
Abstract
Palaeoecological and archaeological studies conducted in Mozgawa (southern Poland) demonstrated a long-term sustainable land-use by inhabitants of a large settlement occupied during the second half of the fourth millennium BC. The Middle Neolithic society established a settlement that covered an area of about 30–35 ha and functioned during at least three centuries, as validated by the absolute chronology. A thorough analysis based on the fuzzy-logic principle combined with archaeobotanical and archaeozoological data showed that the subsistence model was based on agricultural practices, related with both crop cultivation and animal husbandry, together with an important component of hunting, fishing and gathering. Ecological requirements of plants and animals represented by fossil remains suggested that the exploited area included a variety of habitats. In the open landscape, wet and dry grasslands, used partly as pastures, cultivated fields and marshes were present. Wooded areas included deciduous lime-oak-elm forests, also of open canopy type, and riparian forests. This was due to the transitional location of the Mozgawa site on a loess-mantled hill, covered with fertile soils, but right on the border with a vast alluvial plain of the Nida River. Such ecologically diversified micro-region became a managed landscape, which provided rich resources for seasonal diets of the settlement’s inhabitants. The case study of the Mozgawa site offers a unique insight into the palaeoeconomy of the Funnel Beaker culture in central Europe.
Keywords
Introduction
During the first half of the fourth millennium BC a new Neolithic phenomenon developed in central and partly in northern Europe, archaeologically known as the Funnel Beaker Culture (FBC, also known as Trichterbecherkultur in German) (e.g. Midgley, 1992) or the Funnel Beaker Complex (Furcholt et al., 2014). It is considered that it was formed as a consequence of complex interactions between post-Linear farming groups and still existing hunter-gatherers (e.g. Kabaciński et al., 2015; Klassen, 2004; Nowak, 2009, 2019; Sørensen, 2014a, 2014b; Terberger et al., 2018). By the end of the first half of the fourth millennium, this cultural model finally replaced the last post-Linear communities as well as comprised also territories settled so far by hunter-gatherers. This culture appeared in southern Poland, including its eastern part, historically called Lesser Poland. In this territory, the south-eastern group of the Funnel Beaker Culture (SE FBC) has been discerned (e.g. Burchard et al., 1991; Midgley, 1992; Nowak, 2017; Wiślański, 1979). The number of SE FBC sites is very high, although the majority of them are only known from surface surveys (e.g. Król, 2019; Nowak, 2001; Pelisiak, 2018). They differed in their size and possibly functions, ranging from small (1 to 2 ha), and middle (2–5 ha) to large settlements (up to 30–40 ha). The latter group is represented in the SE FBC only by a few sites (Nowak et al., 2022).
One of the recently excavated large settlements of the SE FBC is Mozgawa covering an area of ca. 30–35 ha (Korczyńska et al., 2019; Nowak et al., 2022). In the light of radiocarbon dating, this settlement developed between ca. 3550-3200 BC (Korczyńska et al., 2019; Moskal-del Hoyo et al., 2018). The present work is the first attempt to discuss jointly plant and animal remains excavated at the site in order to understand the rules of land-use practiced by communities inhabiting the settlement. Having analysed bioarchaeological assemblages, both qualitatively and quantitatively, a few research questions have arisen: (1) Is it possible to specify potential habitats from where plant and animal resources were taken? (2) Have there been spatial changes in plant and animal use during the development of the settlement? (3) Does a long-term, sustainable land-use model/s, which ensured food supply to numerous inhabitants existed? Therefore, the main aim of the presented research is to track the subsistence strategy and land-use during the long-term occupation of the settlement with the aim of answering these questions. New results from Mozgawa may broaden the knowledge about the complex use of local resources and landscapes in the south-eastern group of the FBC.
Research area
Location of the site
The archaeological site in Mozgawa is located in the upper part of a hill elevated above the Nida River valley and near the Mozgawka stream (Figure 1), in the Lesser Poland Upland. The site is situated in the borderland between the Wodzisław Hummock and the Nida Valley (Kondracki, 2002). The entire region is characterised by the upland character of the relief, the compliance of the Nida Basin depression with the tectonic structure, the preservation of the pre-Quaternary old relief, and the lack of a continuous and thick cover of glacial deposits significantly altering the older relief (Radłowska, 1966; Gilewska 1972; Cabaj and Nowak, 1986; Rutkowski, 1986; Urban 2014). The Nida River is created from the connection of two rivers, Czarna Nida and Biała Nida, and the area near Mozgawa belongs to the middle Nida valley, which is filled with Quaternary sediments, including glacial deposits, and the right-bank part of the catchment, Wodzisław Hummock, is covered with loess (Łyczewska 1969, 1971, 1972; Gałka 2006). On the plateaus there is the primary loess accumulated by aeolian processes. On the slopes and in the valleys, apart from the aeolian loess, there are secondary ‘loess’ covers formed as a result of the redeposition of primary loess by exogenous processes at various periods (Jersak et al., 1992; Śnieszko, 1995).
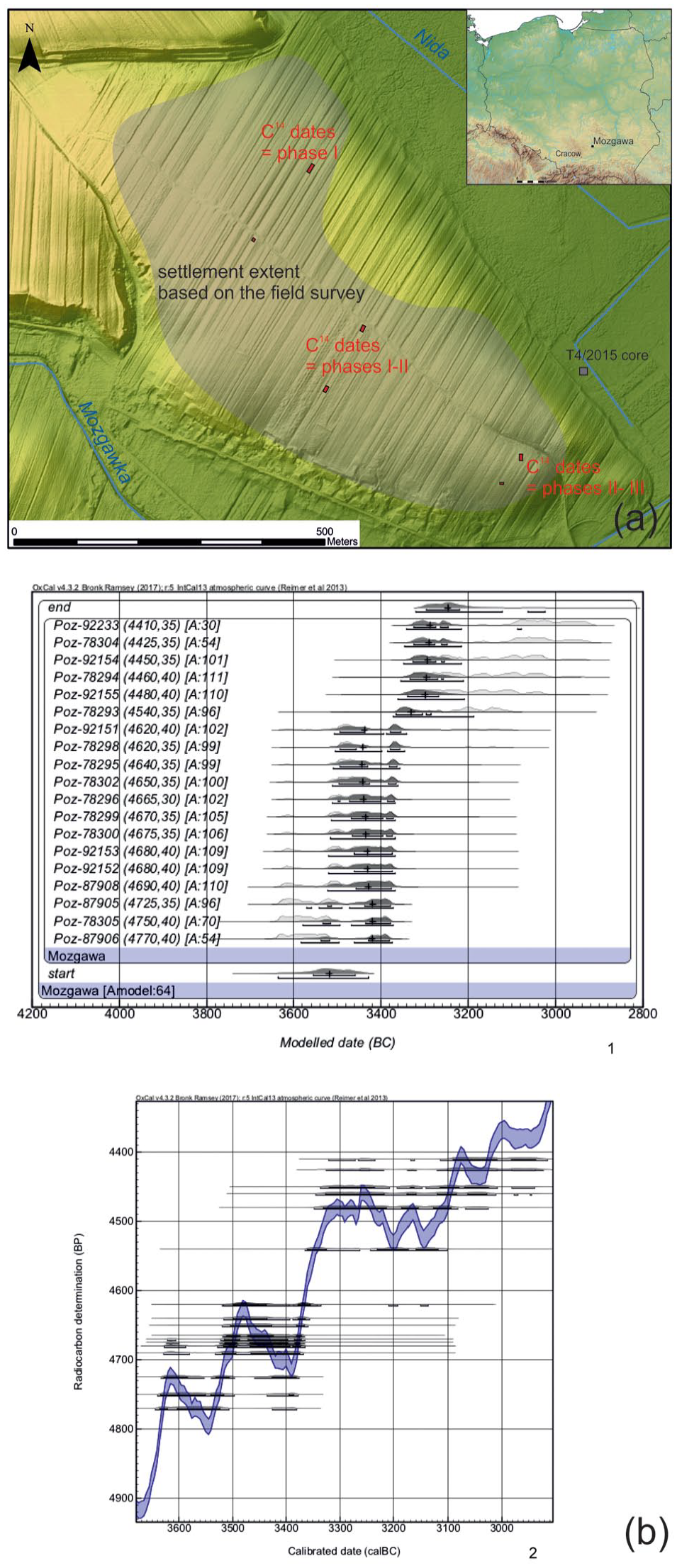
Location of the Funnel Beaker Culture settlement in Mozgawa (a) and calibration of the radiocarbon dates of the settlement against the background of the calibration curve and their Bayesian modelling ((b) – after: Korczyńska et al., 2019). Please note that agreement indices are below 60% in case of three dates, however it does not necessarily indicate that there is a problem with the model as a whole (Bronk Ramsey, 2009: 1025).
Periodisation of the Funnel Beaker Culture settlement in Mozgawa
On the hill where the settlement of the FBC was identified no remains of any earlier settlements or camps were discovered during the archaeological works. Therefore, it seems that it was founded in a pristine and natural environment. According to the Bayesian modelling of the 19 radiocarbon dates (Korczyńska et al., 2019), the occupation of the site by a FBC community started ca. 3550/3500 BC and vanished not earlier than ca. 3250/3200 BCE. Pottery stylistics and reference of modelled dates to the calibration curve have made it possible to discern three basic phases of the settlement (M-I, M-II and M-III). However, the distinction between M-II and M-III is rather vague, both in terms of pottery and radiocarbon dates. Therefore, a division into only two basic developmental stages, namely M-I and MII/MIII, is currently the more reasonable and certain one. The chronological boundary between them would most likely fall in the second half of the 35th century BC.
Reconstruction of the palaeogeomorphology in the region of Mozgawa in the fourth millennium BC
A reconstruction of the paleoenvironment in the Mozgawa region during the fifth and fourth millennium BC has been previously reported (Kalicki and Biesaga, 2022; Korczyńska et al., 2019; Moskal-del Hoyo et al., 2018; Szwarczewski, 2021). It is only worth remembering that the level of the floodplain in the early Holocene was 6–8 m lower than the modern one. Additional research was undertaken in order to study in detail the relief around Mozgawa at the turn of the Atlantic and Subboreal phases, and the formation period of the Mozgawka alluvial cone. These new geomorphological data (Kalicki and Biesaga, 2022) allow defining exploited habitats and facilitate a construction of the land-use models. In the SW part of the cross-section (Figure 2), the alternating sandy and silty layers are separated by lenses of peaty silts and clayey peats. The lowest organic level was dated to the decline of the Atlantic: 5190 ± 70 BP (MKL 5734) cal. 4235-3801 BC (95.4%) and 5040 ± 80 BP (MKL 5738) cal. 3976-3651 BC (95.4%). They are covered with a layer of silt, in the bottom of which the wood was deposited at the end of the Atlantic phase and the beginning of the Subboreal: 4560 ± 110 BP (MKL 5733) cal. 3618–2927 BC (95.4%). There could be overbank deposits of anastomosing riverbeds. The beginning of ‘Mozgawka stream’ fan formation was started at the end of the Atlantic phase. It was concluded that surface runoff was possible within this small valley only when the karstic hollows located on its way were filled with deluvial/colluvial deposits, that is, some 2150 ± 120 BP (Szwarczewski, 2009, 2021). At the beginning of the Subboreal phase the range of this form reached about 600 m into the floodplain, which was accreted by it about 3 m (Figure 2). This large horizontal and vertical extension was sufficient to create a badly drained area northward of it with a water reservoir, in which gyttia could accumulate (T4/2015 core; Moskal-del Hoyo et al., 2018). The Nida River on the other hand, during the transition from Atlantic into Subbboreal phase most likely was characterised by an anastomosing riverbed with interconnected semi-remanent channels that enclosed multiple floodbasins. The higher levels of peaty silts separating the next phases of alluvial fan formation were dated to younger periods (La Tène-Roman and Medieval).

Location of geological boreholes (a) and section across the valley bottom of the Nida River on a digital elevation model (DEM) (b) (by T. Kalicki and P. Biesaga).
Reconstruction of the vegetation in the region of Mozgawa in the fifth and fourth millennium BC
At the base of hillside of the Mozgawa settlement, several cores were taken for palynological analysis. One of them (T4/2015) provided pollen records roughly for the fifth and fourth millennia BC (Moskal-del Hoyo et al., 2018). A reconstruction of the palaeovegetation based on pollen data from areas in close proximity to the loessic plateaus is of special relevance since most of the relatively close territorially cores are from ecologically different regions (i.e. Stanisławice: Nalepka, 2003; Wolbrom: Latałowa, 1976, Latałowa and Nalepka, 1987; Jęzor-Jaworzno: Szczepanek and Stachowicz-Rybka, 2004; Cholerzyn: Madeyska and Obidowicz, 2001). Therefore, the interpretation of the vegetation cover during the Atlantic phase in a regional scale (Ralska-Jasiewiczowa, 1989; Ralska-Jasiewiczowa et al., 2004; Ralska-Jasiewiczowa and Latałowa, 1996; Tobolski and Nalepka, 2004) does not represent the situation that took place in the loess uplands (Moskal-del Hoyo, 2021).
Based on pollen data of the core T4/2015 it was concluded that during the Atlantic phase in the fifth millennium BC and the beginning of the fourth millennium BC, pine forests and mixed pine-oak forests together with spruce and birch dominated the local landscape. Deciduous woodlands with lime, elm, maple and hazel, typical for the Atlantic phase in a regional scale, were present, but it seems that they did not play particularly important role in the local landscape. In the river valley, patches of alder woods grew. Also, grasslands, on both of dry and wet habitats, developed nearby. In the middle of the fourth millennium BC, small changes in vegetation cover were observed: (1) the first occurrence of human indicators was detected, which included the presence of Cerealia pollen (Triticum type) as well as ruderal/segetal nitrophilous plants; (2) there was a slight decrease of arboreal pollen, but pine-oak forests still predominated, while oak-lime-elm woodland grew on smaller surfaces; (3) alder woods occurred as well as patches of riverine forests with willow, birch, poplar and ash in the Nida River valley; (4) there was a higher taxonomic diversity among plants typical for both: drier grasslands (Poaceae, Knautia arvensis, Hypericum, Plantago media and Plantago lanceolata) and fresh/wet grasslands (Cyperaceae, Filipendula, Lychnis and Lythrum). This may suggest that forest communities became more open or their surface was slightly reduced (Moskal-del Hoyo et al., 2018). Due to erosion processes initiated during the first human permanent occupation on the hill. During the first phase of the FBC in Mozgawa the organic layer containing pollen grains was covered by mineral diluvium.
Anthracological analysis conducted on charcoal remains coming from the early phase’s archaeological features from Mozgawa site corroborated that local woodlands were dominated by pine and oak trees, while alder carrs developed in the river valley (Moskal-del Hoyo et al., 2018).
Materials and methods
Carpology
Carpological studies were conducted on 275 samples collected from 25 archaeological features, which were floated with use of 0.5 and 1.2 mm mesh sieves (Mueller-Bieniek et al., in preparation). They represent all chronological phases of the Mozgawa FBC settlement. Only charred seeds and fruits were considered for the carpological analysis (Table 1). Uncharred, fresh and partly rotten remains were excluded as recent contamination. Mineralised remains were also excluded from the analyses because the soil conditions at the studied area can naturally cause mineralisation of organic particles at any time. Taxonomical identification was based on seed and fruit morphology (e.g. Cappers et al., 2006; Jacomet, 2006; Kulpa, 1974) and was conducted with the help of stereoscopic microscopes (6×–63× magnifications). The carpological reference collection of modern seeds and fruits as well as archaeobotanical remains stored in the KRAM P collection of the W. Szafer Institute of Botany Polish Academy of Sciences (IB PAS) were also used.
Assignement of faunal and botanical remains collected from archaeological pits to particular habitats around the Mozgawa site.
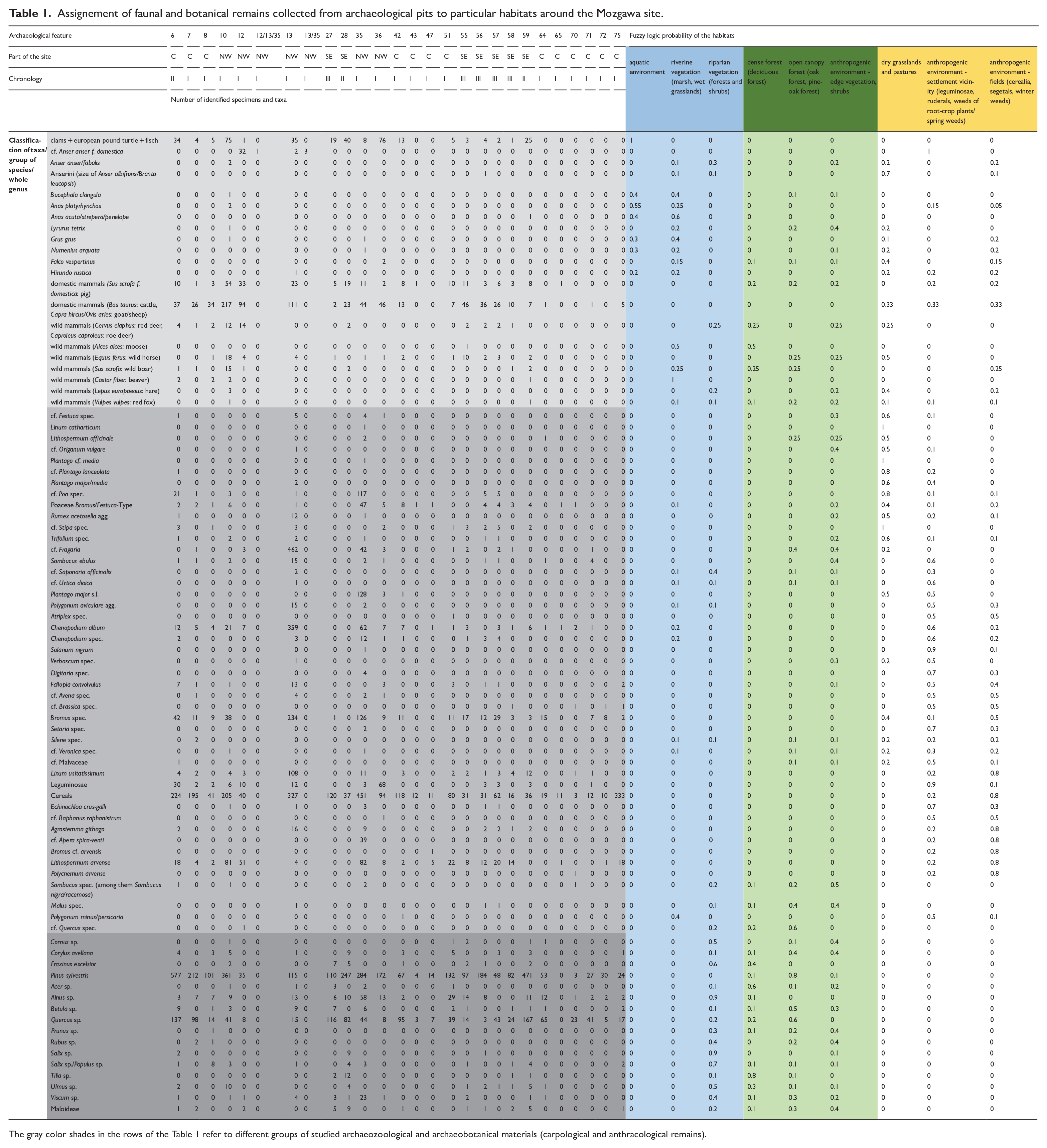
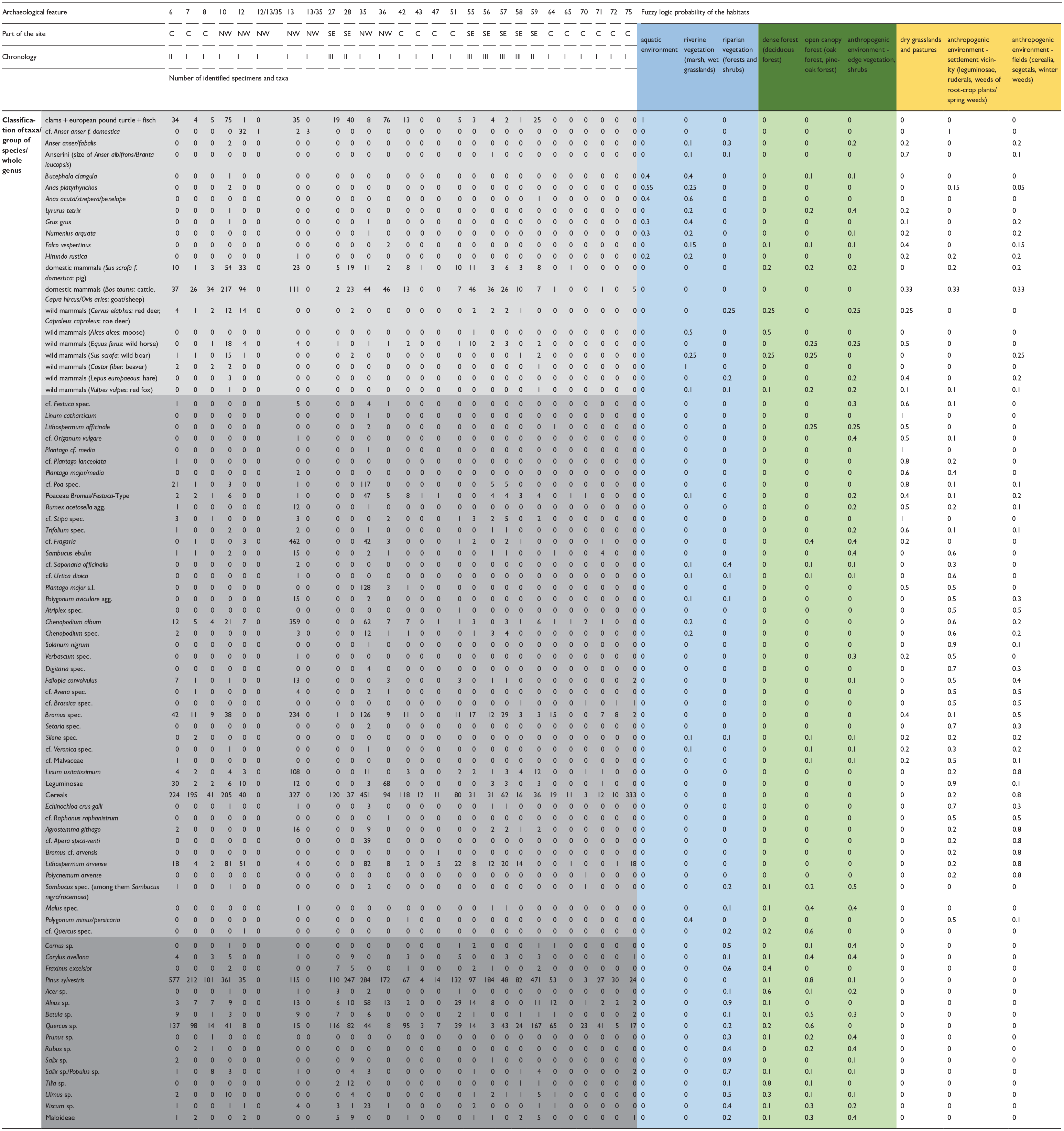
The gray color shades in the rows of the Table 1 refer to different groups of studied archaeozoological and archaeobotanical materials (carpological and anthracological remains).
Anthracology
A total of 5039 charcoal fragments recovered from 24 archaeological features (228 samples) were considered for the anthracological analysis (Table 1). They represent only scatters of charcoals found in the pit sediments. This charcoal assemblage was studied following the standard method of taxonomical identifications with the help of a modern reference collection of wood and charcoals (IB PAS), and atlases on wood anatomy (e.g. Schweingruber, 1990). The majority of Central European species are not differentiated within their genera, and taxonomic identification to species level has been given when one species exists in the present-day vegetation of the region, for example Pinus sylvestris (Lityńska-Zając and Wasylikowa, 2005).
Archaeozoology
Animal remains were identified with the help of a comparative bone collection at the Institute of Systematics and Evolution of Animals (ISEA, PAS) and bone identification manuals (Bacher, 1967; Bocheński and Tomek, 2009; Hillson, 1992; Schmid, 1972; Tomek and Bocheński, 2009). Bone surfaces were observed for modifications made by the bones depositors (e.g. humans, animals) as well as for post-depositional modifications (e.g. made by rodents or plant roots). The identified animals are expressed in number of identified specimens (NISP) and minimal number of individuals (MNI), which are standard units used in zooarchaeology (Klein and Cruz-Uribe, 1984; Lyman, 1994; Reitz and Wing, 1999). However, in Table 1 only NISP was included.
Statistical analysis
Nine probable FBC habitats were defined to examine land-use strategies in the area around the settlement (Figure 3). The main vegetation types are based on the potential natural vegetation (Matuszkiewicz, 2008; Plit and Solon, 1994). In the Nida River valley itself there are various habitats of riparian environment: (1) aquatic environment (river), (2) marsh, including mainly unforested wetlands and (3) riparian woodlands, including willow-poplar forest along Nida River channels, alder carr on swamps and oak-lime-elm floodplain forest. Other forests are divided into two main types: (4) open canopy forest with oak and pine, (5) deciduous forest with lime, oak, elm, ash and maple, which formed rather a dense canopy. Edge vegetation (6) could appear in the area between forests and the human settlement with its fields and pastures. In this zone, which represents an anthropogenic environment, mainly light-demanding trees and shrubs were growing. Since archaeobotanical studies of the Neolithic times offer no clear evidence to separate plants typical for grasslands from those that grow in the historic and pastures, these two habitats have been linked together as (7) grasslands/pastures. Other areas of open landscape that were transformed due to the human activity are divided into: (8) areas in direct vicinity of houses, on which ruderal taxa could have grown and garden-like intensive cultivation of legumes and wheat could have taken place and (9) of fields extensively cultivated cereals accompanied by segetal weeds (cf. Nowak et al., 2020).
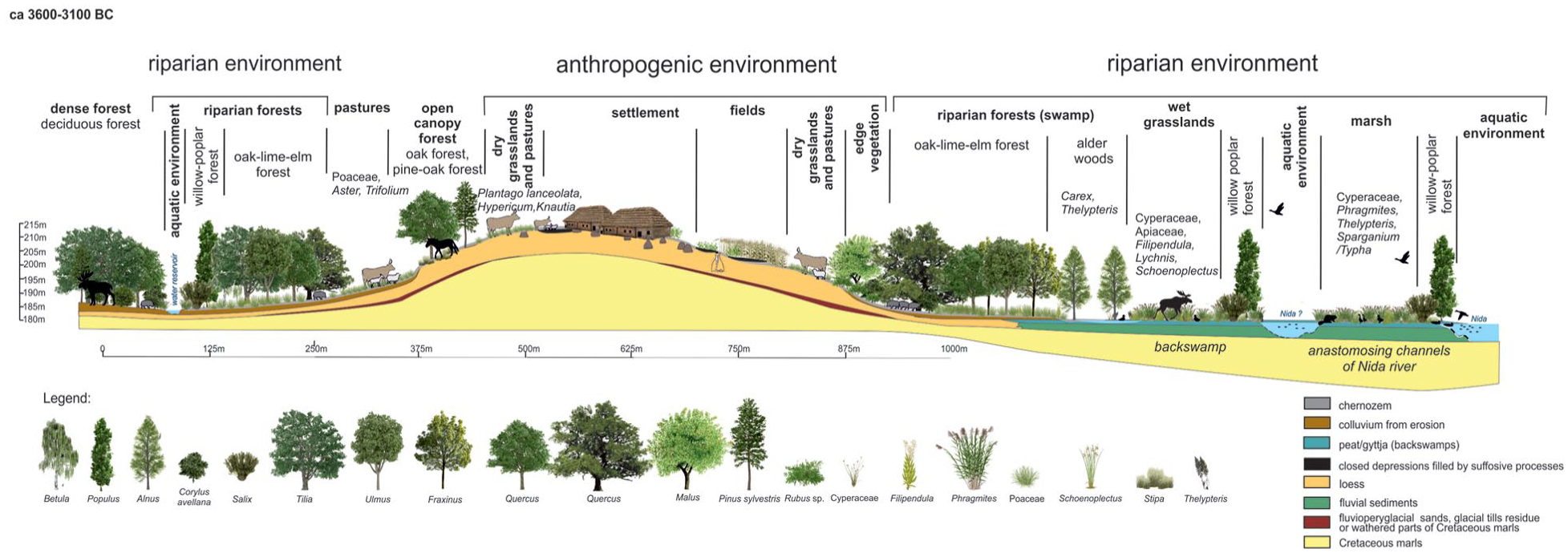
Classification of the habitats at the SE Nida Basin (after Moskal-del Hoyo et al., 2018, changed).
In the next step, ecological requirements of each botanical and zoological taxa (sometimes group of species or whole genus) were described by probability of appearance in diverse habitat types in the range of 0–1, where 0 = completely false and 1 = completely true. The fuzzy logic method was applied using a ‘theory driven’ approach in which the estimated probability is based on the knowledge and experience of experts as well as experimental research (in our case historical and ethnographical comparisons). For carpology, actual botanical information (i.e. Rutkowski, 2008; Szafer et al., 1969; Zarzycki et al., 2002) as well as palaeoecological and archaeological data were used (Kreuz and Schäfer, 2002; Lityńska-Zając, 2005; Mueller-Bieniek and Woch, 2012). In the case of cultivated plants, probable diversity of cultivation methods in a particular period of prehistory (Mueller-Bieniek et al., 2019), including garden-like, intensive cultivation of legumes and wheat (Bogaard, 2004; Bogaard et al., 2013), was taken into account. For anthracology, the ecological requirements of trees and shrubs (Seneta and Dolatowski, 2004; Zarzycki et al., 2002), a map of the potential natural vegetation (Matuszkiewicz, 2008; Plit and Solon, 1994) as well as a description of present-day forest communities with their characteristic features and structures, at the level of plant associations, were taken into account (Matuszkiewicz, 2005). The environmental attribution of birds is based on the International Union for Conservation of Nature (IUCN) database IUCN (2022), modified in three steps for compatibility with the set of the nine habitats. For every bird taxon, step one comprised listing the possible environments from the IUCN database and the removal of the ones clearly non-applicable (as tropical, marine or saline habitats). Step two consisted of translating the listed environments (from IUCN database) to the given set of nine habitats. The translation resulted in a value of 1 for every environment from IUCN database divided and attributed accordingly to a set from the nine possibilities. The attribution and the chosen set were every time based on the actual environmental requirements/preferences of the bird (derived mainly from Voous, 1960, and IUCN, 2022, habitat descriptions). Step three consisted of totting up the partial values of each of the nine possible habitats, and final apportion of the proportional values (the sum = 1) for every habitat (out of nine). In the case of taxa that represent two or more species (e.g. ‘Greylag goose/bean goose’), every constituent taxon (as Greylag goose) was treated separately, up to step two. In step three, partial values of all constituent species were totted up together. The taxa that encompass an extremely large number of possible species (as ‘Anseriformes’) were excluded from the environmental analysis. A similar method was applied to other animal taxa. Then, for archaeobotanical, anthracological and archaeozoological remains the percentage share was calculated, with regard of two occupation phases M-I and MII-III. Finally, for each taxa/group of species/whole genus in the particular phase aforementioned the percentage share was multiplied using fuzzy-logic probability of the appearance in the examined habitat. This allowed to calculate the potential habitat’s exploitation index. This could provide evidence about the intensity of exploitation of the particular habitats in terms of plant and animal palaeoeconomy.
The use of plant and animal assemblages for fuzzy-logic analysis requires some critical comments. In the case of archaeobotany, the percentage composition of the plant assemblages is heavily affected by taphonomy, as of course not all plant materials, employed at the settlement, were charred and afterwards only part of the sediment from archaeological features was analysed. This factor might be mitigated by a high number of analysed carpological and anthracological samples (in our case 275 and 228, respectively). Also, a diverse morphology of the botanical taxa might potentially deliver a different number of seeds and therefore, may lead to the overrepresentation of some taxa on site. The faunal bone assemblage of the Mozgawa site is not the result of an accidental burying of part of the entire animal material or a matter of sampling since all the bone material that was collected during the excavation was described. On the other hand, bone remains discovered during fieldwork (both in terms of the discovered species complex and the representation of skeleton elements) were directly and significantly influenced by human decisions (e.g. grazing and slaughtering animals outside the settlement area, removing selected parts of animals through separation of areas for different stages of carcass processing, such as skinning, dismembering, filleting and marrow extraction). Furthermore, various taphonomic factors (biotic and abiotic) might have affected the number of discovered remains, and that is why the preservation of animal remains was the result of complex processes – the intensity of settlement processes, taphonomy, and fieldwork methods used. For example, it is likely that the presence of dogs at the site was the reason for the discovery of fewer remains of smaller animals, including juvenile individuals, which were entirely consumed by them. Therefore, the preservation of specific animal and plant material was not mere coincidence, but it was the result of specific processes that led to the preservation of a given deposit.
Site catchment analysis (SCA) was conducted to trace the possible location of the habitats within an area of economic activities around Mozgawa. This approach relies on deriving a territory (catchment) belonging to a site by applying some geographical rule (like distance) and to analyse the properties of the catchment area, usually to see the economic benefits (e.g. agricultural yields) that the exploration of the zone of interest would bring. According to this theory, the distance from a settlement rises the efficiency to explore nearby resources. From the very beginning, the most controversial problem consisted in the optimum of setting the norms to the impact variables of the site exploration territory (Binford, 1982; Higgs and Vita-Finzi, 1972). It was assumed that the rationalisation of the economic activities should depend on environmental factors, as well as the subsistence economy, transport’s possibilities and the population’s size (Binford, 1982; Higgs and Vita-Finzi, 1972; Jarman, 1972; Jarman et al., 1982). As most ‘rational’ for agricultural and pastoral activities, an area within a radius of 1 km from the settlement was proposed. This distance has been used in several studies (compare Chisholm, 1968; Kreuz, 1990; applied e.g. by Dębiec, 2006; Dębiec et al., 2005; Kadrow, 1990; Kruk et al., 1996; Zych, 2005). In this area, cultivated fields and, in some cases, seasonally domesticated animals were grazed, while a collection of wild herbaceous and woody plants as well as a hunting of wild game took place less frequently (Kreuz, 1990). An efficiency border of agricultural activities was set at a distance of 5 km from the site (Hodder and Orton, 1976; Vita-Finzi et al., 1970: 230; Jarman et al., 1982). In the case of hunting and gathering activities, such a border was estimated at 10 km radius from the settlement (Bakels, 1978; Binford, 1982; Jarman, 1972). In following studies those distances were subdivided in intervals of 15 min, 1and 2 h of walk. Such isochrones were calculated in the present contribution with the use of a path distance algorithm (Tobler, 1993). This anisotropic formula – comprising the fact that upwards travelling is harder than downwards – calculates the time used to traverse 1 m under certain slope conditions; crossing areas with more than 12° is predefined as not possible. In order to get most accurate results within a radius of ca 1 km, the modern digital elevation model (DEM) was re-calculated according to the interpretation of geomorphological drillings. In the Nida Basin, the bottom of the valley was lowered at about 4.5 m and at the valleys of streams south and north from settlement the relief was lowered at over 6–9 m in order to reach a level from the late Atlantic phase. Also, at the eastern slope of the site a small terrace was modelled, based on the transect of drillings cross the promontory (Korczyńska et al., 2019). Although archaeological trenches may suggest a spatial shift of the location of the entire settlement through time, the results of the field survey did not support such assumptions (Korczyńska et al., 2019). For this reason, a starting point was settled in the middle of the archaeological site for the SCA.
Results
Carpological and anthracological analysis
In Mozgawa, there is a dominance of emmer wheat Triticum dicoccum among crops (Figure 4f). Barley and other crops are found rather sporadically. There are exceptionally high frequency of pulses, including both lentil Lens culinaris and pea Pisum sativum. The presence of flax Linum usitatissimum seeds is also documented (Figure 4d), as well as a large list of wild plants, including taxa associated with segetal/ruderal communities, and grassland vegetation which might suggest a mixture of intensive and extensive agriculture along with importance of gathering (Figure 4, see also: Mueller-Bieniek et al., in preparation).

Cross-sections of the archaeological features of the Funnel Beaker Culture settlement in Mozgawa (a-c); archaeobotanical macroremains (d–f): seeds of flax Linum usitatissimum (d), seeds of wild strawberry cf. Fragaria sp. (e), Cerealia and Poaceae grains dominated by wheat Triticum sp. and emmer wheat Triticum dicoccum (f), archaeozoological remains (g–h): sheep/goat mandible (g), fish vertebrae (h). Photos by: J. Ociepka and M. Korczyńska-Cappenberg (a-c), K. Stachowicz (d-f), A. Walanus (f), J. Wilczyński (g-h).
The results of anthracological analysis from M I and M II–III phases demonstrate a dominance of Scots pine Pinus sylvestris and deciduous oak Quercus sp. in wood charcoal assemblages in both phases. This was interpreted as an result of the exploitation of the open pine-oak forests growing on higher elevations of the loessic hill in Mozgawa (Moskal-del Hoyo et al., 2018). There is a slight decrease of pine charcoal in phase M II–III, from almost 70.7% to 66.9%, accompanied by an increase of oak (from 17.6% to 252%), which might reflect a wider area of wood supply, coming also from deciduous forests. The third taxon in terms of its frequency documented in Mozgawa is alder Alnus sp., which reflects the use of riparian woods, mainly alder carrs that could have developed in the Nida River valley. Other taxa are found in less frequency and each of them do not exceed 1.5% of the assemblages in both phases, although a slight increase of lime, ash, elm and Maloideae is detected in younger phase.
Archaeozoological analysis
Domestic mammals represent a typical spectrum of species, among which cattle Bos taurus remains significantly prevail in M I and M II–III phases (Wilczyński et al., 2021). They constitute around one-third of the whole bone assemblage. This share is lower if we compare MNI, but still is visibly higher than other taxa. Remains of pig Sus scrofa f. domestica and goat/sheep Capra hircus/Ovis aries are comparable in number, but in terms of MNI, pigs are more numerous. According to Gillis et al. (2017), the cattle age profile is characteristic for meat exploitation. Obviously, a breeding strategy focused on meat is observed in the case of pigs. The interpretation of goat/sheep age profiles is unclear, due to various products that goat and especially sheep breeding offered: meat, wool, milk (Figure 4g; Pipes et al., 2014). Generally, with time, the share of the main domestic mammals does not change substantially. The ratio of the main breeding species including cattle, goat/sheep and pig is similar, and the observed changes are insignificant, for example, in phase M II–III we can observe a slight increase of pig remains, which takes place at the expense of smaller ruminants.
The wild animal bone assemblage is very diversified. A share of these remains among all animals is 6.6% (9% if we take into consideration mammal remains). Among wild animals, taxa associated with aquatic environment, including freshwater mussels, different species of fishes (e.g. Lota lota, Esox lucius, Rutilus rutilus and Perca fluviatilis) (Figure 4h), European pond turtle Emys orbicularis, waterfowl (geese Anser anser/fabalis and ducks Anas platyrhynchos), crane Grus grus, Eurasian curlew Numenius arquata, and some mammals (beaver Castor fiber) were present. This proves the intensive exploitation of water resources, being an important part of subsistence practices. The difference is visible in the intensity of water resources use. While in the phase M I the share of fish remains accounts for about one-tenth, in phase M II-III this share is more than one-fifth.
Among wild mammals, species inhabited different environment (forest and open areas) indicate the presence of a mixed landscape in the nearest surroundings of the settlement. The share of wild mammals in faunal assemblage is generally similar. It amounts from 7% for phase M I to 8.7% in phase M II-III. However, the differences are visible in the species composition of this fauna. In phases M I and M II-III the number of remains of mammals inhabiting woodland and riparian areas (e.g. reed deer and roe deer) is similar, but surprisingly in phase M II-III the share of horse remains inhabiting the open canopy forests or/and open areas (dry grasslands/pastures) almost doubles.
Statistical analysis of the habitats
A graphic illustration of the fuzzy logic technique is presented in six pie charts that represent habitats, which were exploited for animal and plant management (Figure 5). The results seem to illustrate land-use in a coherent way. The percentage share of the habitats do not necessary correspond to their spatial size, but it offers evidence that allows to trace some preferences of the exploited habitats and the importance of different ecological zones. In general, in the archaeozoological and archaeobotanical diagrams, a composition of animal and plant taxa suggest an exploitation of very diversified zones, in which anthropogenic, transformed landscape, open forests and riverine areas might have the highest significance (Figure 5).
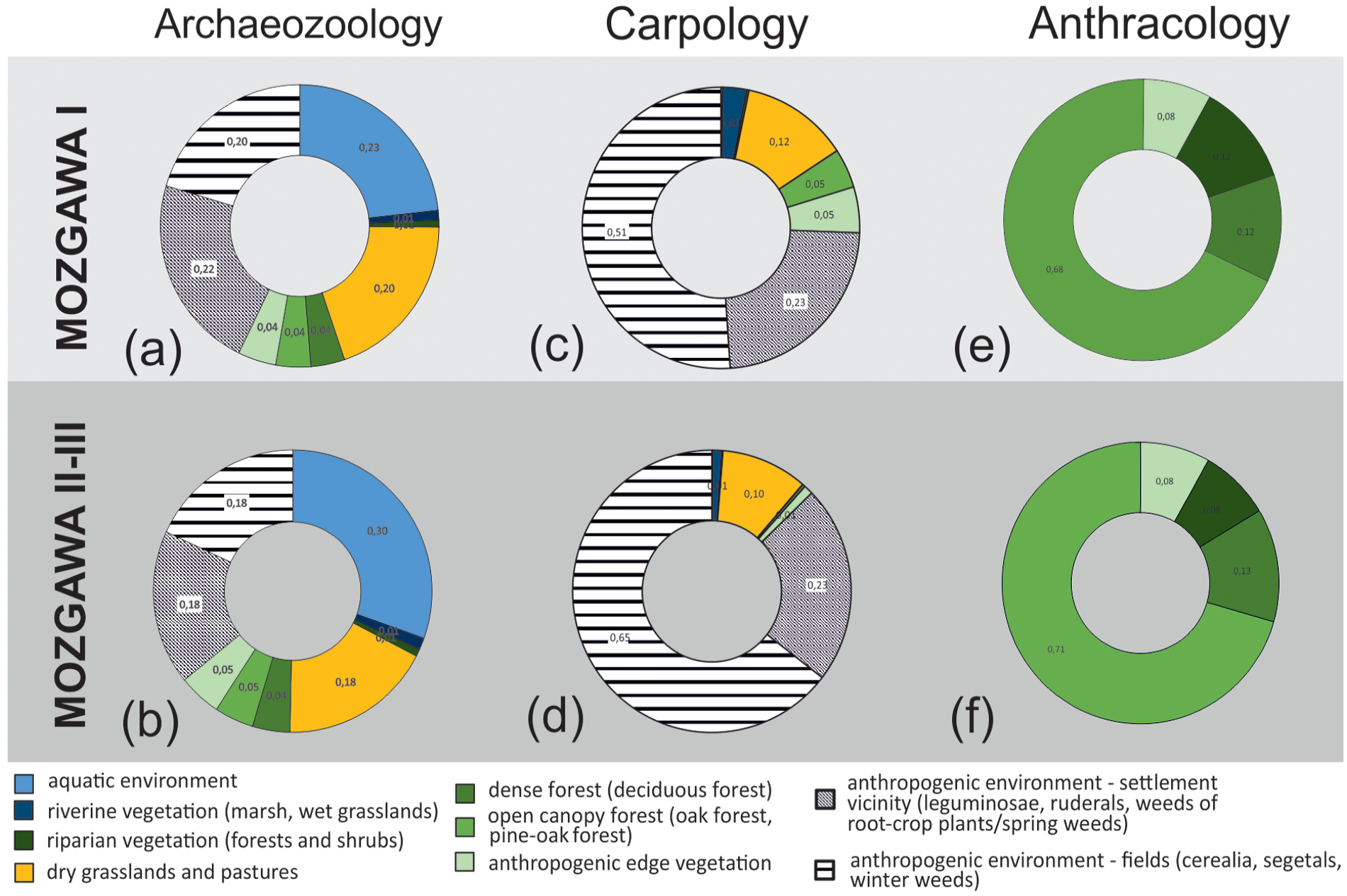
Diachrone composition of potential habitat’s exploitation indices presented as percentage share, based on faunal (a,b) and botanical remains (c–f).
Diachronic analysis of the data do not show any major changes in exploitation of habitats during the development of the settlement. For instance, in both carpological diagrams, crop fields and areas in direct vicinity of houses are habitats that were most intensively exploited and a share of such transformed areas seems to grow in time. The importance of this type of habitats is obvious as it confirms the crucial role of agriculture for food supply in the local FBC. It is worth emphasising that caryopses of cereals are quite equally distributed in the analysed samples (pits), and there are no deposits of charred remains that might increase the amount of error in the results (Table 1). Plants typical for grasslands and pastures demonstrate the relevance of these habitats. Other habitats were less represented, including open forests, edge vegetation and riverine environments, although aquatic plants were completely missing. The lack of aquatic environments in carpological diagrams can be connected with taphonomy and the lower probability of carbonisation of aquatic plants as well as lacking data connected with the use of vegetative parts of plants in carpological assemblages.
The main type of exploited forests for fuelwood was open canopy forest dominated by pine and oak trees (Figure 5; Table 1). Based on a high frequency of pine and a relatively low frequency of all decidous trees and shrubs, other than oak and alder, the importance of other forest types, such as deciduous forests and oak-lime-elm forests, played probably minor importance. This is likely due to the greater distance of these forest communities to the settlement, which means that for a gathering of the fuelwood an ease of its collection and transportation to the settlement was an important factor determining wood supply strategy (e.g. Asouti and Austin, 2005; Chabal, 1997). The only more conspicuous change in time is a decreasing role of riparian habitats (Figure 5). This might be related to a minor share of charcoal fragments of Alnus sp. (from 6.5% to 2.4%) found in the younger phase, which in turn might be correlated with the lack of archaeological features dated to phase M II-III from northern areas of the settlements, those adjacent to the Nida River valley.
In the case of faunal remains, there is also a lack of major changes in habitats’ exploitation during the development of the settlement. Only the role of aquatic environment seems to be more significant in the younger phase (increase from 0.23 to 0.30). Nevertheless, both diagrams underline the importance of the rivers around Mozgawa for hunting (fishing) activities.
SCA allows to estimate the exploited zone of 4.2 sq km in the case of isochrone of 15 min, 67.3 sq km in the case of 1 h and 265.9 sq km within an area of 2-h walk distance (Figure 6). Interestingly, almost half of the 15-min catchment area are backswamps along the Nida River and deep valleys around the promontory. In the case of 1-h distance, deep valleys of the rivers constitute almost 40% of the zone, and in the catchment area of 2 h almost 30% of the zone. The aforementioned potentially flooded areas are located in the eastern parts of the catchments, while the western part is characterised by hilly terrain with deep river valleys cutting through it.
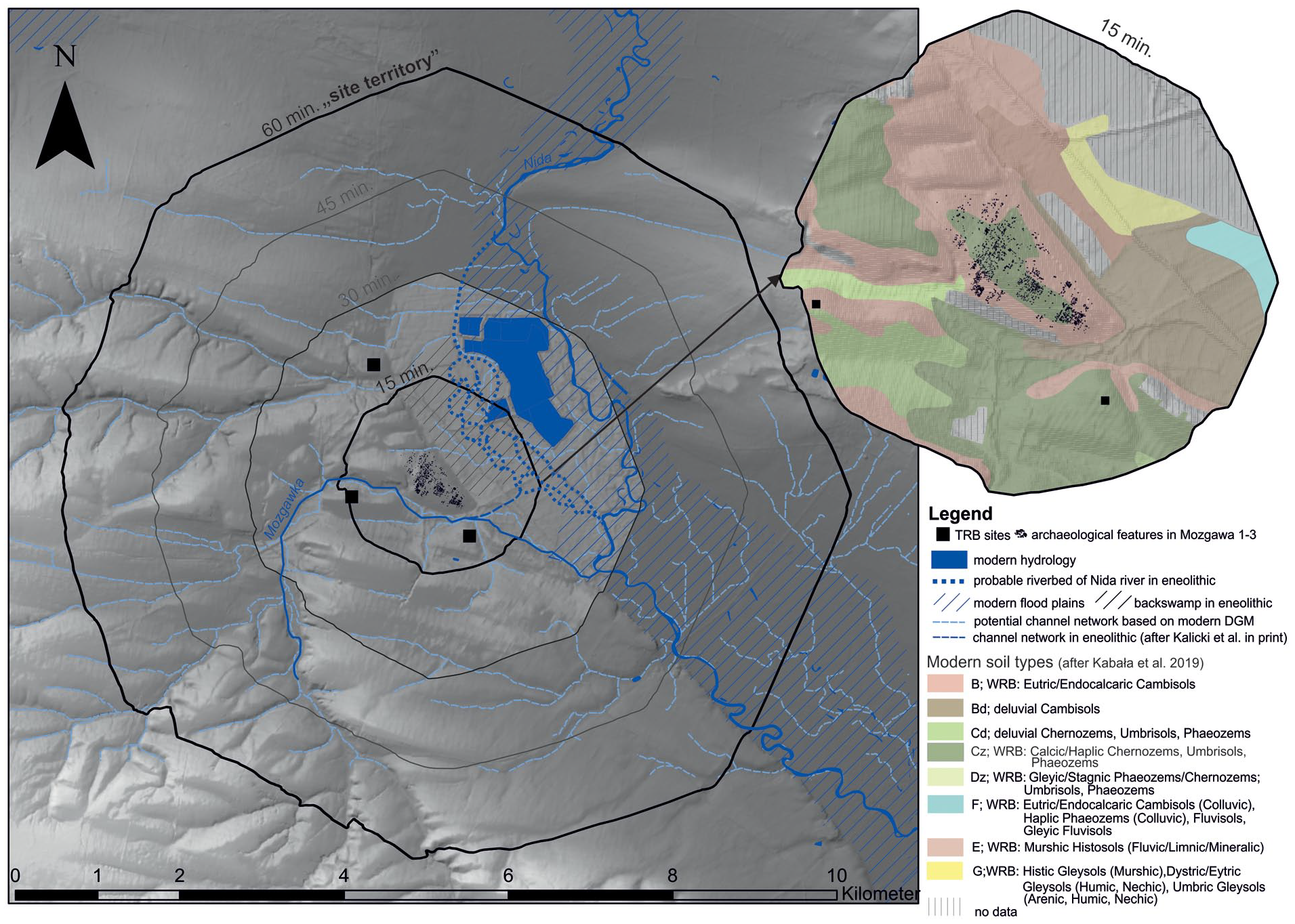
Site catchment analysis of the Funnel Beaker culture settlement in Mozgawa.
Discussion
The Neolithic agricultural communities triggered soil erosion at the loess areas. This process is observed locally in the Nida valley, where mineral deluvia sedimentation observed at the core T4/2015 right next to the FBC settlement started around 4890 ± 40 BP, ca. 3770–3630 BC. Obviously, this is not a surprise. Analogous sedimentation changes also were noticed in the river valleys adjacent to the Mozgawa area to the south-west in the fourth millennium BC (Kruk et al., 1996; Michno, 2004). Interestingly, at the base of the edge of the Vistula River the accumulation of thick loess colluvium, interrupting the organic accumulation, started slightly earlier (ca. Second half of the fifth millennium BC) (Godłowska et al., 1987; Wasylikowa et al., 1985). In the bottoms of upland tributaries of the Vistula River the accretion of the organic overbank sediments was diachronic, several 100 years earlier in their upper reaches (ca. 4000 BC) than in the lower ones (ca. Second half of the fourth millennium BC) (Alexandrowicz, 1988, 1997; Kalicki, 2006; Pazdur and Rutkowski, 1987; Rutkowski, 1984; Rutkowski et al., 1988; Rutkowski, 1991).
A wide spectrum of plants and animals together with the existence of diverse habitats that were potentially exploited are observed in both occupation phases of the FBC settlement in Mozgawa. Although the majority of plant and faunal remains represent cultivated species and livestock, there are also taxa clearly reflecting food collecting from natural environments, suggesting gathering, hunting, and fishing. This might be interpreted in terms of a food supply strategy based on multiple resources, which could have been applied to avoid hunger crisis. It is likely that such a strategy played the main role in the selection of this place to settle down, especially if we consider that the major area of the site was occupied in the early phase and was also exploited for hundreds of years. The evidence of exploitation of the river valleys shows the relevance of a location of such a large settlement in close vicinity to the riparian environments. The decision of building a settlement at a spot that allows easy access to the diversified habitats and their resources seems to be very reasonable.
The method of identifying the ecological zones that were exploited by the inhabitants of the settlement used in the present study has essentially no counterparts in the literature on the southern FBC groupings. It is therefore difficult to refer to related publications and make comparative analyses. It is perhaps only appropriate, due to geographical proximity and similar (but not identical!) conditions, to cite the classic study made for the nearby, large FBC settlement at Bronocice, despite its nearly 40-year age (Milisauskas and Kruk, 1984). However, it must be stressed that its main purpose was to reconstruct settlement organisation and palaeodemographic simulations; studies on the land-use model were rather secondary. It is undoubtedly worth emphasising that the cited study indicated that the FBC in the Bronocice microregion was associated to the highest degree with the zone of the edges of the upland, which is also characteristic for Mozgawa. In a way also similar to Mozgawa, the habitat differentiation of the settlement surroundings is evident. Within the area delimited mechanically by a 1 km radius around the Bronocice settlement, the alluvial zone plays the largest role; it covers nearly 32% of the area. This share is therefore of the same order as in Mozgawa, reconstructed on the basis of archaeozoological data and SCA. The other zones delimited by the cited authors, occupy correspondingly lower portions of the area: valley edge − 14%, slopes of the uplands − 14%, edges of the uplands − 19.7%, and uplands − 14.6%. The other larger settlements in the Bronocice micro-region that were over 5 ha in area, were also located in the ‘edges of uplands’ zone. In their 1 km surroundings, the alluvial zone is also of considerable importance (ca. 20%), but is inferior to the ‘slopes of the uplands’ and ‘uplands’ zones (ca. 30% each).
In order to better understand the agricultural model of the FBC community in Mozgawa for a site catchment within 15 min distance (corresponding to the so-called ‘agrarischer Nutzungsraum’ after Kreuz, 1990) an attempt was made to estimate the possible location of the analysed habitats. During the decision-making process, factors such as relief, hydrological conditions, type of soils and potential natural vegetation were taken into consideration. In the case of cultivated fields, a short-distance visibility from the settlement was considered as well (Figure 7). In this predictive modelling, we suggest that plots of legumes that were intensively cultivated could have been located in the direct vicinity of the houses (Figure 8). In turn, flax, a plant that requires moderately moisture and fertile soils, in general can grow on ploughed fallow lands, pastures, and perennial meadows (Heller, 2012: 10), so it could have been cultivated on the north-eastern loess slopes (Figure 8). In turn, cereal plots might have been located at the promontory in the building-free areas, in the sunny south-eastern and southern slopes of the site at the terrace that is well visible from the middle of the site on the north-eastern slope and at the southern slopes of the small hillside in the north-western vicinity of the settlement (Figures 7 and 8). In general, this assumption corresponds to the theoretical approach, claiming that the fields to be cultivated should be located about 10–15 minutes’ walk from the settlement (Kreuz, 1990: 153–157; Hochuli et al., 1998: 224; Posluschny, 2010, 317; Posluschny et al., 2012: 417). Such a hypothesis is supported after performing analysis of malacofauna. The presence in the archaeological pits of Mediterranea inopinata, a xerophilous species that lives in cultivated soils, indicated an agricultural land-use in areas located nearby the households (Alexandrowicz et al., 1997; Moskal-del Hoyo et al., 2018). A recent study of the stable carbon (δ 13 C) isotopic compositions of carbonised cereal grains, also including specimens from Mozgawa, showed that the FBC plots probably cover areas with lower water availability than in earlier periods of the Neolithic or were more open (the level of deforestation was higher than before), supporting the ideas expressed already in the early 1970s by Janusz Kruk, who stated that the Middle Neolithic agriculture spread into new, more elevated areas (Mueller-Bieniek et al., 2019). The general variability of Middle Neolithic δ 15 N and δ 13 C values obtained from emmer wheat grains derived from Lesser Poland sites suggest rather variability of cultivation methods than one specific model. Nevertheless, at all studied sites from southern Poland field manuring was probably practiced as suggested by δ 15 N values (Mueller-Bieniek et al., 2019). Only four emmer wheat samples from the Mozgawa site were studied and the results were generally very close to the ones obtained from Early Neolithic measurements in that area (δ 13 C VPDB: −24,7; −23.5; −24.2; −23.5‰ and δ 15 NAIR: 6.0; 6.9; 7.0; 6.4‰ respectively). The rather high δ 15 N values, exceeding 6‰, after correction for the effect of charring (Nitsch et al., 2015) indicate permanent, well manured crop fields (Bogaard et al., 2013, 2016; Mueller-Bieniek et al., 2019).
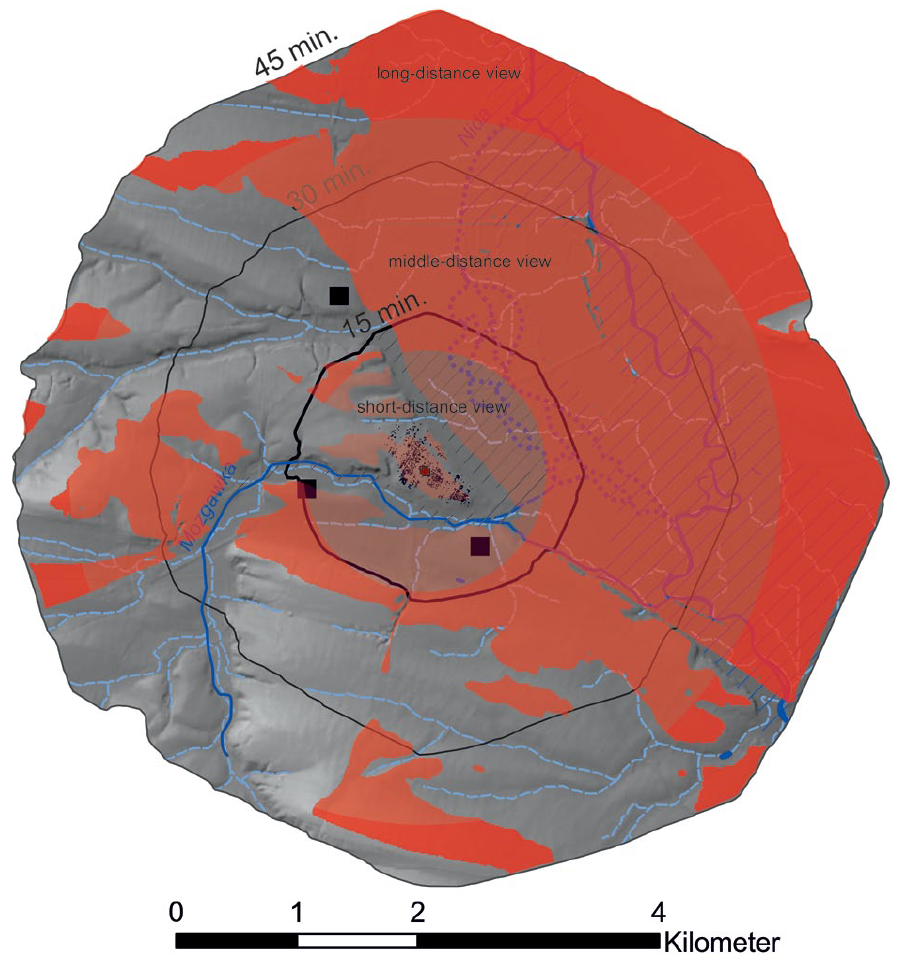
Viewshed analysis compared with the site catchments distances of 15, 30 and 45 min of walk. Observer location in marked by the red cross. The height of the transmitter (observer) - the height of the observation point above the surface at the transmitter position is defined as 1.7 m. The height of the receiver (plants and animals) – the height above the surface that is evaluated at each grid node is defined as 1 m. Short-distance view is modelled for a radius of 1 km and middle-distance view for a radius of 3 km (after Higuchi, 1983).
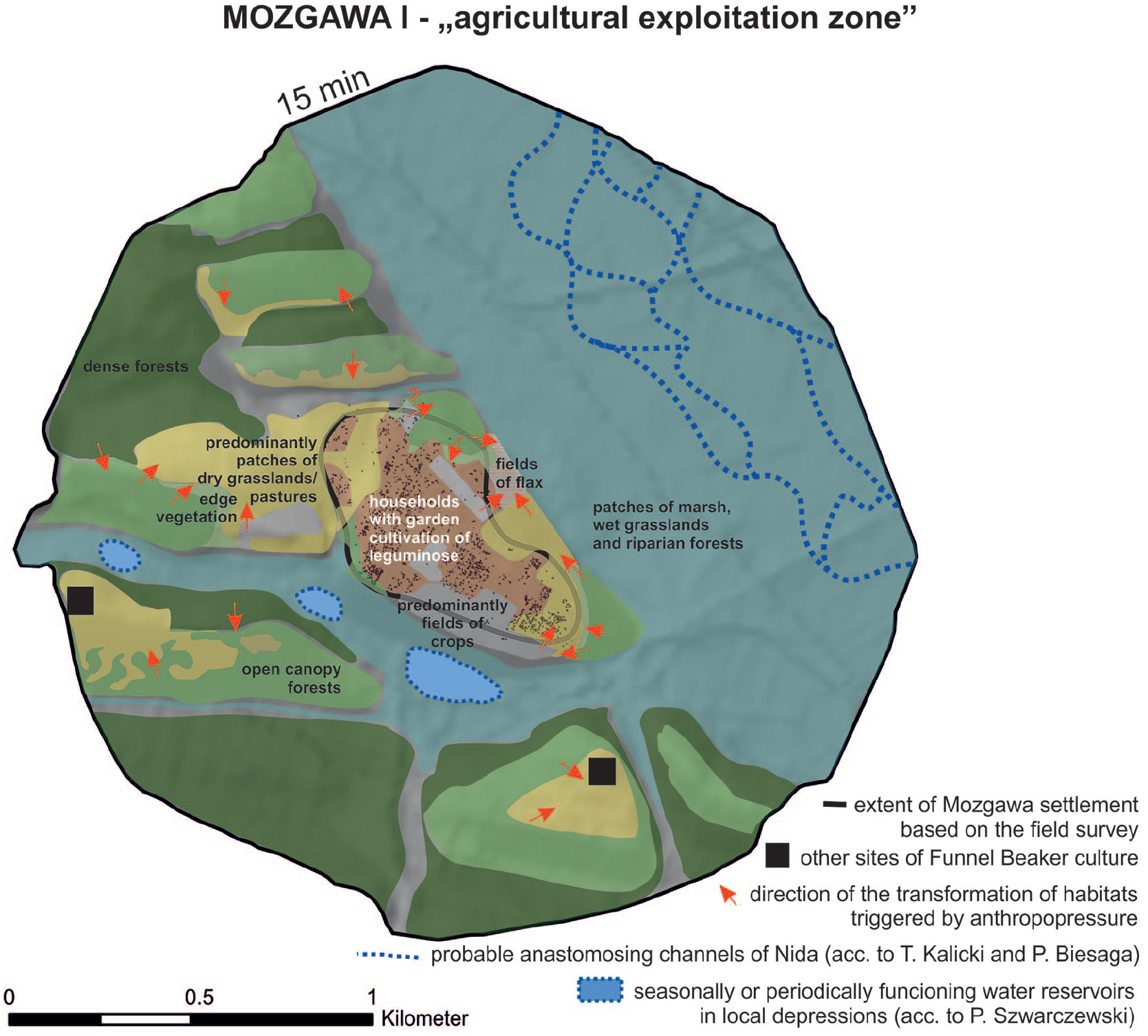
Outline of hypothetical location of the exploited habitats within the site catchment of 15 min distance (ca 1 km). Arrows illustrates directions of transformation of the habitats due to the human impact.
As a consequence, in the case of hilly loess areas in Mozgawa at least three scenarios of cultivation could be taken into consideration: (1) the majority of the crop plots were permanently cultivated, (2) some fields with a short-time cultivation was combined with an utilisation of the abandoned plots as pastureland for cattle, goats/sheeps, which prevented the re-establishment of trees and shrubs or/and (3) long-term fallow phases of some areas made possible the partial return of the forest (Rösch, 2012: 249).
In the case of the first scenario, manuring would result in quite high yields, so most of the crop fields would be relatively small, located in the close vicinity of the settlement. Nevertheless, at some areas far apart from the settlement due to a tillage with a hoe plow, a short-term cultivation followed by grazing on the fallow lands could have existed. Plots would eventually resemble grasslands because of a variety of grasses and herb species that grew between the cereals (Poschold, 2017). A high index of grasslands/pastures in the potentially exploited habitats can be multicausal, as the diasporas of grassland vegetation could have originated from fodder and dung as well as reflect taxonomic diversification of the extensive cultivated fields. Though stable nitrogen (δ 15 N) and carbon (δ 13 C) isotopic analysis clearly indicates permanent cultivation, opening of the landscape visible in the pollen data, decreasing intensity of exploitation of forests for gathering of wild plants in the younger phase, as well as evidence of flax cultivation on follow lands and increase of share of the wild animals, such as geese Anserini (size of Anser albifrons/Branta leucopsis) and wild horse (Equus ferus) that are associated most of all with open areas suggest that in some areas adjacent to the Mozgawa settlement, the second model could be justified as well. On the other hand, the third scenario, which is characteristic for a shifting extensive cultivation in the forest that results in extremely high yields and therefore requires small fields (Rösch, 2012: 249), could be supported by a constant high percentage share of open canopy forests (Figure 5e and f). However, there is no clear evidence that could support this hypothesis like, for example, an increasing number of pioneer trees such as birch that rapidly colonise open ground especially in secondary successional sequences following a disturbance or fires. This was also not reflected in the palynological record, where birch pollen reached consistently low values (up to 5%–6%), slightly lower than the regional average (10%; Ralska-Jasiewiczowa et al., 2004), confirming the sporadic occurrence of birch trees at some distance from the drilling site. In anthracological samples, the share of birch in both phases is about 1% of the assemblages. The above-mentioned presence of open canopy forest might also be related to an enlargement of the fuelwood collection zone, from 15 min walk to a longer distance (compare below). Usually, firewood collection took place in local and easily accessible woodlands (e.g. Asouti and Austin, 2005; Chabal, 1988, 1997; Kabukcu and Chabal, 2021). However, forests clearing, that occurred near the settlement for fields and/or pastures might result in a larger area of wood supply, located in longer distance from the site and should have taken place in various woodland types. Forests were also cut for construction of the houses, which required a use of timber. Although wood quality certainly played a role in this aspect, the cost/benefit relationship was a determining factor for a choice of the exploitation areas. The third scenario would trigger natural reforestation, but also eventually result in changes of the forest character and structure (old and strong trees in particular can become scarce) and led to limitation of timber available in the forest. Similar as in the second model, deficiency of forests in the direct vicinity of the settlement, that could deliver valuable woods for constructions, would eventually result in an increase of the exploitation zone to the 1 h walk distance (‘site territory’ in German ‘wirtschaftliche Nutzungsraum’ after Kreuz, 1990).
In theory, it is possible that in Mozgawa, depending on current demand and capacities, aforementioned agricultural models operated in parallel. Similar conclusions, for example, were reached by the authors of studies on Neolithic (ca. 4300–2400 cal. BC) landscape of the Central European lake dwellings, that based on off-site and on-site data (macro and micro, botanical and zoological remains). They show disagreement between these two types of sources, that is, while off-site analysis indicates slash-and-burn cultivation, on-site data gathered in the settlements indicated a general dominance of permanent cultivation of crops (wheat and barley) (Jacomet et al., 2016). Nevertheless, it should certainly be that a cessation of accumulation of organic layers and the beginning of mineral deluvia sedimentation visible in the deposits of T4/2015 core are correlated with an early occupation phase of the settlement (Moskal-del Hoyo et al., 2018). This would imply that there had already been a fairly significant and sustained opening of the landscape since the 36th century BC. Consequently, a hypothesis can be put forward that the first and second model were practiced, possibly with territorial advantage of the latter one.
The anthracological data suggest that open canopy forests dominated by pine and oak could have prevailed on the loessic area. This kind of forests in the second half of the Atlantic phase might have been the most widespred on loess regions of southern Poland, as it was demonstrated by anthracological analyses conducted on multiple archaeological sites from these regions (Moskal-del Hoyo, 2021). The constant use of this kind of woodland favoured the maintainance of open character of forests, which in turn favoured a regeneration of pine and oak. The high similarity between early and late phases might also be a result of ubiquity of oak trees, which played important role not only in open canopy pine-oak forests, but also in more dense deciduous forests, where oak could grow together with lime, maple, ash, elm and hazel, and in oak-lime-elm forests that could have developed in wetter conditions. Figure 8 shows a potential location of these forests communities, but in time human activities based on fuel and wood collections as well as above-mentioned enlargement of areas of fields and grasslands could have eventually changed the location of these forests. For example, making dense woodlands of more open canopy character. These observations are not confirmed by the analysis of the livestock remains, which shows a lack of changes. However, this may indicate that in the vicinity of the settlement there was a mosaic-like vegetation, composed of open forests and grasslands, throughout the whole period of the site occupation. Concerning the importance of the landscape for domestic animals, it should be added that while grasslands were mainly used for pasture of herds of cattle and goats/sheeps, maintainance of open forests with oaks was relevant for grazing of pigs.
The presence of wild animals at the settlement, including red deer Cervus elaphus, might also indicate a development of woodland (Apollonio et al., 2010; Niedziałkowska et al., 2021). Also, wild boar Sus scrofa and moose Alces alces could have been hunted in the forests, although the latter one rather in a dense and/or riparian forests. In turn, the remains of wild horse Equus ferus and a single remain of a geese Anserini (size of Anser albifrons/Branta leucopsis) in the faunal assemblage might suggest exploitation of open canopy forest or/and open areas, such as steppes and grasslands for hunting activity. Increasing of those taxa in phase M II-III is noticeable, however, in our opinion it is difficult to infer about the transformation of the entire surroundings of the settlement on the basis of generally sparse remains of the horse. In our opinion such a result can be interpreted, for example, as the intensification of wild horse hunting. This species became almost extinct in the Mid-Holocene because of the afforestation and loss of open habitats (Sommer et al., 2011). However, locally it persisted in Europe, and this was demonstrated in the case of the horses’ remains from Mozgawa. Their DNA analysis showed that they belonged to an older horse population (Librado et al., 2021). These data corroborate the assumption that in the ‘site catchment zone’ of the Mozgawa settlement, naturally open areas and/or open canopy forests existed and were exploited in terms of wild animals’ supply. Hunting in these kinds of habitats, although including also human-made edge vegetation and open landscape, might be inferred after the presence of a hare Lepus europeaeus and a red fox Vulpes vulpes remains. However, in the case of hunted animals it is very difficult to estimate the distance from where those were acquired.
The relevance of the aquatic habitat, that probably covered almost a half of the agricultural exploitation zone (Figure 8), is especially evident based on faunal remains related to the river. As mentioned before, clear evidence of fishes, molluscs and birds related to water environments in a archaeozoological dataset is a unique case in the FBC sites in Poland (Wilczyński et al., 2021).
The importance of riverine habitats for FBC societies was observed not only in the case of the Bronocice settlement (Milisauskas and Kruk, 1984), but also is indicated by the strong relation of settlement clusters to the river valleys found in south-eastern Poland (Król, 2018). Riverine habitats delivered food resources, that diversified dietary strategies, as in Middle Neolithic Mozgawa molluscs, which were used as food for both humans and domesticated animals. A significant increase of fish remains in phase M II-III, with a relatively similar proportion of clam remains in both phases, may indicate increasing the role of fishing. Beside direct consumption, some fishes were most likely not only dried, but also processed by a fermentation to preserve them for a long time (Wilczyński et al., 2021: 56). Profitability of such farsighted storage strategy might be one of the reasons of the increase of the number of fishes in the younger stages of the settlement. In this context, the scarcity of aquatic plants in the archaeobotanical data might not surprise due to the dominant type of preservation by charring and possible use of vegetative parts not reflected by carpological data (compare above). Among the plant macro-remains from core T4/2015 from the phase correlated with the onset of FBC settlement, aquatic and marsh plants were found, mainly associated with rush communities developed near the shore and within the shallow water body (M4-2 L MAZ, 470 −440 cm; det. R. Stachowicz-Rybka). They were represented by Schoenoplectus tabernaemontani, Schoenoplectus lacustris, Eleocharis palustris, Cyperus fuscus, Ranunculus sceleratus, Sparganium erectum, Alisma plantago-aquatica and Typha sp. (Moskal-del Hoyo et al., 2018). Some of them could have provide edible parts, such as rhizomes, young stems, could be used as green vegetables or plaiting material Schoenoplectus lacustris, Typha sp. or Phragmites australis (found as pollen; Kubiak-Martens, 1996, 1999; Kurzawska et al., 2010; Lityńska-Zając and Wasylikowa, 2005). It is probable that some minor changes of the riparian environments could have occurred, especially that the aquatic environment experienced an increase in taxa diversity in the younger phase, at the same time that alder forest (generally occupying limited areas in the neighbourhood according to pollen data) experienced a possible decrease as suggested by the anthracological data. As stated before, the reason for the decrease of Alnus in the charcoal assemblage and thereby decrease of riparian forests in the potential habitat’s index can be possibly found in the location of houses in M II-III phase, which in younger period were absent in the area facing the Nida River valley. It should be added that alder carrs are difficult to penetrate, especially during a high water level, and wood gathering from this kind of community is usually limited to the marginal areas or to periods of drying up or freezing of the sodden ground. On the other hand, a lower share of Alnus in M II–III might reflect some additional changes in local environment, which perhaps might be connected with some movement of the Nida River channel, which could have resulted either in its greater proximity to the settlement or its longer distance with dried up the closest areas. Despite the reason, these changes might explain a decrease of alder woods in the close proximity of the hill.
Unfortunately, palaeoecological data available from the above-mention core T4/2015, reflecting local changes of water level at the base of the hillside contained organic layers that corresponded to the period prior the Neolithic occupation and to the older phase of the FBC settlement (M I). The malacological analysis showed five phases based on a dominance of either terrestrial or water species, which illustrated changes of local environmental conditions. The malacofauna of the fourth phase (det. W.P. Alexandrowicz, F-IV) represented the upper part of the organic deposits. It is correlated with the second local zone distinguished by the pollen analysis, and thus with the early phase of the Neolithic occupation (M I). The snails documented in F-IV reflected important humidity growth as a share of water species exceeded 90% of the entire assemblage from this phase. This data indicates that during the development of the early phase of the settlement (M I) water levels were relatively high near the hill, forming good environmental conditions for riparian vegetation, including mosaic of rushes and alder woods growing along the Nida River.
The importance of palaeoenvironmental studies combined with palaeoeconomical data obtained for the Mozgawa settlement should be underlined, as the exploration of activity zones’ is calculated in relation to various natural resources, as the analysed dataset contains both, faunal and botanical remains. Based on this investigation we may address positively our research questions: (1) Various potential habitats from where plant and animal resources were taken were characterised. (2) Beside slight increase of the utilisation of the aquatic environment there is no evidence of significant changes in the intensity of exploitation of these habitats in terms of plants and animals resources between early and late phases of the development of the settlement. (3) There were very similar patterns of land-use during both basic phases of the settlement, consisting of exploitation of various habitats and relying the subsistence on domesticated plants and animals with a complement of wild species. Several centuries of existence of the settlement, with no obvious symptoms of any breakdown in the social structures and economic development, might indicate a high efficiency and stability of this land-use model, being likely effective to ensure food supply to its numerous inhabitants.
It is still a conundrum, why the settlement in Mozgawa disappeared, but based on the available data we cannot demonstrate that some shortage of food supply or some overexploitation of the habitats took place. In general, during the period c. 3100–2800 BC southern Poland was infiltrated and culturally transformed by societies which archaeologically have been denoted as the Baden and the Globular Amphorae cultures, and finally by the groupings of the Corded Ware culture (Nowak et al., 2022). Materials of the Baden culture were identified in the later phase of the settlement at Mozgawa. This may reflect the first changes associated with the appearance of the new cultural models and trends, which led to the disappearance of the settlement.
Conclusion
The combination of carpological, anthracological and archaeozoological data allows to demonstrate a mosaic of potential habitats from which plant and animal resources came. It seems that every habitat that might have existed around the settlement was exploited. For agriculture, plant gathering, livestock, and wood supply, fertile hilly areas around the settlement, at the settlement itself, as well as in the western part of the ‘agricultural exploitation zone’ were the most significant. In these areas, FBC settlers cultivated wheats (mostly emmer), legumes (pea and lentils), and flax, within settlement mostly on permanent fields. Outside the settlement new areas were continuously adapted for cultivation. The fields established there were most probably cultivated for a short time and then transformed into grasslands and eventually used as pastures. Wet grasslands, marshes and rivers were on the other hand highly important in terms of fishing and hunting activities. Moreover, for hunting and forest exploitation most probably a ‘site catchment area’ of about 60 min distance from the settlement was the most significant. Interestingly, diachronic analysis of the plant and animal assemblages did not show big changes in natural resources’ exploitation during the development of the settlement. This leads to the hypothesis that the land-use patterns consisted of uses of all possible types of food resources coming from various environments. Such patterns secured a stable and long-term supply of food and, as a consequence, ensured a duration and persistence of a large settlement in the fourth millennium BC. The long-term anthropogenic impact triggered constant changes in the environment and transformation of the habitats, but there are no signs of their overexploitation.
Footnotes
Acknowledgements
The Authors are grateful to Dr. Julio M. del Hoyo-Meléndez for proofreading the final manuscript.
Author contributions
Marta
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was financially supported by the National Science Centre (Poland) in the framework of research project number DEC-2013/11/B/HS3/03822 and also by the Statutory Research Tasks of the W. Szafer Institute of Botany (PAS).