such as the endorheic basins of Pampa del Tamarugal and Salar de Atacama, as well as the lower and middle Loa River. Here, azonal formations, including salares, oases, and riverine vegetation host flora comprising Fabaceae trees of the genus Prosopis belonging to Algarobia, algarrobos (P. alba Gris., P. flexuosa DC., P. chilensis Mol., P. nigra Gris.) at least from 3000 BP (McRostie et al., 2017), and Strombocarpa sections (Prosopis tamarugo Phil.), Geoffroea decorticans (Gillies ex Hook. and Arn.) Burkart, together with shrubs such as Atriplex spp., Thessaria absinthoides (Hook. & Arn.) DC. and a dense herbaceous cover of Distichlis spicata (L.) Greene. Surface and groundwater discharge also supports the growth of green algae and marsh species such as Juncus sp., Schoenoplectus spp., Baccharis juncea (Lehm.), Phragmites australis (Cav.) Trin. ex Steud., and Sarcocornia fruticosa (L.) A. J. Scott. At this low altitude, grasslands have a less diverse composition than at higher altitudes (Latorre et al., 2002; Villagrán and Castro, 1997).
Given the productivity of the Humboldt Current along the north-south coastal and marine corridor of the littoral zone, the Pacific coast is an immeasurable “larder of protein” and seabird guano (Bittmann, 1984; Santana-Sagredo et al., 2021). Furthermore, minerals and pigments were, and still are, obtained from these “desert landscapes”, which are also the location for the oldest known pigment mine, dated to around 12,000 BP (Salazar et al., 2010, 2013). Following the long-term exploitation and use of copper, silver, gold, and manganese (Sepúlveda et al., 2019), it is no surprise that the Inka state’s interest in these latitudes derived primarily from these ores.
The Atacama Desert has sustained a growing population to the present day, but socioecological problems have been exacerbated on account of it having been a mining, energy, and marine source for the Chilean and global economy since the 19th century (Gayo et al., 2019). Therefore, human interaction with this hyperarid landscape remains a challenge that we must assess using an interdisciplinary and transdisciplinary approach and across a much larger chronological scale.
Risk avoidance or mitigation strategies that traditional societies have used might be observed in more detail in the face of global and local challenges. These run counter to the concentration of resources and monoproduction associated with modern extractivism (Romero-Toledo, 2019). Tactics such as social interdigitation, diversification of productive techniques, broad spectrum economies, or household level cooperatives are considered important in the economic organization of dryland producers from the Andes (Browman, 1987a, 1987b, 1987c; Castro et al., 1984; Hidalgo, 2004; Martínez, 1998; Murra, 1985; Olivera, 1998; Sillar, 2000; Tapia, 2000). These social mechanisms are deployed to reduce the impact of unpredictability and uncertainty in resource availability, thus cushioning their effects (Bollig and Göbel, 1997; Halstead and O'Shea, 1989). Pastoralists employed a range of risk-sharing techniques, such as seasonal shifting, enhancing wild food productivity, dispersing herds over large areas to take advantage of local patch types, multispecies diversification, or increasing herd size (Browman, 1974, 1987c; Dillehay, 2013). Water management and the use of manure of both marine and terrestrial origins were key to expanding the patchy green areas (Hidalgo, 2009; Hidalgo et al., 2019; Santana-Sagredo et al., 2021; Santoro et al., 1998), and agroforestry or silvopastoralism could provide a fertile cycle of nutrients to support desert life within a highly dynamic socioecological context.
Silvopastoralism and Prosopis forests in the Atacama
Camelid domestication and grazing practices precede the introduction and adoption of domesticated plants. “Domesticated camelids facilitated a new way of life that brought complexity and adaptive improvements, creating the necessary conditions for neolithization in the South-Central Andes” (Núñez and Santoro, 2011: 495).
Andean camelids, such as the llama (Lama glama Linnaeus) and alpaca (Vicugna pacos Linnaeus) would have been domesticated from their wild relatives, guanacos (Lama guanicoe Müller) and vicuñas (Vicugna vicugna Molina). The ethologies of these camelids differ in some aspects. The guanaco is a browser and grazer, tolerant of arid conditions; hence, it has greater habitat flexibility (0–4250 masl) and the widest distribution in South America. Groups of guanacos are still seen around the Salar de Atacama and coastal lomas. The vicuña is mainly a grazer and restricted to higher altitudes (3700–4800 masl). Both guanaco and vicuña were hunted for their meat, fat, and hides. The alpaca has been bred primarily as a fine-fiber producer, whilst the llama is more versatile, not only as a source of food, hide, and fiber, but also as a pack animal (Mengoni and Yacobaccio, 2006).
Based on morphological, osteometric, lanimetric, pathological analyses, as well as age profiles and other indirect lines of evidence such as corrals and rock art, it has been proposed that this domestication occurred during the Late Archaic (ca. 7500–4000 BP) period by hunter-gatherers living mainly in the highlands or pre-puna (4500–2400 masl), where vegetation and climate are more suited to South American camelid biotypes (Berenguer, 1996; Cartajena et al., 2007; Dransart, 2002; Gallardo and Yacobaccio, 2005; Núñez, 1981; Núñez et al., 1999; Núñez, 1986; Yacobaccio, 2021; Yacobaccio and Vilá, 2016).
During the Late-Holocene, wetter conditions were coeval with what has been recognized as the local Formative period, when caravans likely began ca. 3000–2000 BP (Berenguer, 2004; Briones et al., 2005; Núñez and Briones, 2022; Pimentel et al., 2011). Water supply favored human occupation of endorheic basins and riverbanks within the central depression or hyperarid core (Santoro et al., 2017; Segura et al., 2022). Legume forests of algarrobo are ubiquitous and abundant in these rising settlements. These have been recovered in the form of stumps, charcoal, wood, thorns, leaves, pods, seeds, and endocarps, and although scant attention has been paid to their historical ecology (Bessega et al., 2021; McRostie, 2014; McRostie et al., 2017; Palacios and Brizuela, 2005), there is a close correlation between human settlements, caravans, and the flourishing of Algarobia patches during this period (Adán and Urbina, 2007; Agüero and Uribe, 2011; Benavente, 1982; García et al., 2014; Llagostera et al., 1984; McRostie et al., 2019; Rivera and Dodd, 2013; Uribe et al., 2020).
So far, research of these lowland sites has focused mainly on agricultural-plant problematics, leaving unexplored the complex socioecological dynamics between animals, forests, and humans. Neither the corrals nor retaining walls in watered areas nor coprolites have been evaluated in detail. Modern local silvopastoral studies do not provide enough information about these relationships, as most of these food webs are either blurred or oriented toward P. tamarugo or European taxa (goats, cattle, alfalfa, grasses, etc.) (Campos et al., 2011). Silvopastoralism systems refer to the active role that a given tree species plays in animal production, by providing tree fodder, refuge, or shade. Still, these systems could involve different management intensities, combinations of the tree/shrub component, or other types of agroforestry such as intercropping (Nair, 1993). Also, particularly the endozoochory of Prosopis might regenerate these ecosystems; given that algarrobo seeds present waterproof seminal cover, requiring scarification to germinate, being related to animal dispersal outside the mother plant range (Campos and Velez, 2015).
Hence, by analyzing camelid dung recovered in Formative residential sites within Pampa del Tamarugal, Salar de Atacama and river Loa basins (800–2500 masl), we aim to evaluate whether silvopastoralism and endozoochary of algarrobo trees occurred, considering traditional risk-spreading strategies in arid environments, which probably nurtured and reproduced these Formative and pre-Columbian ecosystems.
Methodology
We analyzed 34 pellets recovered from seven archeological residential Formative sites in different localities (Figure 1).

Localities of archeological sites mentioned in the text. Pampa del Tamarugal basin: (1) 01GUA02; Quillagua: (2) QUI93, (3) QUI94, and (4) QUI89; river Loa basin: (5) Chiu Chiu 200, (6) Aldea San Salvador, Río San Salvador; Salar de Atacama basin: (7) Tulán 85.
Pampa del Tamarugal
01GUA027 is a domestic/productive site associated with forests in the Guatacondo locality. This latter was intensively occupied during the Formative period and nourished by stationary floods and underground water (1380 masl) (Meighan, 1980).
River Loa
QUI89, QUI93 (with two differentiated areas, REC-enclosure and MCC-mound), and QUI94 sites are in Quillagua, a fertile oasis in the Loa River (810 masl), combining riverbed vegetation and abundant Prosopis forests to the present day. Aldea San Salvador is a village settlement in the homonymous affluent of the Loa River (1370 masl), tightly associated with the exploitation of a fossilized Prosopis forest, llama herd maintenance, and connectivity with other areas (De Ugarte and Gallardo, 2018). Chiu Chiu 200 is a semi-agglutinated village used seasonally by pastoralist groups who managed llama herds, the nearby Chiu Chiu meadows, and Loa riverbed (2500 masl) (Benavente, 1982).
Salar de Atacama
Tulán 85 (Tu-85) on the southeast border of the Salar de Atacama (2300 masl), is defined as a pastoralist site, near to the Tilocalar meadows, where grasses and shrubs predominate (González-Silvestre et al., 2013).
Raw and calibrated ages relating to each of these sites are given in Supplemental Table S1.
We measured the width and length of each pellet and photographed them before analysis. Following Chame (2003: 74), they have a “cylindrical or rounded shape usually pointed at one end and concave in the other extremity.” Although this shape characterizes Artiodactyles, they have low diagnostic value at the family level and almost none at the generic level (Chame 2003). Using dissecting needles, we carefully opened each pellet and macroscopically analyzed it in search of entire algarrobo seeds or endocarps (Table 1). From this initial sample, we analyzed four subsamples larger than 500 µm from 01GUA027, Qui 93MCC, Qui94, and Tu-85, while adapting Yagueddú’s (2009) protocol for coprolites. The sample was discolored using a 5% solution of sodium hypochlorite until darkish particles became clearer; it was then rinsed with distilled water. One aliquot was mounted in glycerin, and the entire cover slide (24 ×48 mm) was scanned at 20×. The remaining material was saved for future analysis (Velázquez et al., 2019). Phytoliths were classified into different morpho-anatomical categories, which in some cases could be assigned to different taxonomic levels and classified as either monocot or dicot, and, at the most precise level, we were able to compare their identification with individual species (cf.) currently growing in the area. Other phytoliths could not be identified taxonomically due to their redundancy in both classes (e.g. tracheids or hairs); these were therefore characterized only at the morphological-anatomical level. We developed a reference collection using Dacar and Giannoni’s (2001) protocol, which combined reference samples stored at the Archeology Laboratory of Pontificia Universidad Católica de Chile 1 with previous reference databases of microfossils and tissues (Babot, 2009; Giovannetti et al., 2008; McRostie, 2013; Musaubach and Babot, 2019). Microsamples were observed under a LEICA DM750P microscope and photographed with a mounted LEICA MC170 HD camera; macro remains were observed in more detail using a LEICA stereomicroscope S9D and photographed with a LEICA MC170 HD camera.
Coprolite samples analyzed in this study. PUClab and UCH indicate samples stored at the Pontificia Universidad Católica de Chile and the Universidad de Chile laboratories, respectively. We estimated the age range of sites with multiple date chronologies by calculating site phases using the Bchrondensity function in the Bchron package; for single date sites, age range was sourced by estimating credible intervals of the calibrated ages.

Results
Macroscopic observation
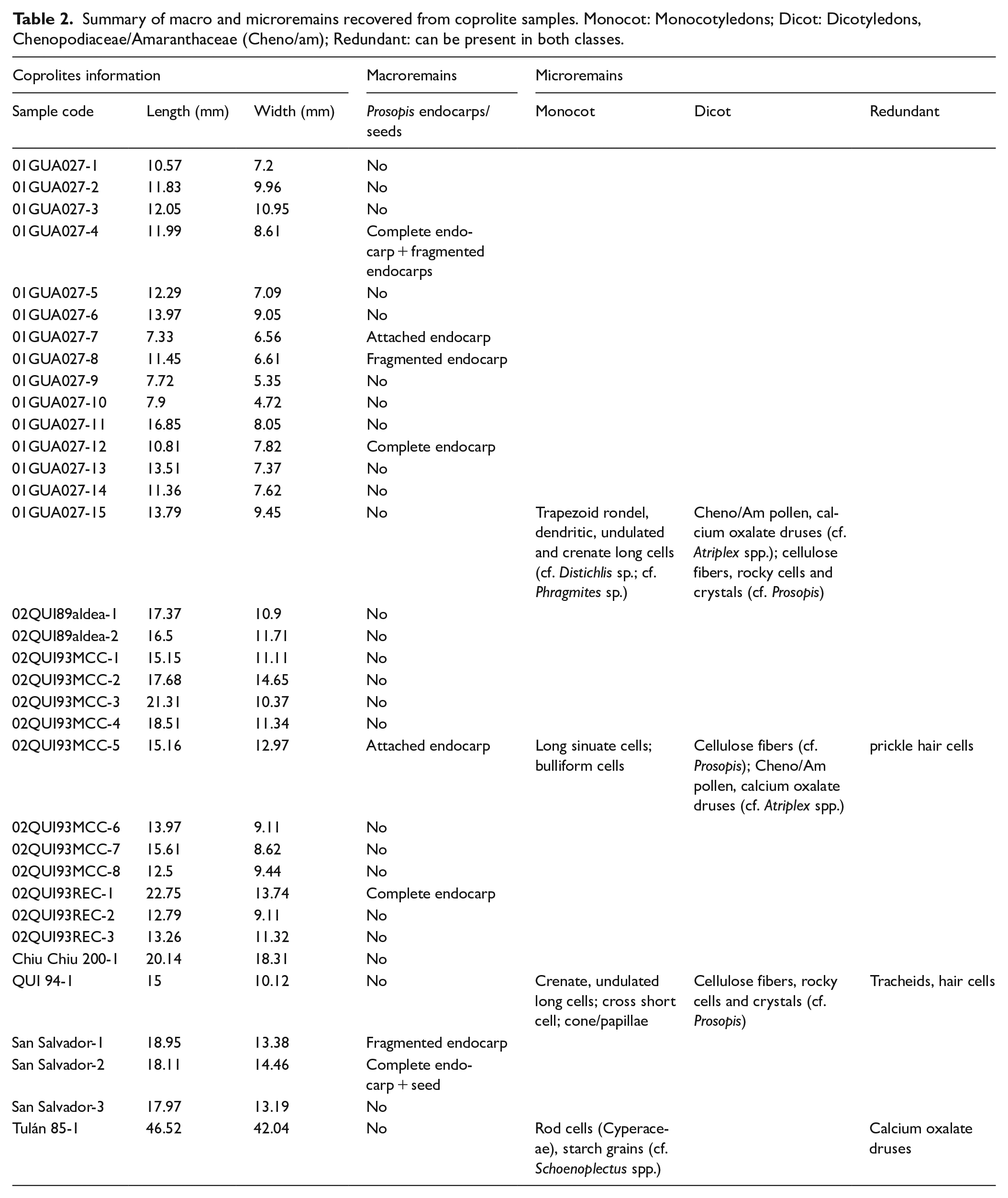
Macroscopic observation shows an intense fragmentation and stripping of chewed and ingested plants. Coprolites are yellowish and heavily fibrous, with an average length and width of 14 and 11 mm, respectively. In our preliminary analysis, we identified tissues such as grasses, reeds, and Prosopis spp. epicarp cellulose fibers (Figure 2a). Tissues and fibers have a maximum length of 18.21 mm and are highly twisted. Macroscopical observation of the reference algarrobo pod collection lead us to conclude that the fragments of yellowish fibers belong to the fibrous external epicarp (Capparelli and Prates, 2015) (Figure 2b). For four of the 34 pellets (11%), entire endocarps and a single algarrobo seed were recovered (samples 01GUA027-4, 01GUA027-12, 02QUI93REC-1, and San Salvador-2); three samples showed fragmented endocarps (samples 01GUA027-4, 01GUA027-8, and San Salvador-1); and two samples showed endocarps attached, probably due to post depositional processes (samples 01GUA027-7 and 02QUI93MCC-5) (Table 2).
Summary of macro and microremains recovered from coprolite samples. Monocot: Monocotyledons; Dicot: Dicotyledons, Chenopodiaceae/Amaranthaceae (Cheno/am); Redundant: can be present in both classes.

(a) Plant macroremains observed in a camelid coprolite from Aldea San Salvador archeological site, (b) epicarp fibers cf. Prosopis, (c) general view of a Prosopis tree, and (d) detailed view of Prosopis tree leaves and seed pods (McRostie, this study).
Microfossil analysis
Analysis of the four coprolites allowed for a preliminary identification of some diagnostic articulated and disarticulated phytoliths, plant tissues, cells, pollen, starch grains, and calcium oxalates (Table 2).
Monocotyledon short and long cell phytoliths were identified as Poaceae and Cyperaceae (Figure 3a and b). Elongated cells found in sedges coincide with those of the grass family (Majumder et al., 2020; McRostie, 2013; Ollendorf, 1992). Compared to the local reference collection, the most probable taxa could be Distichlis spicata, Phragmites australis, and Schoenoplectus spp. A cross was found (Figure 3c). The cross shape has been considered a panicoid marker and hence useful for maize identification (Pearsall, 2000; Twiss et al., 1969), though it is also common in certain Bambusoideae and occurs in small numbers in the Chloridoideae, Arundinoideae, and Pooideae (Piperno and Pearsall, 1998). Dicotyledons were identified by their cellulose fibers, rocky cells, and crystals characteristic of Fabaceae plant fruits, including Prosopis spp. (Figure 3d) (Giovannetti et al., 2008) and pollen grains of Chenopodiaceae/Amaranthaceae associated with calcium oxalate druses, probably Atriplex spp. (Figure 3e and g). Redundant phytoliths were identified as hair cells (Figure 3f). Starch grains were scarce, but in the Tulán 85 sample, we found several grains in situ in parenchyma tissues similar to those retrieved from local reference samples of Schoenoplectus rhizomes (Figure 3h).

Microfossils recovered from camelid coprolites: (a and b) monocotyledon short and long cells, (c) cross-shaped phytolith, (d) cellulose fibers, rocky cells, and crystals, (e) pollen grains of Chenopodiaceae/Amaranthaceae, (f) hair cell phytoliths, (g) calcium oxalate druses, and (h) starch grains in parenchyma tissue packages of cf. Schoenoplectus.
Discussion
Although the sample of camelid coprolites is still small 2 and might be biased by the subsampling of pellets, the taxa identified by macro and microscopical observation correspond to distinct local patches of grazing plants (Villagrán et al., 1998). Some of them, abundant in the archaeobotanical record are still part of the landscape (Distichlis, Phragmites, Atriplex, Schoenoplectus, Prosopis). Only Chiu Chiu 200 and Tulán 85 showed no Prosopis remains. This is consistent with the fact that these sites are located at higher altitudes (2300–2500 masl) and closer to meadows than forests. It is also likely that it reflects a continuity of late Archaic herding practices in the extensive highland grazing patches. Colonization of the lower basins during the Formative period was triggered by a positive but variable wet hydroclimate phase established around 3000 years BP, coupled with the introduction of domestic plants, camelids, and Prosopis forests (Agüero and Uribe, 2011; Berenguer, 2004; Gallardo et al., 2021; Labarca and Gallardo, 2015; Núñez and Dillehay, 1979; Núñez and Nielsen, 2011; Pimentel et al., 2011; Santoro et al., 2017). Social factors such as population growth, tension and conflict, or emerging social stratification, among others (Bollig and Göbel, 1997), would also typify risks emerging during the Formative period in the Atacama Desert (Núñez and Santoro, 2011; Núñez et al., 2006; Uribe et al., 2020). Within this new socioecological scenario, silvopastoralism could have been a relevant risk-spreading strategy in the occupation of these basins. Indeed, at lower elevation sites (800–1300 masl) with fossil or modern Prosopis forests (Guatacondo, Quillagua, San Salvador), macro or micro scale analyses of pellets reveal that camelids were ingesting pods as well as other local grazing plants. This environment is not ideal for camelid herding when compared to the puna’s rich, stable and productive grasslands and bofedales (Kuznar, 2001; Núñez, 2005; Núñez and Santoro, 1988; Rivera, 2005; Santoro et al., 2005; Szpak and Valenzuela, 2020). The breeding of camelids outside of this range might have been restricted to watered salares, oases, and riverine vegetation or irrigated areas, where crops such as maize or seasonal transhumance complemented natural restrictions (Dufour et al., 2014). For the arid coast of northern and southern Peru, several authors have documented the successful breeding of camelids, although in broader and lusher valleys than those in Northern Chile (Cadwallader et al., 2012; Guillet, 1987; Thornton et al., 2011; Wheeler, 1995). Shimada and Shimada (1985: 22–23) propose that camelid maintenance along the Peruvian south coast “may have involved various strategies: grazing in algarrobo forests, lush side valleys, canal and riverbank vegetation, harvested fields and feeding in corrals.” In this sense a high wild-food productivity will be managed for herd animals (Browman, 1974). This encouragement probably also attracted wild fauna, reproducing hunting gathering strategies within the context of these new economies. Most of the ultimately stable isotopic studies carried out to gain insights into lowland husbandry have shown a diverse set of strategies to feed and maintain these herds (Gayo et al., 2020; Mader et al., 2018; Szpak and Valenzuela, 2020; Tomczyk et al., 2019).
Digestion and excretion pathways made by camelids, however, may not have been an efficient seed dispersal mechanism within the Atacama. We found evidence of entire Prosopis seeds/endocarps in 11% of the samples, but suspect that this is not the only variable to affect successful seed germination. Altamirano (2006), referring to P. tamarugo regeneration in Pampa del Tamarugal, describes how fruits fall off the trees when mature, and different herbivores (caprines, ovines, and camelids) can act as disseminators; but seeds dispersed in this way are unlikely to germinate due to aridity of the soil and the environment in general. The author says that natural regeneration is occasional, scarce and depends on floods from the Andes that scarify these seeds with mud and stones, decanting them in watered areas where they are prone to germination. This process has also been detected in Quillagua, within the Loa riverbed, where small Algarobia tree species grow following the path of water. In Argentina, endozoochoric studies of Prosopis relating to different herbivores, among them guanaco, show that this latter specie allows for seed germination in 41% of P. chilensis and 24% of P. flexuosa in San Juan province. It may be possible that an opportunistic frugivore such as the guanaco is followed by rodents or other animals that recover seeds and fruits from their dung (Campos and Velez, 2015). Survival of seeds therefore depends on several variables, emphasizing aridity, plant species, animal dispersers, as well as bruchid attacks (Campos and Velez, 2015; Campos et al., 2008).
Bruchids are not a minor variable in the equation (Velez et al., 2018; Pasiecznik et al., 2001). Although no precise studies have been made to assess the extent to which these insects attacked pre-Columbian algarrobos, their activity is evident in many archeological endocarps and pods. “Bruchids oviposit exclusively on seeds, or seed envelopes, suitable for larval development” (Kingsolver et al., 1977). They start to form in immature pods until after the latter have matured and fallen to the ground (Johnson, 1983). These insects greatly reduce the numbers of viable seeds.
The ethnographic holding of camelids in Prosopis spp. forests are difficult to find nowadays and most traditional cultural-culinary practices associated with Prosopis have been replaced by European foodways and resources (Carmona, 2018). In some modern oasis and riverine systems deprived of herding practices, there is a heavy accumulation of pods below the parent trees. The high concentration of sugary pods is a good spot for bruchids to lay their eggs and reproduce, minimizing the spread and regeneration of these forests. Delatorre (2005) interprets ancient P. alba populations in the Pampa del Tamarugal basin as relicts, considering that most of these northern Algarobia populations are confined to isolated patches (Bessega et al., 2021; Carevic et al., 2012; Delatorre, 2005). This scenario resembles what Janzen and Martin (1982: 21) described as neotropical anachronisms, regarding the extinction of Gomphotheres and the Scheelea palm, “within a month as many as 5000 fruits accumulate . . . as the pulp rots off fallen fruits beneath the parent palm, the bruchids oviposit on virtually all of the exposed nuts.” The bulk of the seeds perish directly below the parent, inhibiting the spread of still viable seeds by overshadowing.
As preliminary analyses become more solid, we will be able to establish the role of camelids as vectors of these trees, while understanding human agency and anthropogenization of these oases, especially if desert distances between them are too large great for herbivores to act as dispersers, or for water to connect trees populations by floods (Bessega et al., 2021; McRostie, 2014; McRostie et al., 2017). Indeed, when Gajardo (1994) defines the vegetation of these oases,’ he states that the current level of human intervention makes it hard to define what was the “original vegetation composition.” It is likely that the cultural modification of these landscapes began several millennia ago during the Formative period, with agroforestry and silvopastoralism being a primary activity in the reproduction and inhabitation of these landscapes. “Even the simplest agroforestry system is more complex, ecologically (structurally and functionally) and economically, than a monocropping system” (Nair, 1993: 14). Local knowledge of silvopastoralism requires further attention. Studies in Santiago del Estero, Argentina, have shown that a silvopastoral strategy of pasturing camelids in areas dominated by P. torquata will encourage in them strong fiber growth (Díaz and Roger, 2015).
As global climate change impacts territories and traditional risk-spreading strategies are hard to accomplish for several reasons related to modern lifestyles impacting ancient practices – including changes in land use, water scarcity, reduction in the scale of pastoralism, monocultivation, loss of pedestrian long-distance trading and nets – we look forward to understanding how our landscapes came into being. By improving our understanding of the past, we might be able to co-produce knowledge with local communities in major initiatives such as ecosystem restoration and conservation. The present-day scenario of the Atacama Desert seems riskier than that of the Formative period. By looking at these old ancient socioecological strategies, we might be able to achieve greater levels of resilience.
Supplemental Material
sj-xlsx-1-hol-10.1177_09596836221122636 – Supplemental material for Silvopastoralism and the shaping of forest patches in the Atacama Desert during the Formative Period (ca. 3000–1500 years BP)
Supplemental material, sj-xlsx-1-hol-10.1177_09596836221122636 for Silvopastoralism and the shaping of forest patches in the Atacama Desert during the Formative Period (ca. 3000–1500 years BP) by Virginia McRostie, Pilar Babot, Elisa Calás, Eugenia Gayó, Francisco Gallardo, Carolina Godoy-Aguirre, Rafael Labarca, Claudio Latorre, Lautaro Núñez, Karla Ojeda, Calogero M Santoro and Daniela Valenzuela in The Holocene
Footnotes
Acknowledgements
We are grateful to Alison Damick, Camille Weinberg, Emily Dawson, Arlene Rosen, Graeme Barker, Matías Frugone, Mauricio Uribe, Rocío Mardones, Camila Opazo, Antonia Benavente, Horacio Larraín, and reviewers of this manuscript. We also thank the Universidad de Chile Archeological Laboratory and the Pontificia Universidad Católica de Chile Archeological Laboratory.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding for this work was provided by Fondecyt projects #11180805, #1160045, #3150638, #1070040, #1201687, SOC 114005, Grant ANID PIA/BASAL FB210006, UTA MAYOR 3738-19, ANID/FONDAP/15110009, and the ANID-Millenium Science Initiative Program - NCN19_153.
Supplemental material
Supplemental material for this article is available online.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.