
Abstract
The underrepresentation of many important rainforest species in pollen records still hinders accurate paleovegetation reconstructions in tropical Southeast Asia. In this study, we conducted a modern phytolith study of 141 plant and 15 surface soil samples around the forests of Jerai Hill in western Peninsular Malaysia to evaluate the potential of phytolith assemblages as a proxy for indicating tropical vegetation types. In the plant samples, including many Dipterocarpaceae species,
Introduction
The Quaternary vegetation history of Southeast Asia remains enigmatic although it bears directly on the biogeographical evolution (Woodruff, 2010), terrestrial carbon cycle (Page et al., 2011), and hominin dispersion processes (Roberts and Amano, 2019) within the region. For instance, many efforts based on marine sediments (Chivas et al., 2001; Kaars et al., 2000; Moss and Kershaw, 2007; Sun et al., 2000, 2002; Wang et al., 2009), lacustrine deposits (Hunt et al., 2012; Jones et al., 2014; Penny, 2001; Taylor et al., 2001; White et al., 2004), and biogeographical records (Meijaard, 2003) have been made to explore the vegetation pattern of Southeast Asia during the glacial periods, whether there were widespread grasslands on the emergent Sundaland is still hotly debated (Heaney, 1991).
In paleovegetation reconstructions, fossil pollen is considered valid; however, many arboreal species in the rainforests of Southeast Asia have very low pollen representation (Cheng et al., 2020). In particular, plants of the Dipterocarpaceae family, which are the most popular components of lowland rainforests, are rarely recorded in palynological records (Barboni and Bonnefille, 2001; Cheng et al., 2020; Sun et al., 2002; Wang et al., 2009). Phytoliths are made from noncrystalline silicon dioxide, distinct from pollen exine, which is mainly composed of organic sporopollenin; the former is less vulnerable to oxidizing conditions than the latter in paleosols, which induces different implication biases for paleoenvironmental reconstruction (Strömberg et al., 2018). For instance, Bambusoideae, Oryzeae, and Panicoideae produce many diagnostic phytoliths, such as tall narrow
In Southeast Asia, Kealhofer (2003) reported a detailed classification of Bambusoideae, Oryza, Panicoideae, Araceae, Musaceae, and some other arboreal phytoliths, confirming the potential of phytolith analysis for reconstructing past tropical landscapes during the Late Pleistocene. Additionally, Kealhofer and Piperno (1998) presented many phytolith morphotypes of tropical plants, in which some diagnostic forms were significant to indicate a variety of plants, such as
In tropical regions, the relationship between phytoliths and plants could be complex for three reasons: (1) a given plant can produce several phytolith morphotypes, and the same morphotype occurs in different species (Collura and Neumann, 2017; Kealhofer and Piperno, 1998; (2) variations in phytolith production influence its representation in sediments (Mercader et al., 2009); (3) arboreal phytoliths have a low resolution for distinguishing different families, of which nondiagnostic phytoliths are common in the tropical plants of Southeast Asia (Kealhofer and Piperno, 1998). To date, a few diagnostic phytoliths have been extracted from tropical trees, in which spheroids with varying characteristics are the most typical (Alexandre et al., 1997; Kealhofer and Piperno, 1998; Neumann et al., 2009).
In this study, we analyzed phytoliths collected from plants and surface soil along tropical Jerai Hill in western Peninsular Malaysia (Figure 1). The objectives of this research were to (1) clarify the phytolith morphotypes of some tropical plants that have never been investigated, and the characteristics of the phytolith assemblages in tropical forests and (2) evaluate the potential use of the phytolith assemblages in the reconstruction of past tropical forests in Southeast Asia.
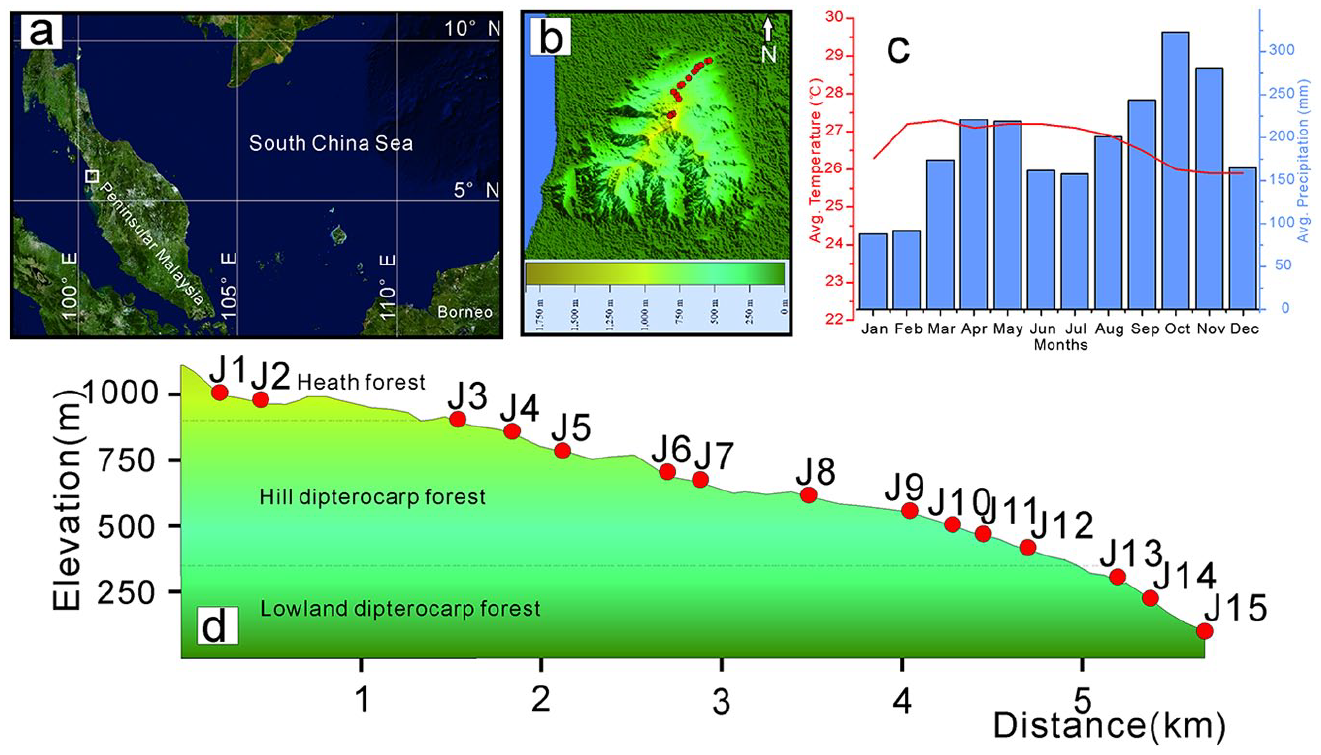
Location of sample sites: (a) the white box marks the location of Jerai Hill, (b) the topography of Jerai Hill and distribution of sampling sites (red dots), (c) monthly mean precipitation (blue bars) and air temperatures (red curves) of the Jerai Hill region, and (d) vertical profile of Jerai Hill, with sampling sites labeled.
Materials and methods
Study area
Jerai Hill is located in Kedah State of Peninsular Malaysia, where the major vegetation communities are tropical rainforests and montane rainforests, which are typical in Southeast Asia (Saw, 2010). The climate is characterized by a moderate seasonal contrast in precipitation, with a relatively dry season from January to February (Figure 1c). The summit (05°47′N, 101°26′E) of the hill has an elevation of 1200 m (Scientific team under the auspices of the WWFM, 1977) (Figure 1d). There are three dominant vegetation types (Whitmore, 1998). At low elevations (<300 m), the typical vegetation type is lowland rainforest rich in Dipterocarpaceae species, including Anisoptera spp., Dipterocarpus spp., Dryobalanops spp., Hopea spp., Parashorea spp., and Shorea spp. Dipterocarps comprise 50% of the individuals of the upper canopy (Saw, 2010; Whitmore, 1998). Other common large trees include Dyera costulata (Apocynaceae), Gluta spp. (Anacardiaceae), Heritiera spp. (Malvaceae), Intsia palembanica (Leguminosae), Koompassia malaccensis (Sapotaceae), Palaquium spp. (Sapotaceae), and Sindora spp. (Leguminosae). The understory tree layer mainly comprises saplings of the upper canopy, shrubs, and climbers from some families such as Annonaceae, Euphorbiaceae, Flacourtiaceae, Melastomataceae, and Rubiaceae (Saw, 2010). With an increase in elevation, the richness of dipterocarp species decreases sharply between 300 and 700 m a.s.l., and Shorea spp. begin to be the most dominant species (Whitmore, 1998). The upland taxa frequently occurring above 900 m a.s.l. are predominated by Leptospermum spp. (Myrtaceae), Fagaceae species, and Dacrydium elatum (Podocarpaceae) (Scientific team under the auspices of the WWFM, 1977). Poaceae plants are patchily distributed along forest margins or at open sites at middle and high elevations. Other nonarboreal species are rare and are primarily represented by Marantaceae and Zingiberaceae.
Sampling
We analyzed 141 plant samples from 53 families and 132 species, including many important taxa of tropical forests, such as Hopea spp. and Shorea spp., among which 44 samples were collected around Jerai Hill, and 97 samples were obtained from the collections of Universiti Sains Malaysia (herbariums) and Penang Botanical Garden (fresh) (Table 1). Fourteen surface soil samples were collected beneath the forest litter at altitudes ranging from 200 to 1000 m a.s.l., where the vegetation types change from lowland rainforest to heath forest (Figure 1). No samples were taken below 200 m a.s.l., because of obvious anthropogenic disturbances along the northern slope of Jerai Hill. Instead, we used a surface sample from an adjacent site (75 m a.s.l.; 5°26′19.31″N, 100°17′15.42″E), which has a more natural floristic composition than that of Jerai Hill.
List of plant samples and phytolith morphotypes.
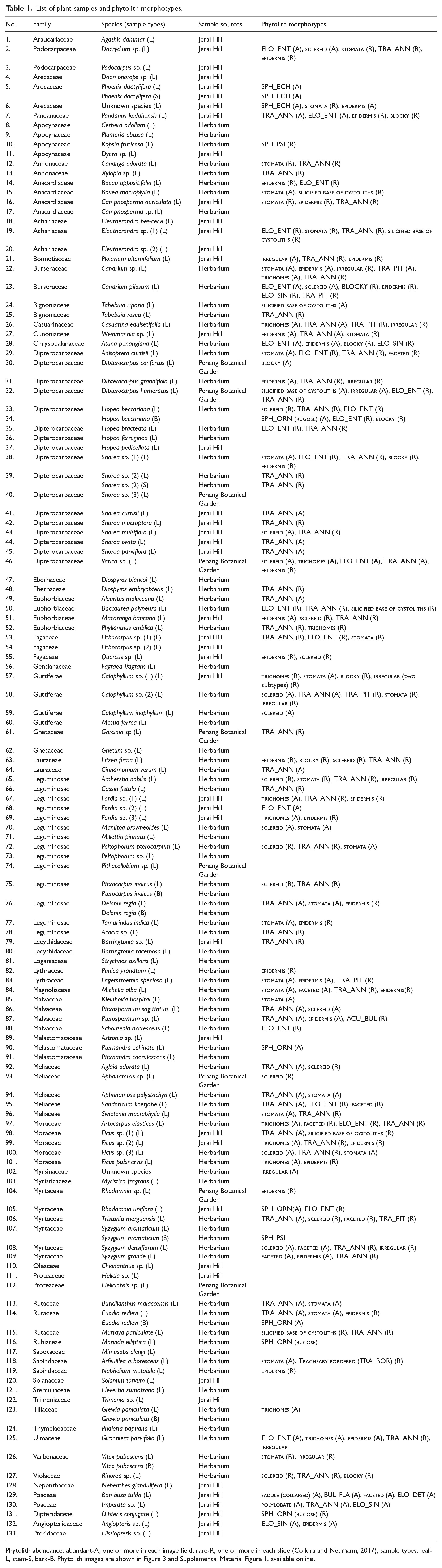
Phytolith abundance: abundant-A, one or more in each image field; rare-R, one or more in each slide (Collura and Neumann, 2017); sample types: leaf-L, stem-S, bark-B. Phytolith images are shown in Figure 3 and Supplemental Material Figure 1, available online.
Laboratory analysis
Approximately 10 g of soil from each surface sample was analyzed. Samples were heated in hydrochloric acid (HCl) (15%) and hydrogen peroxide (30%) at 60–80℃ for 2–3 h to remove carbonates and organic components. Phytoliths were extracted from the residues using a heavy liquid extraction technique. The heavy liquid was composed of Zn grains, potassium iodide, and hydroiodic acid, and the specific gravity was set to 2.385. To increase the detection probability of rare and crucial taxa, such as Poaceae short cells (Weng et al., 2006), the phytolith count was raised to at least 500 phytoliths.
For morphological observation, the leaf samples were heated in nitric acid (HNO3) at 100℃ for 4 h to remove the organic components, and the remaining phytoliths were concentrated after the organic components were dissolved. Before the acid treatment, all the specimens were repeatedly cleaned using a toothbrush in an ultrasonic water bath. The phytolith samples were mounted on slides in Canada Balsam and counted at 400× or 1000× magnification. For each sample, at least five slides were observed.
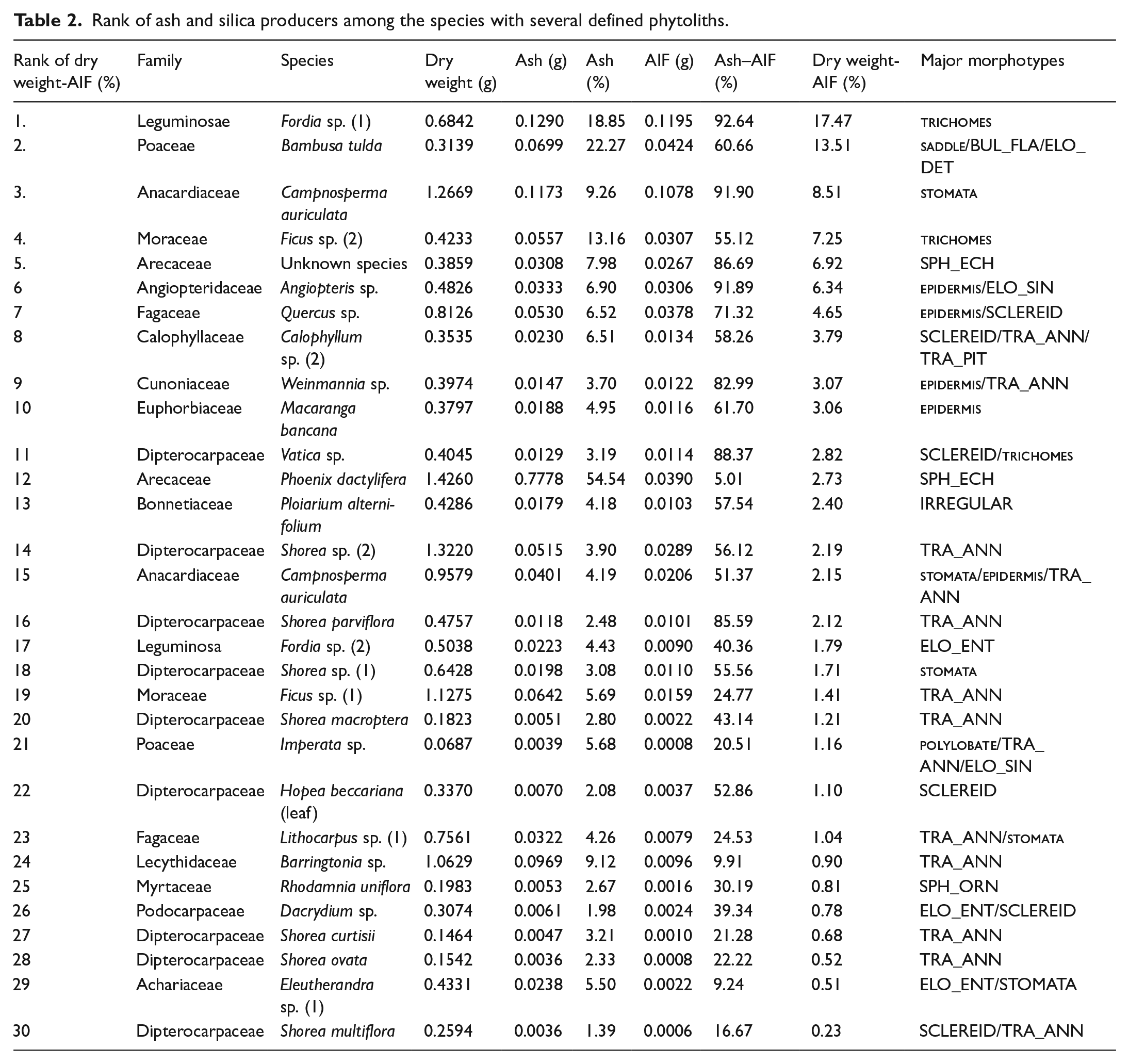
To detect phytolith productivity, we resampled 30 species that contained an identified phytolith (0.07–1.43 g dry weight) and used a modified dry and wet ashing method, according to Collura and Neumann (2017), Mercader et al. (2009). The processes were as follows: (1) the leaf sample was dried and weighed; (2) the samples were placed in a muffle furnace at 500 ℃ for 8 h, and the mass was measured on a high-precision balance after cooling; (3) the mass of the resulting ash was transferred to a dried and weighed centrifugal tube, and then ash was treated with 10 ml HCl at 95℃ to remove the carbonate components; (4) the acid was removed from the centrifugal tube through successive washing cycles by centrifugation for 5 min at 3000 rpm; (5) to remove the organic matter, the remainder of the sample was treated with 10 ml HNO3 at 95℃ for 4 h, washed three times with 95% ethanol, centrifuged and dried; (6) we weighed the centrifugal tube and calculated the difference between it and an empty tube to obtain the weight of the phytoliths. The resulting biominerals formed the acid-insoluble fraction (AIF), which could be considered to be phytoliths (Mercader et al., 2009). The naming and description of the phytoliths were based on the International Code for Phytolith Nomenclature (ICPN) 2.0 (International Committee For Phytolith Taxonomy (ICPT), 2019) and on studies of modern phytoliths produced by woody taxa (Kealhofer and Piperno, 1998; Mercader et al., 2009).
Result
Phytoliths in plants
Among the 141 studied plant samples, 73% yield phytoliths, in which
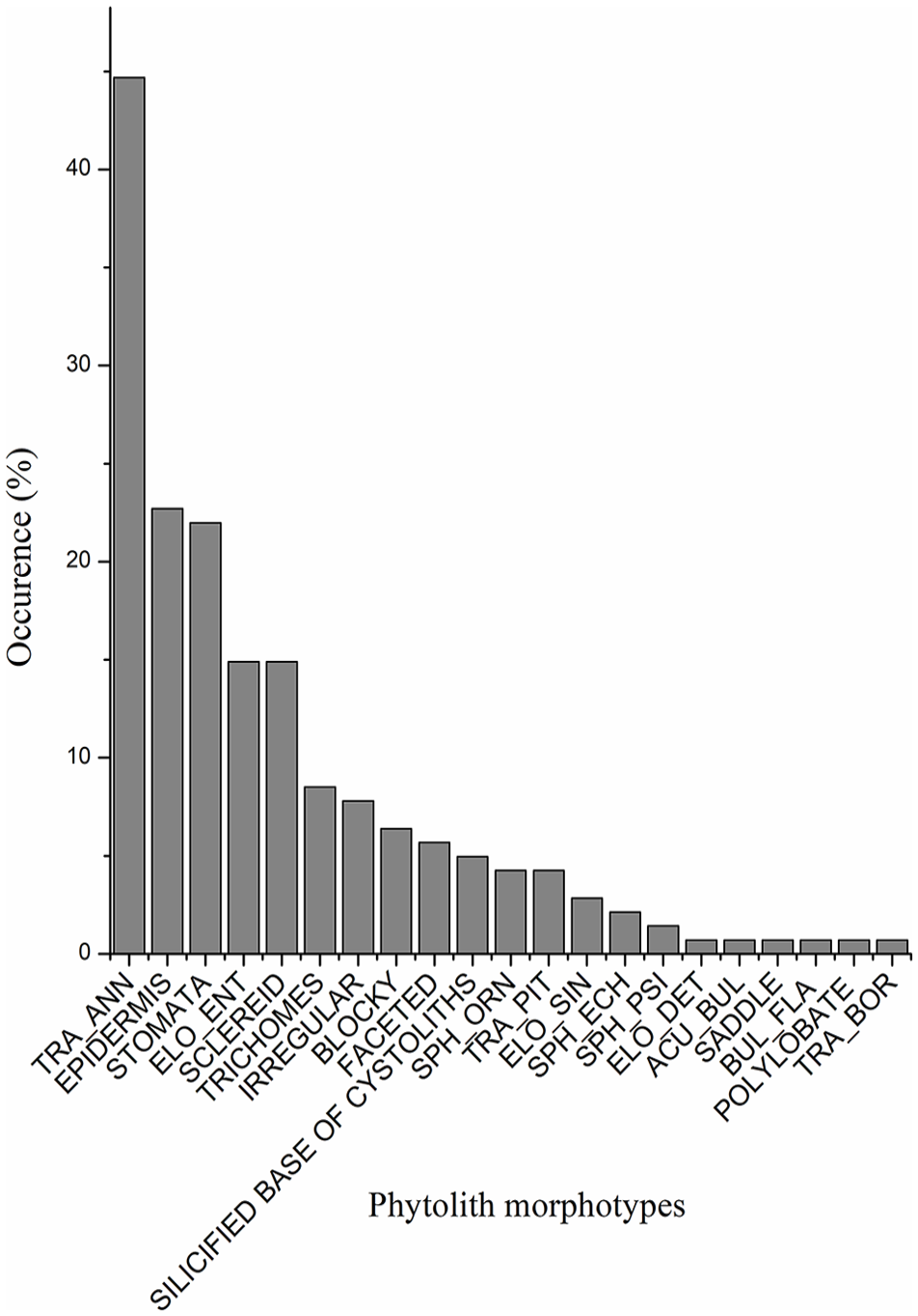
Distribution of morphotype classes in 141 plant samples.

Phytolith morphotypes from the plants. a–i, TRA_ANN; j–l, ELO_ENT; m,
Several plants contain
Phytoliths in surface soil samples
The phytolith assemblages are dominated by spheroidal types (on average 75%) (Figure 4), among which

Phytolith assemblages in the surface soil on Jerai Hill. The F/P index is the ratio of the forest indicator phytoliths to the short-cell phytoliths of Poaceae. In samples #13 and #14, the F/P values were missing because of the absence of Poaceae phytoliths. Partial data are referred from Bai et al. (2020). The column on the right lists the dominant pollen taxa and their percentages in the topsoil pollen assemblage on Jerai Hill (Cheng et al., 2020).

Morphological images of phytoliths from the surface soil.
Discussion
Comparison between phytoliths from plants and surface soils
Phytoliths with a high occurrence in the plant samples do not correspond to prominent phytoliths in the surface sediments (Figures 2 and 4). The former mainly includes TRA_ANN,
Rank of ash and silica producers among the species with several defined phytoliths.
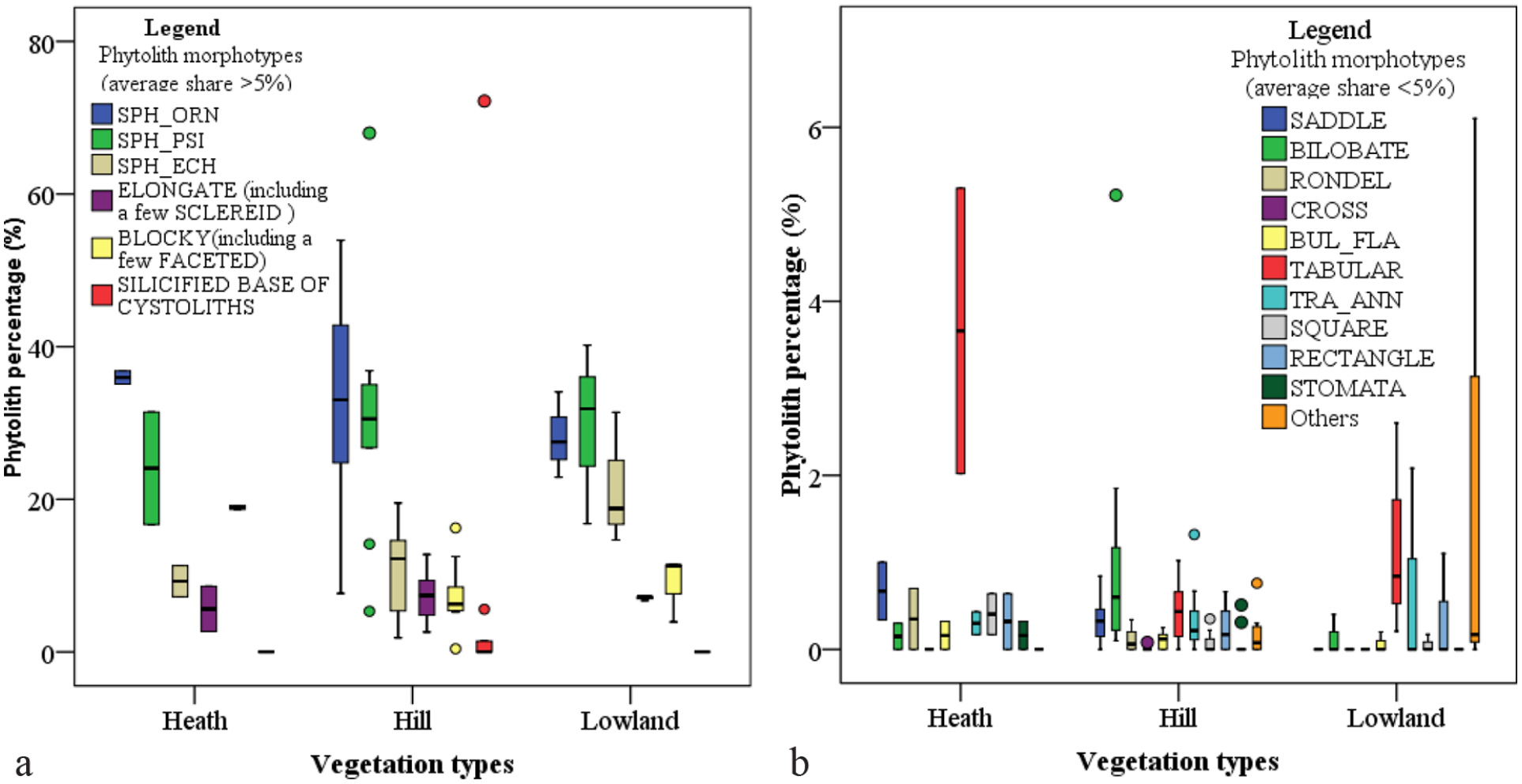
Box plot providing the phytolith percentages and standard deviations in the three vegetation types. The dots mark the samples at different altitudes with a large percentage deviation.
The implication of phytolith assemblages for the rainforest structure and openness
On Jerai Hill, dipterocarp forests rich in Dipterocarpaceae species are the most popular vegetation types. In this study, phytolith investigation of 18 Dipterocarpaceae samples showed that they mostly produced nondiagnostic phytoliths, such as TRA_ANN and
The abundance of Poaceae phytoliths in the heath forest and hill dipterocarp forest was higher than that in the lowland dipterocarp forest, where the forest canopy is denser and the understory is sparser (Figure 4). According to our field investigation and publications (Scientific team under the auspices of the WWFM, 1977), there was sparse nonarboreal distribution in the Jerai forest, particularly Poaceae plants primarily restricted within the canopy gap. The high silicic productivity of Poaceae species, such as Bambusa tulda (Table 2), favors the maintenance of its representation in phytolith assemblages (Figure 4). Therefore, the abundance of Poaceae short-cell phytoliths is closely related to forest openness. In addition, the D/P (or F/P) index based on Poaceae phytoliths may be used to distinguish between grassland and forest in Southeast Asia.
The D/P index was first proposed by Alexandre et al. (1997), who considered that the proportion of the dicotyledonous woody phytoliths (SPH_ORN) versus Poaceae phytoliths is an indicator of forest openness in intertropical Africa. A similar D/P index was also used in Ethiopia (Barboni et al., 1999) and was further calculated using the quantitative relationship between the D/P index and the vegetation cover in southeastern Cameroon (Bremond et al., 2005, 2008). Many studies have determined that SPH_ORN phytoliths are dominant in tropical forests (Alexandre et al., 2012), and their share in the forests of the southwest Amazon can reach 30–60% (Watling et al., 2015), which is much more than those in the steppe or savanna. In the mountain forests of tropical Africa, the value of the D/P index ranged from 1 to 6 (Bremond et al., 2005), and a shift in this index from 5.54 to 0.21 was considered to be an indication of the change from forests to open habitats (Astudillo, 2018).
In a variety of studies, the D/P index might be given a different definition. In the tropics of the eastern equatorial Pacific, the ratio of forest indicator phytoliths to the sum of woody and grass short-cell phytoliths (FI-t ratio) was estimated (at 93.61–99.76), where all phytolith morphotypes produced by monocotyledonous and dicotyledonous trees were considered (Crifò and Strömberg, 2020). On Jerai Hill, the ratio of the forest indicator phytoliths to the Poaceae short-cell phytoliths, that is, SPH_ORN phytoliths versus the sum of
Compared with pollen, the phytolith assemblage provides some specific details regarding vegetation components: (1) The SPH_ORN-dominated phytolith assemblage highlights the wide distributions of palm, some trees, and understory herbs, which are poorly reflected in gymnosperm- and fern-dominated pollen assemblages (Figure 4) (Cheng et al., 2020), (2) the share of Poaceae pollen irregularly changes with altitude due to its outstanding dispersal ability (Cheng et al., 2020). By comparison, the spatial distribution of Poaceae phytoliths over horizontal space is more sensitive to their parent plants (Figure 4).
The implication of phytolith assemblage for distinguishing vegetation types
There are three dominant vegetation types on Jerai Hill, which, however, seem to be ambiguously distinguished by phytolith assemblages (Figure 4). The principal component analysis (PCA) results for the phytolith morphotypes indicate that only 25.1% of the total variance in the dataset could be accounted for by ordination in the first principal component (Axis1) (Figure 7a). The PCA result demonstrates a poor relationship between the phytolith assemblages and environmental factors, and the reason could be further revealed by the nonmetric multidimensional scaling (NMDS) analysis (Hammer et al., 2001; Shi, 1993). The NMDS result showed an ambiguous distance gradient of the phytolith data between the three vegetation belts, especially within the hill dipterocarp forest (Figure 7b). The stress value of 0.1159 (full score = 1) was not sufficient to indicate a close relationship between the phytolith data and the spatial distribution of the sample sites (Shi, 1993). However, we found that if several scattered sample points in the hill dipterocarp forest are excluded, the distributions of the sample points along coordinate 1 correspond well to their distributions along the altitudes (Figure 7b). In fact, the hill dipterocarp forest contains many generalist species along the hill slope, and it is a transitional zone from lowland dipterocarp forest to heath forest (Figure 1d). Additionally, several samples, such as those at altitudes of 595 and 700 m, contain an abundant

Biplot of the PCA (a) and NMDS (b) analysis results. For both analyses, the variate included all the phytolith type percentages and 15 samples presented in Figure 4. The NMDS analysis was based on the similarity measure of the 2-dimensional Euclidean solution, and the labeled numbers are the elevations (m).
Conclusions
This study provides a new clue to explore the distribution of phytolith morphotypes in tropical plants, their relationship with phytolith assemblages in sediments, and the potential implications of phytoliths for tropical forest structures in Southeast Asia. The following conclusions can be made:
In 141 plant samples, most of the phytoliths were nondiagnostic, such as TRA_ANN,
In Jerai Hill, the phytolith assemblage in the topsoil is dominated by the SPH_ORN, SPH_ECH, and SPH_PSI phytoliths. Their rare occurrence in the plant samples likely indicated that these phytoliths are mainly not from the dipterocarps that form the vegetation community but from the nonconstructive species.
The F/P index and many unrecorded diagnostic phytoliths in the plant and sediment samples show the potential of phytoliths to distinguish between forests and open habitats in Southeast Asia; phytoliths originated from Palmae, Poaceae, and some forest indicators are more sensitive to indicate their parent plants than pollen.
The statistical analysis revealed the differential phytolith assemblages in the lowland dipterocarp forest and the heath forest, indicating the potential significance of phytoliths for making discrimination between various vegetation types in Southeast Asia.
Supplemental Material
sj-docx-1-hol-10.1177_09596836231157064 – Supplemental material for Phytolith analyses of tropical plants and topsoil from western Peninsular Malaysia and their implications for paleoenvironmental reconstruction
Supplemental material, sj-docx-1-hol-10.1177_09596836231157064 for Phytolith analyses of tropical plants and topsoil from western Peninsular Malaysia and their implications for paleoenvironmental reconstruction by Lu Dai, Yu Bai, Swee Yeok Foong and Zhongjing Cheng in The Holocene
Research Data
sj-xlsx-2-hol-10.1177_09596836231157064 – Research Data for Phytolith analyses of tropical plants and topsoil from western Peninsular Malaysia and their implications for paleoenvironmental reconstruction
Research Data, sj-xlsx-2-hol-10.1177_09596836231157064 for Phytolith analyses of tropical plants and topsoil from western Peninsular Malaysia and their implications for paleoenvironmental reconstruction by Lu Dai, Yu Bai, Swee Yeok Foong and Zhongjing Cheng in The Holocene
Footnotes
Acknowledgements
We are deeply grateful to Prof. Xiangjun Sun in Tongji University, Prof. Houyuan Lu in Institute of Geology and Geophysics, Chinese Academy of Sciences, Prof. Yansheng Gu in China University of Geosciences, Prof. Dongmei Jie in Northeast Normal University, and Prof. Xinxin Zuo in Fujian Normal University for their constructive discussions throughout this study. Special thanks to the two reviewers for their critical comments to improve our manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Natural Science Foundation of China (NSFC, Grant 41776063 and 42002026) and State Key Laboratory of Marine Geology, Tongji University, Open Project Fund (Grants, MGK1822 and MGK202101).
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.