
Abstract
Both modern and historical data show that lake systems are undergoing profound changes because of human impacts from local to global scales. Research on this topic shows that human pressure on water systems can take various forms and is highly variable in time and space. This article presents the results of Ostracoda fauna research as an indicator of anthropogenic changes covering the time span from 7000 BP to the Mediaeval Period. The palaeoecological information (abundance and habitat preferences of Ostracoda assemblages) obtained in studies of fossil Ostracoda fauna communities amplified by palynological and archaeological data allowed to answer the following questions: (1) to what extent the impact of successive archaeological settlement cultures caused ecological changes in the lake catchment areas without directly affecting the lakes themselves, and when anthropogenic pressure caused transformations in the functioning of lake ecosystems and (2) whether the neolithization, as the first clearly legible stage of anthropopression, had a decisive influence on the functioning of lake ecosystems. Results of Ostracoda fauna studies have been compared with archaeological data available in the literature and multiproxy paleoenvironmental data previously published in a study of lake sediments in the Chełm Hills in South-Eastern Poland. Presented results of study of Ostracoda in lake sediments as a bioindicators of anthropogenic impact suggest that the pressure of prehistoric settlement in older phases (up to the early Middle Ages) was most clearly reflected in the landscape, while the functioning of lake ecosystems was disturbed only by the periodic increase in lake trophic status. A substantial change resulting from the analysis of the subfossil Ostracoda in all the studied lake ecosystems occurred in the phase of settlement related to the Early Middle Ages, which was reflected in the permanently changed species composition of the fauna.
Introduction
Lake sediments are a natural archive of past environmental changes. Historically, research has focussed on generating regional climate registers, but registers of human impacts caused by land use and freshwater exploitation are now attracting scientific (Cohen, 2003; Dubois et al., 2018). Palaeolimnology (studies on the history and evolution of lakes) supplies information on environmental change for periods prior to instrumental observations. Long-term records are helpful in determining the extent of natural ecosystem variability in the absence of extensive human influence. The timing and magnitude of anthropogenic disturbance can be figured out by reference to the state of the ecosystem at a time when its functioning was not subject to anthropopression (Armesto et al., 2010; Barr et al., 2013; Dubois et al., 2018). In the works describing the results of palaeolimnological studies, it is emphasised that the human impact on aquatic ecosystems registered in lake sediments is very variable in time and space. According to archaeological studies, although freshwater ecosystems occupy only approximately 1% of the Earth’s surface, they have had an enormous impact on the development and functioning of human cultures, and therefore have been subject to strong human pressure in the past (Saunders et al., 2002). Anthropogenic deforestation in the lake’s catchment area causes an increased inflow of allochthonous matter (soil erosion, nutrient enrichment), leading to a decrease in water transparency, an increase for organic matter in sediments and changes in the availability of nutrients affecting the biota of the lake (Cohen, 2003). In the case of lakes with a small area of a few ha and the maximum depth not exceeding 3–8 m, changes in living conditions for aquatic organisms may occur relatively quickly. Shallow lakes are known to have proportionally higher loadings and lower losses of nutrients than deep lakes (Tammelin and Kauppila, 2018; Wetzel, 2001). Deeper naturally eutrophic lakes seem to have been better able to keep their pre-disturbance conditions under anthropogenic forcing.
The response of individual species that make up the lake’s biota is different. This reaction in the form of the occurrence or disappearance of species occurs most quickly in the case of organisms characterised by high migratory ability and short life spans. One of many groups of organisms used as ecological indicators are Ostracoda. They are small (0.2 mm to approximately 3 mm) crustaceans occurring in modern times as well as the geological past in diverse water environments, often in lakes (Holmes, 2001; Meisch, 2000; Sywula, 1974). Most of the species exclusively inhabit sediments (benthic species), some of them are active swimmers, and some constitute a part of the pleuston (Meisch, 2000; Sywula, 1974).
Research on Ostracoda permits increasingly precise identification of the range of ecological tolerance of the species in reference to most environmental factors. This allows for the designation of the most characteristic species or communities whose presence or abundance fluctuations can be treated as for example, an indicator of changes in temperature, salinity, and indirectly water depth, dissolved oxygen concentration, type of sediment and sedimentation rate, or water dynamics (Smol et al., 2001; Viehberg, 2006). This is exemplified by study results of modern fauna communities and their seasonal changes in freshwater ecosystems staying under the influence of intensive agricultural activity. They suggest that some species are resistant to a particular type of organic pollutants, for example, to high content of nitrate nitrogen (Lim and Wong, 1986; Rossetti et al., 2004). Lack of live individuals was recorded in the case of a considerable increase in content of phosphates in the surface sediments of some lakes (Rossetti et al., 2004; Wünnemann et al., 2006). More detailed research showed that phosphates cause considerable disturbances in certain species of Herpetocypris, nitrates strongly limit the development of Candona neglecta (Milhau et al., 1997), and Ilyocypris inermis shows no occurrence in ecosystems with an elevated level of biogenic pollutants (Pieri et al., 2012). Ecosystems of rivers and lakes strongly polluted with municipal or industrial wastewaters often show complete disappearance of Ostracoda assemblages, or considerable drop of both their species composition and abundance (Poquet et al., 2008). High intensity of human pressure limits and changes the conditions of functioning of Ostracoda fauna assemblages in inland water ecosystems. It therefore seems justified to try to use these organisms as indicators of human pressure from the period for which archaeological data show a significant impact of anthropopression on the natural environment (the process of neolithization). In the study area the first Neolithic cultures developed in areas dominated by forest vegetation communities (deciduous forests). The greatest transformation of forest cover into arable fields occurred in the periods of development of the Funnel Beaker culture, when new cultivation techniques were introduced, and in the early Middle Ages, when the spatial extent of arable fields expanded considerably.
This paper introduces a summary of existing archaeological knowledge and paleoenvironmental research within the study area and is enriched by original data on Ostracoda analysis to estimate: (1) to what extent has the pressure of settlement of subsequent cultures influenced the landscape and to what extent has it disturbed the functioning of lake ecosystems and (2) whether the neolithization as the first clearly legible stage of anthropopression had a decisive influence on the functioning of lake ecosystems.
Determining the timing and intensity of anthropogenic changes in lake ecosystems helps in scientific understanding of the complex interactions between humans, climate and the environment. Knowing the time of first impact supplies data on the response and/or regeneration of the water system for periods predating instrumental observation, and an insight into the legacy and cumulative effects of early human impacts on contemporary water systems. This article is an example of a feasible way to expand the ability to accurately (due to the sensitivity and rapidity of reaction of the Ostracoda fauna described) determine the extent of anthropopression on an aquatic ecosystem when instrumental data cannot be obtained.
Study area and lakes
The lakes under study are situated in Volhynian Polesie, where although there are no clear landscape boundaries, the structural differences, and the set of physical-geographic features (hydrographic, climatic and biogeographic features) allow to define the boundaries of Western and Eastern Europe. At the same time, the area is the border between the Central European Lowlands (Western Polesie in the north) and the Polish Uplands (the Lublin Upland and the Volhynian Upland in the south). The lakes are in the northern part of the Chełm Hills mesoregion, belonging to Volhynian Polesie (Kondracki, 2002) (Figure 1). Chełm Hills with maximum absolute height reaching 290 m a.s.l., do not constitute a compact uphill landform (Harasimiuk et al., 2008), but elevations of the Cretaceous bedrock are separated by deep troughs.

Location of the analysed lakes (I) and cross-sections (II) of the area. The location of the lakes under study (I), the course of the cross-section lines (II) and cross-sections with plans of the modern bathymetry of the reservoirs (III).
The examined lakes are some of the few water reservoirs in Volhynian Polesie. The Lublin Upland, found south of these areas, is an area devoid of natural lakes (not including oxbow lakes). In turn, the specificity of the natural environment of the areas lying to the north, that is, numerous peat bogs and lakes, was a barrier to prehistoric and early mediaeval settlement as it is emphasised in archaeological studies (Libera and Taras, 2011). Settlement processes were concentrated here only on the outskirts of the Lublin and Volhynian Uplands, and in the middle, longitudinal belt of Western Polesie (Bałaga, 1991). The morphometric features of selected lakes, their easy availability, enabling them to be used as a water source and in economic activities (such as hemp retting or fishing), make them a good example of the impact of the economic activity of settlement cultures on the functioning of the water ecosystem.
Lake Słone (N: 51°18′16,88″ E: 23°21′55,89″, surface area – 3.4 ha; maximum depth – 8.1 m) is found at an absolute height of 185.6 m a.s.l. The lake catchment is found between the elevations of the Upper Cretaceous occurring to the north and south, covered with Quaternary deposits. The lake basin is surrounded by a peat plain. To the south and south-west, the depression occupied by the lake-peatland complex is confined to the elevated areas of moraine plains. To the north and north-west, the peat plain is next to an outwash plain and moraine plain. The eastern part of the lake catchment covers the area of a denudational plain.
Lake Syczyńskie (N: 51°14′46.97″, E: 23°20′33.19″, surface area – 5.7 ha, maximum depth – 3.9 m) is found at an absolute height of 179.6 m a.s.l. The extensive eastern part of the catchment covers elevated areas of Sub-Quaternary bedrock, featuring moraine plains. Long slopes of the moraine plateau extend towards the west, transitioning into surfaces of lacustrine plains, and further into the surfaces of a narrow belt of a peat plain, surrounding the lake only over a small area. The western boundary of the catchment runs along the elevation of a kame hill directly next to the peat plain.
Lake Pniówno (N: 51°14′46.97″, E: 23°20′33.19″, surface area – 3.94 ha, maximum depth – 2.8 m) is found at a hight of 179.6 m a.s.l. between two Cretaceous hills. It directly surrounds the narrow area of the peat plain to the north and west, next to the areas of the lacustrine plain. The southern part of the catchment is occupied by a moraine plateau, transitioning into a moraine plain in the south-eastern fragment of the catchment.
The analysed lakes differ in their hypsometric position. They also have different bathymetric features (Figure 1), and currently different trophic status of waters: from mesotrophic Lake Słone to periodically hypereutrophic Lake Syczyńskie (with build-up areas located very close to the shoreline) (Pęczuła et al. 2014).
Materials and methods
The analysed lake sediment cores were sampled by means of a set by Eijkelkamp for manual coring, with the application of an ‘Instorf’ sampler (length 0.5 m; diameter 5 cm) from the ice cover in the central, currently deepest parts of the lakes. The sampled cores covered entire profiles of biogenic sediments, subject to palynological analyses, analyses of percent share of organic matter and calcium carbonate, and radiocarbon datings. The analyses of changes occurring in the water ecosystems of selected lakes presented in this paper are based on changes in total abundance as well as presence and disappearance of specific Ostracoda species in subfossil assemblages. The analysis of fossil Ostracoda valves and carapaces covered sediment samples collected from each of the 5 cm sections of the cores, with a volume of 5 cm3, so that each sample contained a 5 cm section of depth. The collected samples were left in distilled water until complete dampening. The dampened sediment samples were rinsed on analytical sieves with a mesh diameter of 125 μm, and the residue dried at room temperature was subject to analysis under a stereoscopic microscope involving identification of fossil Ostracoda. The obtained material was then classified in terms of species affiliation based on features of their physical structure. The material for species identification was rinsed under the microscope with distilled water. After species identification of shells, their abundance was determined by species. The results are presented in the form of diagrams created through the PANGAEA/PanPlot programme (Diepenbroek et al., 2013). The interpretation of the obtained results are based on information on the ecology of Ostracoda species, which was collected from scientific literature generally characterising the ecology of Ostracoda and their importance in paleoenvironmental analyses (Danielopol et al., 1985; Delorme, 1991; Frenzel et al., 2010; Fuhrmann, 2012; Geiger, 1993; Griffiths and Holmes, 2000; Kiss, 2007; Külköylüoğlu, 2004, 2005; Külköylüoglu and Vinjard, 2000; Löffler, 1969, 1975; Meisch, 2000; Roca and Danielopol, 1991; Scharf, 1980; Sohar, 2010; Sohar and Kalm, 2008; Sywula, 1974; Wetterich et al., 2005).
The archaeological context is presented based on published archaeological data describing sites of archaeological research, and studies synthesising current knowledge about the development of settlement processes in the Chełm Hills (Bienia, 2003; Dąbrowska, 1988; Dulinicz, 2001; Godłowski, 1984; Kadrow and Zakościelna, 2000; Libera, 2006; Libera and Taras, 2011; Nogaj-Chachaj, 2019; Parczewski, 1988, 2005) and neighbouring areas (the lowland part of Polesie in the north and the Lublin Upland in the south) (Taras, 2005; Taras and Bałaga, 2012; Zakościelna, 1996).
The overview of the paleoenvironmental context is based on study results published earlier as paleoenvironmental reconstructions covering the entire period of functioning of lakes – from the end of the last glaciation (Kulesza, 2017; Kulesza et al., 2011, 2012; Kulesza and Bałaga, 2016). The primary task of these reconstructions was to evidence correlations between natural climate changes in the period of the Late Glacial and Holocene and changes in the functioning of lake systems and their catchments. This article uses part of such paleoenvironmental data corresponding to the period from 5000 BC, that is, from the second phase of the Atlantic (Mangerud et al., 1974).
The palynological analyses and designation of successive phases of changes in plant assemblages were performed by Irena Agnieszka Pidek and Krystyna Bałaga following the adopted method (Berglund and Ralska-Jasiewiczowa, 1986; Birks and Berglund, 1979; Janczyk-Kopikowa, 1987).
The analyses of percent contribution of calcium carbonate were performed by volumetric analysis in a Scheibler apparatus. Content of CaCO3 is calculated from volume of emitted carbon dioxide because of reaction of the sediment sample with HCl. The analyses of percent contribution of organic matter were performed through the method of loss on ignition at a temperature of 550°C (Myślińska, 2001).
Radiocarbon dating was conducted in the profile from Lake Pniówno, and from Lake Słone with the 14C AMS method at the Gliwice Radiocarbon Laboratory. Radiocarbon dating of the upper sections of the sediment cores of Lake Syczynskie has not been done due to the lack of selectable material for the AMS dating method.
Results
Changes in Ostracoda fauna
The first designated stage of changes in fauna (at the depth of: 9.65–9.55 m – Słone Lake, 11.85–11.33 m – Syczyńskie Lake and 12.50–12.15 m – Pniówno Lake – Figure 2) at the end of the Atlantic period was reflected in a rapid decrease in the total amount of Ostracoda. It resulted both from a decrease in the abundance of species and decline of a part of species represented in the Atlantic period (Figure 2). The main changes in the species composition involved the disappearance of Darwinula stevensoni, disappearance or a considerable decrease in the share of Fabaeformiscandona protzi and Limnocythere inopinata, and appearance of Cyclocypris laevis. The appearance of Paracandona euplectella, Notodromas monacha and Dolerocypris fasciata is also worth noting.
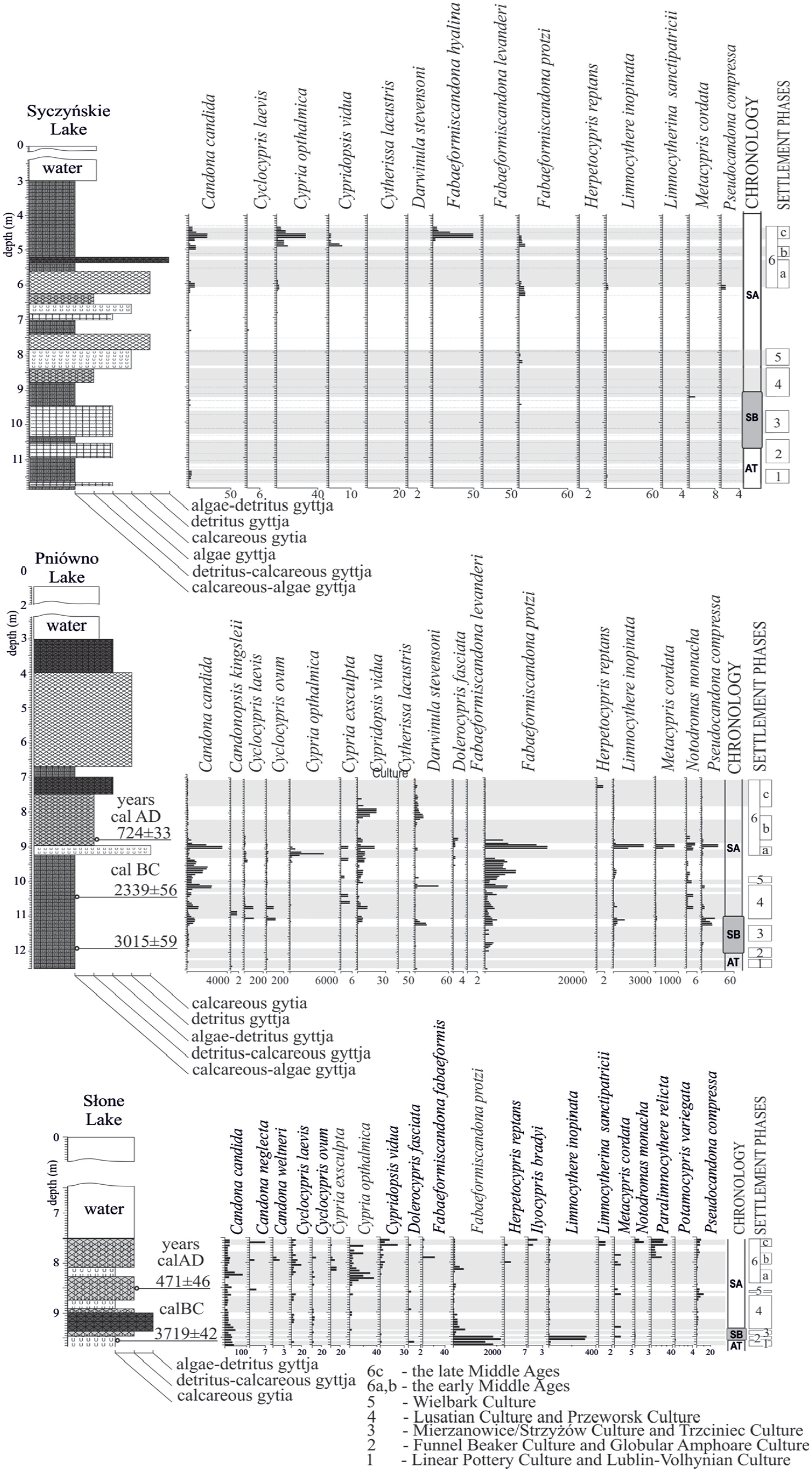
Fragments of diagrams illustrating changes in the frequency and species composition of the Ostracoda fauna in the analysed lake sediments (figure prepared based on previously published diagrams (Kulesza, 2017)).
Over the first half of the Subboreal (the second designated phase of Ostracoda at the depth of: 9.55–9.45 m – Słone Lake, 11.30–10.85 m – Syczyńskie Lake and 12.15–11.60 m – Pniówno Lake), periodical abundance fluctuations with periods of disappearance were primarily observed for Fabaeformiscandona protzi. Periodical changes in the contribution of species Pseudocanadona compressa, Cyclocypris laevis, Cyclocypris ovum and Metacypris cordata are also pronounced (Figure 2).
Towards the end of the Subboreal (the third Ostracoda phase designated at the depth of: 9.45–9.30 m – Lake Słone, 10.85–9.15 m – Syczyńskie Lake and 11.60–11.10 m – Pniówno Lake), regeneration of the species composition of subfossil Ostracoda assemblages is seen. A relatively extended period of functioning of lake ecosystems in stable conditions is suggested by the appearance of a higher number of species periodically absent in the first half of the Subboreal, among others Candonopsis kingsleii, Cyclocypris ovum and laevis, Cypridopsis vidua, Darwinula stevensoni, Limnocythere inopinata, Metacypris cordata and Pseudocandona compressa.
The turn of the Subboreal and first part of the Subatlantic in the record of changes in Ostracoda fauna (the fourthOstracoda phase designated at the depth of: 9.30–9.05 m – Lake Słone, 9.15–8.70 m – Syczyńskie Lake and 11.10–10.65 m – Pniówno Lake) is manifested as a slow process of another decrease in species diversity and total abundance. The clearest changes concerned a gradual decrease in the abundance of Fabaeformiscandona protzi. Other species showed periodical abundance fluctuations, or occurred only in short phases, although it is significant that they formed a fossil assemblage unchanged in terms of conditions. Moreover, Cyclocypris ovum and laevis, Cypridopsis vidua, Darwinula stevensoni, Limnocythere inopinata, Metacypris cordata and Pseudocandona compressa appeared.
In the following phase (the fifth Ostracoda phase designated at the depth of: 9.05–8.45 m – Lake Słone, 8.70–6.40 m – Syczyńskie Lake and 10.65–9.5 m – Pniówno Lake), the species composition structure of Ostracoda showed the appearance of most species occurring previously, although their presence was not constant. The contribution of Fabaeformiscandona protzi, Cypridopsis vidua, Metacypris cordata and Limnocythere inopinata increased, and Dolerocypris fasciata appeared for a brief time. Unfavourable changes in the conditions of functioning of the Ostracoda fauna were supported in the case of Lake Syczyńskie, where only Candona candida and Cyclocypris laevis appeared in small abundance.
Another stage (the sixth Ostracoda phase designated at the depth of: 8.45–8.15 m – Lake Słone, 6.40–5.35 m – Syczyńskie Lake and 9.5–9.05 m – Pniówno Lake) of functioning of lake ecosystems is reflected in the clear response of the Ostracoda fauna involving the depletion of the species composition, primarily through the disappearance of most benthic species. Only Candona candida and Cypria ophtalmica remained, and Cypria exculpta appeared in the subfossil assemblage recorded in Lake Słone.
The seventh Ostracoda phase (designated at the depth of: 8.15–8.00 m – Lake Słone, 5.35–5.15 m – Syczyńskie Lake and 9.05–8.80 m – Pniówno Lake) was marked by significant changes in Ostracoda assemblages. Benthic species appeared again, including Fabaeformiscandona protzi. This short phase of an increase in species diversity and abundance of Ostracoda appeared in all the analysed lakes, as suggested by changes in the functioning of lake ecosystems of supralocal character.
In the eighth phase (designated at the depth of: 8.00–7.85 m – Lake Słone, 5.15–4.90 m – Syczyńskie Lake and 8.80–8.20 m – Pniówno Lake) of changes in subfossil Ostracoda fauna again involved a decrease in the abundance, or even disappearance of the fauna, like in the case of Lake Syczyńskie and Pniówno. Only in Lake Słone, Cypria ophtalmica and Cypria exculpta remained.
The distinguishing feature of the next – ninth phase of changes (designated at the depth of: 7.85–7.65 m – Lake Słone, 4.90–4.60 m – Syczyńskie Lake and 8.20–7.80 m – Pniówno Lake) was regeneration of aquatic fauna. Among Ostracoda, some of the species previously occurring appeared, such as Candona candida, Cypridopsis vidua, Fabaeformscandona protzi, Darwinula stevensoni and Limnocythere inopinata. Next to these species, species not registered before were found, such as Paralimnocythere relicta, Herpetocypris reptans, Fabaeformiscandona fabaeformis, Candona weltneri and Candona neglecta. Such a large share of new species in subfossil assemblages marks a new stage in the functioning of the ecosystem.
The last – tenth phase of changes (designated at the depth of: 7.65–7.50 m – Lake Słone, 60–4.00 m – Syczyńskie Lake and 7.8–7.50 m – Pniówno Lake) was reflected in the disappearance of the Ostracoda fauna. Complete disappearance of fauna was characteristic of Lakes Pniówno and Syczyńskie. In Lake Słone, species Candona candida, Cypria ophtalmica and Paralimnocythere relicta were periodically present, and for a brief period also Pseudocandona compressa and Candona neglecta.
Archaeological context
In the majority of paleoenvironmental analyses, a considerable impact on the functioning of lake ecosystems is manifested in the period of sustained land-use changes at local scale, caused by the agriculture of the Neolithic period (Ruddiman, 2005; Whittle and Bickle, 2014). Regions in various parts of Europe, also in contemporary Poland, have their own history of human disturbances:
(1) In Volhynian Polesie, the beginning of the Neolithic is marked with scarce traces of settlement of the Linear Pottery culture. Settlement of an agricultural population inflowing from the south (Danubian circle) in several subsequent waves beginning from about 5500 BC (Bienia, 2003; Libera, 2006; Libera and Taras, 2011) was primarily focussed on loess uplands 50–70 km south of the Chełm Hills. In older stages, it only slightly encroached on the Lowland, and therefore on Polesie, as suggested by scarce archaeological sites with single stone tools characteristic of the Linear Pottery culture. The archaeological traces of more intensive human activity in Polesie refer to the settlement of the Lublin-Volhynia culture (Kadrow and Zakościelna, 2000; Zakościelna, 1996).
(2) Intensified Neolithisation of Polesie occurred together with the appearance, according to archaeological data at the beginning of 3600 BC, of the Funnel Beaker culture (Nogaj-Chachaj, 2019). It is showed by abundant archaeological sites also north of the Chełm Hills (Taras, 2005). Moreover, it is a Neolithic culture best represented in terms of number of sites in Polesie (Bienia, 2003). Ca. 3500 BC, in the Central Polish Lowland area, the Globular Amphorae culture developed. It also spread to the areas of Polesie, although sites of that culture are scarce there.
(3) Ca. 2200 BC (Early Bronze Age), the Mierzanowice culture and after Strzyżów culture started appearing in Volhynian Polesie. The proper beginning of the Bronze Age within this area is marked by the emergence of the Trzciniec culture, around 1900 BC. The Trzciniec culture settlements in Polesie are poorly recognised. According to 14C dating, the structures were created between 1900 and 1600 BC. Colonisation of Polesie by the communities of the Trzciniec culture, in consequence caused a quite rapid assimilation of several separate local Neolithic traditions, and eventually cultural unification (Libera and Taras, 2011; Taras and Bałaga, 2012).
(4) At the end of 1200 BC, the discussed areas were inhabited by the population of the Lusatian culture. Its development showed evolutional course, and its importance according to Kłosińska (2007), increased in the earlier part of the age (from ca. 1300 BC to ca. 1200 BC), although it still existed at the beginning of the Iron Age. Co-existence in this area and gradual replacement of the Lusatian culture by the Pomeranian culture spreading from the north in the earlier period of the Iron Age was a push for the development of the Przeworsk culture (Dąbrowska, 1988). Archaeological sites suggest that it appeared in the analysed area in the period of ca. 550–200 BC.
(5) Ca. 200 AD, the Wielbark culture reached the study area (influence of Goths). In the first half of the 5th century AD, settlement became scarcer, and disappeared completely, which in archaeological studies is associated with the Migration Period started by the attacks of Huns in 375 (Godłowski, 1984). Communities inhabiting the vicinity of the analysed lakes mostly abandoned the area, as also confirmed by pollen diagrams (a clear decrease in the proportion of plants indicative of human activity and increase in the share of tree pollen in pollen diagrams from the sites Durne Bagno, Moszne and Krowie Bagno located in the Polesie adjacent to the study area – Bałaga and Taras, 2014).
(6) The beginning of the Middle Ages is marked by archaeological findings related to the beginning of the Slavic culture. High intensity of settlement processes is expressed in a considerable number of archaeological sites from the period throughout the area of the Chełm Hills, but also presence of settlements, and even fortified settlements in the vicinity of the analysed lakes, showing the permanence of the settlement processes. In 1031, after the period of Polish-Ruthenian battles, the areas between the Wieprz and Bug Rivers, including the study area, were eventually found under the rule of Ruthenian princes. Another considerable development of settlement was recorded in the 12–13th century. It was probably related to the agrarian revolution. The younger Mediaeval settlement phase in archaeological research is marked by thorough socio-cultural changes caused by urbanisation and development of new technologies (e.g. pottery) (Dulinicz, 2001; Parczewski, 1988, 2005). Rapid limitation of settlement processes occurred because of Tatar raids in the mid-13th century. Another limiting factor were raids by the Yotvingians and Ruthenians, marking the end of the Early Middle Ages.
In the 14th century, eastern Slavs dominated the area. Development of settlement was inhibited by long-lasting Polish-Lithuanian-Hungarian fights and Tatar raids.
Paleoenvironmental context in the light of palynological data
The concept of anthropogenic indicators in pollen record has been presented in numerous studies, which supply the characteristics of diverse groups of indicators serving in the assessment of the human activity (Berglund and Ralska-Jasiewiczowa, 1986; Faergi and Iversen, 1989; Latałowa, 2003, 2007). When analysing the results of palaeoenvironmental studies conducted for the sediments of the studied lakes (Kulesza, 2017; Kulesza et al., 2012; Kulesza and Bałaga, 2016), a clear change in pollen spectra at the turn of the palynostratigraphically designated Atlantic and Subboreal periods can be noticed. The results of radiocarbon dating of the sediments of Pniwno and Słone Lakes showed an age of about 3700 BC. In the case of Lake Syczyńskie in the sediments of which no material for AMS dating was obtained, the identification of the chronology of paleoenvironmental changes was based on palinostratigraphic correlation with dated profiles. (Kulesza, 2017). The boundary between the Atlantic and Subboreal (Figure 3) is determined by a decrease in the percentage of values of Ulmus. Percent curves of Quercus, Corylus and Alnus show considerable fluctuations of values. Characteristic is the presence of the grazing indicator in this horizon, namely Plantago lanceolata (Latałowa, 2003).

Fragments of percentage pollen diagrams illustrating the variability of the share of taxa of trees and shrubs and indicator plants of anthropogenic activity found according to: Berglund and Ralska-Jasiewiczowa (1986), Latałowa (2003, 2007). Figure compiled from previously published palynological diagrams (Kulesza, 2017) created in POLPAL (Nalepka and Walanus, 2003).
The curve of Carpinus and Fagus appears, with an increasing share in the central part of the Subboreal, and decreasing in its younger part, like the values of Corylus and Fraxinus. The decrease correlates with the simultaneous increase in the curve of Quercus, Salix, and Populus. The recorded paleoenvironmental changes during the beginning of the Subboreal period in the analysed objects, however, also covers an increase in the share of nitrophilous vegetation (Urtica, Chenopodiaceae, Artemisia).
At the beginning of the Subatlantic a characteristic element of paleoenvironmental changes was an increase in the share of hornbeam, and a decreasing share of lime and elm in forest assemblages. The development of oak-hornbeam forests resulted in a significant reduction in the share of hazel in plant communities. The presence of nitrophilic plants and grazing indicators was also characteristic for this period. The next stage was the phase of secondary succession of forest communities clearly marked in the pollen spectrum. According to the radiocarbon result for sediments from Lake Słone, it occurred 471 ± 46 cal. years AD. In Lake Pniówno, the age of sediments from the final phase of forest regeneration was 724 ± 33 cal. years AD (Figure 3).
The next stage was reflected in palynological diagrams as a bipartite period of high contribution of NAP, large expansion nitrophilic plants and grazing indicators, and a considerable decrease in the share of most trees developing forest assemblages, except for pine. The key feature of environmental changes recorded in palynological diagrams of that stage of transformation was presence of pollen of cereals and more abundant grazing indicators (Behre, 1981; Latałowa, 2003; Lityńska-Zając and Wasylikowa, 2005). Characteristic was intensive development of nympheids reflected in the palynological spectrum and the remarkably high contribution of Cannabis pollen in the pollen spectrum. Next changes are demarcated by a slight increase in the share of the main taxa of trees in the palynological diagram. A considerable decrease in the contribution of Cannabis pollen is also characteristic (clear in all the analysed objects). The next stage in palynological diagrams is marked by continuous curves of percent values of pollen of rye (Secale cereale) and other cereals (Cerealia undiff.) with a simultaneous decrease in the share of hornbeam and hazel.
The contribution of pollen of plants of ruderal habitats simultaneously increased, including nitrophilous nettle (Urtica). The grazing indicator, namely pollen of ribwort plantain, occurred regularly in palynological diagrams accompanied by taxa of herbaceous plants, characteristic of humid meadows (Filipendula, Apiaceae). Another increase in the share of Cannabis sativa pollen was marked.
At the next stage of paleoenvironmental changes, values recorded in palynological diagrams suggest intensive development of pioneer assemblages of birch-pine forests, but also the development of oak-hornbeam assemblages. It is reflected in a considerable increase in percent values of hornbeam, oak and hazel, and formation of a continuous percent curve of lime and ash.
The last stage of environmental changes recoded was characterised by rapid decreases in percent curves of the main trees, and in an increase in numerous herbaceous species, including cereals, nitrophilous, and grazing plans (Figure 3). Cannabis was again present in palynological diagrams, characteristic was also disappearance of water lilies and considerable limiting of the rush reflected in the palynological diagram in lower values of pollen frequency curves.
Interpretation and discussion
Large and deep lakes tend to have high buffering ability. This means that they tend to be lagging the nutrient input from the catchment area. Smaller lakes may show little lag between the change in land use and the lake response (Dearing and Jones, 2003). At the same time, small lakes are more sensitive to climate change, which makes it difficult to clearly interpret the anthropogenic impact. The conducted research shows that climate changes recorded in lake sediments can be correlated with the phases of deep changes in settlement cultures (periods of migration or influx of new settlement cultures) (Figure 4).
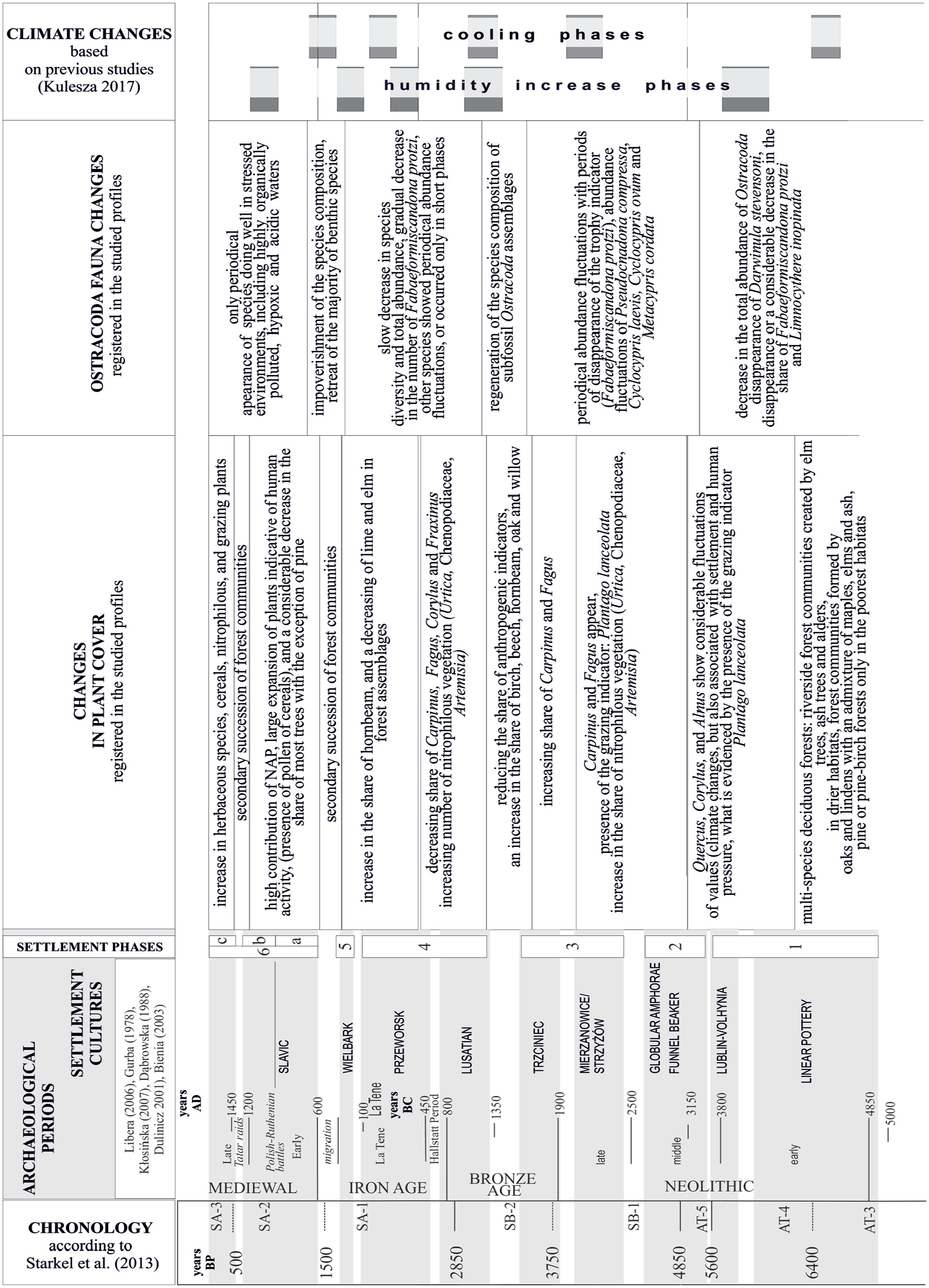
Chronology of anthropogenic and climate changes recorded in sediments of studied lakes Słone, Pniówno and Syczyńskie in Chełm Hills region.
Periods of cooling down and lowering water levels in lakes can highly likely be related to changes in plant cover, which are not accompanied by the appearance of indicators of anthropogenic economic activity (as highlighted in Figure 3). Examples of cooling phases showed in this way are related to the periodic increase in the share of sedges, among them Cladium mariscus. Simultaneous disappearance of Pediastrum algae (decrease of water depth) and an increase in the share of Equisetum spores and Sphagnum shows the development of wetlands around lakes with communities composed of sedges with paniculate logs, sphagnum mosses and horsetails (e.g. in the profile from Lake Słone at depths of 8.2 and 9.5 m, Lake Syczyńskie at depths of 9.3, 10.5 and 11.60 m, and in the sediment profile from Pniówno Lake at depths of 11.30 and 10.8 m).
In turn, the functioning of lake ecosystems changed primarily in the phases of full development of next cultures. However, the intensity of these changes increased gradually, even though the selected objects belong to the group susceptible to anthroporession due to their morphological features.
According to archaeological sources, the beginnings of Neolithic settlement in the areas of Volhynian Polesie date back to the first phase of the spread of Linear Pottery culture (one settlement phase; Figure 4) from the middle Danube (Bohemia, Moravia, Hungary) northward across Carpathians around 5500 BC (Kozłowski et al., 2014). The territorial distribution of archaeological finds shows that the largest concentrations of settlements of this culture were in the upland loess areas, mainly in the western Volhynian Upland. Further north, in Volhynian Polesie, the sites of this culture are very scattered, usually occurring individually and at considerable distances from each other. It may show that they are probably a remnant of occasional economic expeditions to areas outside the main settlement ecumene of the Linear Pottery culture. The scattered nature of the settlement and the occasional use of environmental resources were not reflected in the changes of Ostracoda. The area around the lakes was occupied by multi-species deciduous forests, and the slight decline in species diversity and the periodic decline of some Ostracoda species (Darwinula stevensoni and Limnocythere inopinata) can be attributed to climate change in the latter part of the Atlantic period for around 4000 years BC.
Considering the changes in the functioning conditions of lake ecosystems, the second stage of the neolithization of Volhynian Polesie, that is, the development of the Lublin-Volhynian Culture (end of one phase of settlement, Figure 4), can also be included in the first phase of settlement development. According to archaeological data, the economic model of the Lublin-Volhynian was based on small, compact areas in the vicinity of settlements (Reder et al., 2010). The intensification of anthropopression was characteristic mainly for upland areas, like the development of Linear Pottery culture. The changes found (decrease in Fabaeformiscandona protzi, Limnocythere inopinata and Candona candida) can indicate anthropogenic impact on the ecosystem (Table 1), but the impact was not strong enough to permanently alter ecosystem conditions.
Primary data characterising ecological conditions affecting the development and possibilities of survival of species and sources of information.

The next phase (two phase of settlement, Figure 4) is found as a more intensified Neolithisation of the area in question. Funnel Beaker culture formation dated according to archaeological data around 3100 BC introduced of new agrotechnical methods such as large-area grass burning and horse ploughing (Gurba, 1978). The development of settlement occurred during the period of climate change at the turn of the Atlantic and Subboreal periods. Despite the increase in the intensity of settlement and the enlargement of cultivated areas, a pattern of changes in the Ostracoda fauna was like the previous settlement phases. Only the periods of decline in the share of Fabaeformiscandona protzi, Limnocythere inopinata and Candona candida were longer. The increase in the share of nitrophilic vegetation (Urtica, Chenopodiaceae, Artemisia), characteristic for this period, is interpreted as an expression of anthropogenic pressure (Behre, 1981; Latałowa, 2003). Simultaneous recording of longer periods of decline of Fabaeformiscandona protzi can be interpreted as an increase in the fertility of lake waters caused by the increased inflow of allochthonous matter (soil erosion, nutrient enrichment) (Table 1).
After the period of functioning of communities with mixed traditions of Subneolithic and Neolithic cultures, late Globular Amphora culture, and Corded Ware culture settlement, changes related to the inflow of the Mierzanowice and then Strzyżów culture about 2500 years BC could have been related to limiting economic activity (in palynological diagrams expressed in a decrease in the percent share of anthropogenic indicators). This is suggested by an increase in the abundance of Fabaeformiscandona protzi.
In the third settlement phase (Figure 4) (settlement of the Mierzanowice, Strzyżów since about 2500 years BC, and Trzciniec cultures), Fabaeformiscandona protzi declined in lakes once again, and total abundance of Ostracoda considerably decreased.
The fourth phase of settlement (Figure 4) according to archaeological data primarily related to the development of the Lusatian culture about 1300–800 years BC and Przeworsk culture up to about 100 years AD involved a similar response of the Ostracoda fauna. Changes in palynological diagrams indicate that human economy, eliminating species of fertile soils (lime, elm), contributed to the immigration, and spread of hornbeam (Ralska-Jasiewiczowa et al., 2003). The economic activity had a breeding-agricultural character, with a dominant position of animal breeding, and considerable share of assimilation economy (Czopek, 1997; Taras, 1995). Despite a small share of farming, this phase was also recorded as unfavourable for the functioning of the Ostracoda fauna. This is indicated by much longer periods of disappearance of species and larger periodic fluctuations in abundance. However, the species composition itself remained without significant changes (disappearance and reappearance of the same species). Stronger anthropogenic impact can be also associated with a direct impact on lake waters by hemp retting (Milecka, 1998), which is reflected in palynological diagrams.
Between about 800 and 450 years BC, the development of benthic species shows stable oxygen conditions in the zone of contact between water and lake sediments. In all analysed lakes, fauna development was accompanied by an increase in the share of organic matter in sediments. The phase of regeneration of the vegetation (forest) covers also occurred, with a considerable increase in the share of hornbeam, lime, elm, oak, and hazel (at the end of Subboreal period).
From about 450 BC another gradual decrease in the abundance of Fabaeformiscandona protzi, which can be treated as a manifestation of a permanent trend involving an increase in water fertility (second part of the fourth phase of settlement; Figure 4). Other species showed periodical fluctuations of abundance, or only occurred in short phases. This is likely caused by the nature of settlement changes occurring in the area in the beginning of the Iron Age. According to the archaeological data, the development of the Lusatian culture was of evolutional character, while more comprehensively developed economic activity of the culture’s population was only characteristic in the earlier part of the Bronze Age and survived until the beginning of the Iron Age. The Lusatian culture was also gradually replaced by the Pomeranian culture expanding from the north in the earlier period of the Iron Age. These evolutional changes can be reflected in the multidirectional character of changes in the functioning of lake ecosystems themselves. A gradual increase in fertility of lake waters followed by a periodical decrease in human pressure resulting in a reduction of trophic status and relative stabilisation of oxygen conditions could have favoured the appearance of subfossil fauna assemblages of Ostracoda in various configurations of species at later stages of changes occurring in this settlement phase. A characteristic element of the human economy was hemp retting in lake waters (again high percent share curves of Cannabis pollen in the palynological diagram). This direct interference in the functioning of lake ecosystems was certainly also a factor changing the functioning of water fauna, although the intensity of this factor was ambiguously reflected in subfossil Ostracoda assemblages as a new element of human pressure. The observed changes in the abundance and species composition only allow to conclude on periodical increases and decreases in water trophic status (most likely related to the fifth phase of settlement in which the development of the Wielbark culture took place; Figure 4).
In the phase corresponding with a clear break in settlement processes recorded in palynological diagrams and correlated with the Migration Period started by the attacks of Huns in 375 (Godłowski, 1984), the structure of the species composition of Ostracoda includes most of the species previously occurring, although their presence was not constant. The contribution of Fabaeformiscandona protzi, Cypridopsis vidua, Metacypris cordata and Limnocythere inopinata increased. Dolerocypris fasciata appeared for a brief time. Unfavourable changes in the conditions of functioning of Ostracoda fauna occurred in the case of Lake Syczyńskie, where only Candona candida and Cyclocypris laevis occurred in scarce abundance. In sediments of all the lakes, content of organic matter decreased during that time. Combined with new development of benthic species, it may be evidence of a decrease in the trophic status of lake waters.
The most significant changes in the previously described phases of human pressure intensification found in the Ostracoda analyses primarily cover the disappearance of Fabaeformiscandona protzi. It is a species declining in habitats with higher-than-average trophic status (Table 1) (Danielopol et al., 1985). A large majority of Ostracoda, such as Fabaeformiscandona protzi, are benthic forms (Table 1) inhabiting the surface of bottom sediments, incapable of free swimming. The occurrence of anoxic conditions resulting from increased trophic status usually first covers the zone of contact of lake waters and bottom sediments. In such conditions, limiting the availability of dissolved oxygen directly affects benthic species, and it is the most frequent cause of a clear decrease in species diversity of Ostracoda assemblages (Rieradevall and Roca, 1995). It is essential that the estimated time of response of Ostracoda to low oxygen content is less than 1 month (Martín-Rubio et al., 2005). Simultaneously with the disappearance of Fabaeformiscandona protzi, and later other benthic species such as Darwinula stevensoni, Limnocythere inopinata, Pseudocnadona compressa and Metacypris cordata, the appearance of Cypria ophtalmica was recorded. It was determined to be one of the scarce species able to tolerate low oxygen concentration over short periods (Table 1). This suggests periodical occurrence of anoxic conditions in the lakes. The confirmation of these changes is supported by the appearance of Paracandona euplectella which is in turn a species resistant to strong anoxia (Douglas and Healy, 1991). The appearance of Notodromas monacha – a pleuston species (Sywula, 1974), and Dolerocypris fasciata – an actively swimming form (Meisch, 2000) is also very characteristic. Forms capable of free swimming (Table 1) and inhabiting the pleuston could have functioned with no considerable disturbances if periodical oxygen deficits occurred only in the near-bottom layer and did not last long.
A change in the functioning of lake ecosystems at described above phases of settlement was probably concerned with an increase in the trophic status of lake waters, although they were periodical changes and they did not last sufficiently long to cause complete disappearance of Ostracoda assemblages, or permanent transformation of the species composition and lake ecosystems had the ability to adapt rather than shift into an alternative state.
Strong and direct anthropogenic impact on lake ecosystems from the Early Middle Ages (since about 600 AD) can be referred to the clear response of Ostracoda fauna consisting in the decline in the diversity of species composition due to the disappearance of most benthic species. Only species that tolerate periodical deficits of water dissolved oxygen, such as Candona candida or Cypria ophtalmica, remained. The key element of these changes was the persistence of the abundance dominance of species more resistant to organogenic water pollution – Cypria ophtalmica and Cypria exculpta (Table 1) (Meisch, 2000), recorded in Lake Słone. Palynological indicators also show a considerable increase in eutrophication (intensive development of nympheids – Kulesza, 2017).
A brief period of regeneration of forest covers after the settlement phase of Early Middle Ages (phase 6a; Figure 4) recorded in palynological diagrams corresponds with substantial changes in Ostracoda assemblages. Benthic species appeared again, including Fabaeformiscandona protzi. This short phase of increase in species diversity and abundance of Ostracoda occurred in all the analysed lakes. It suggests that the changes in the functioning of lake ecosystems in the form of a decrease in water trophic status and discontinuation of hemp retting can be ascribed to the decline of settlement processes and a decrease in human pressure. This period also shows a high decrease in the share of organic matter in sediments.
In the second stage of settlement development of the Early Middle Ages (since about 1030 years AD, phase 6b; Figure 4), lake ecosystems were again subject to direct strong human pressure. Lake sediments showed not only a substantial increase in the share of organic matter, but also high content of the sand fraction, suggesting intensive denudation processes in lake catchments managed as agricultural fields over a large area. The response of subfossil Ostracoda was another decrease in the abundance, or even disappearance of the fauna, like in the case of Lakes Syczyńskie and Pniówno. Species resistant to organogenic pollution, namely Cypria ophtalmica and Cypria exculpta, only remained in Lake Słone.
The clear phase of regeneration of the tree stands recorded in palynological diagrams (referred to changes in settlement processes archaeologically described for the 11th century) corresponds with the period of regeneration of water fauna. Among Ostracoda, some benthic species occurring earlier appeared again and some species not previously registered were also recorded. The first one was Candona neglecta, characterised as tolerant of periodically occurring (usually occurring in the summer period) low content of water dissolved oxygen (at a level of 0.3 mg/l O2) (Danielopol et al., 1985, Danielopol et al., 1993). Secondly, Herpetocypris reptans was described as tolerant of changes in salinity (mineralisation) of waters in a range from 0.5 to 6.0‰ (Usskilat, 1975), and of an equally broad range of values of water oxygenation (3.8–7.6 mg/l O2) (Külköylüoglu and Vinjard, 2000). Paralimnocythere relicta is a species weakly examined in terms of ecology, although it is known to prefer small water bodies, and it is observed in the shallow littoral of lakes (Meisch, 2000). The analysed period can therefore be described as a new stage of functioning of the ecosystem. The anthropogenic impact appears sufficiently strong to permanently change the species composition of Ostracoda. Whereas in earlier phases the changes primarily involved fluctuations in the abundance of the occurring species, periodical disappearance, and reappearance of the same species after ceasing of human pressure, at the stage of the Early Middle Ages new assemblages of the fauna developed, primarily consisting of species resistant to deficits of dissolved oxygen and tolerant towards the levels of water mineralisation.
The last, 6c phase (Figure 4) of clear intensification of human pressure recorded in the analysed profiles according to archaeological and paleoenvironmental data is the period of the Late Middle Ages with indicators of agriculture and pastoralism. In the palynological diagram, changes also covered intensive deforestation, and the share of sand fraction again increased in the sediments. Record of these changes corresponds with another phase of disappearance of the Ostracoda fauna, suggesting another intensification of anthropogenic pressure. Complete disappearance of the fauna was characteristic of Lakes Pniówno and Syczyńskie. In Lake Słone, species tolerant of high trophic status, organogenic pollution, and low content of dissolved oxygen persisted periodically in low abundance. Differences in the record of changes in the fauna in particular lakes reflect the diversity of the properties of the water bodies. The sensitivity of Lake Syczyńskie to changes in trophic status, high in comparison to the remaining analysed lakes, can be associated with low contribution (only 1%) of the lake’s surface in the total catchment area. This suggests that today as in the past it was particularly prone to supply of biogenic substances from the catchment area. The effect is the occurrence of considerably longer phases of complete disappearance of the Ostracoda in the analysed stages of anthropogenic impact. In the case of Lake Pniówno, with a considerably smaller catchment area, its effect on the chemical composition of lake waters was probably less intensive, as demonstrated by considerably more seldom phases of complete disappearance of Ostracoda, relative continuity of the occurrence of the fauna was characteristic, except for periods identified as phases of strong anthropogenic pressure. Lake Słone can be treated as a lake-peatland complex. The direct vicinity of the lake, constituting peatlands, is currently, and likely was in the past a buffer zone limiting the supply of nutrients from the catchment area. Therefore, the phases of disappearance of the Ostracoda fauna in this lake were the shortest.
The obtained data clearly show substantial limitations in terms of interpretation resulting from the lack of detailed dating of sediments. The studied lakes are shallow, which means that sediments may be subject to mixing. In addition, the lake basins were formed in Upper Cretaceous carbonate rocks. Radiocarbon dating of carbonate deposits are subject to error due to reservoir effects arising when older carbon sources, such as bedrock limestone, result in significant lowering of the initial 14C–12C ratio relative to the global mean atmospheric ratio at the time of formation (Benson, 1993; Broecker and Walton, 1959; Stuiver and Polach, 1977). Due to the limited number of identifiable remains of organic terrestrial plants in the sediments of the Pniówno, Syczyńskie and Słone lakes under study, it was necessary to radiocarbon dating of the mass total organic carbon samples. Bulk samples obtained in this way are prone to incorporate carbon from various sources that are not contemporaneous with sedimentation (terrestrial soils, or reworked sediment), and often yield anomalously old ages (Abbott and Stafford, 1996; Colman et al., 1996). With awareness of these limitations, the coherent palynostratigraphic image corresponding with archaeological facts allows for comparison of changes in subfossil fauna characterising the extent of the effects anthropopression on the functioning of lake ecosystems.
Conclusions
The populations of organisms present in each ecosystem are sensitive to changes in the external environment, and this sensitivity can be written as proportional changes in abundance, evolutionary changes, or extinction. The quick response of the indicators to the forcing variables and the sensitivity of the indicators are crucial in studying the response of the ecosystem to anthropogenic pressure. Ostracoda is characterised by a rapid response to changes in habitat conditions caused by anthropopressure. The reason for this is the brief time required for the emergence of successive generations (from 1 to 2–3 years). Therefore, new faunal assemblages that appear correspond to already changed conditions of temperature, water chemistry and depth or sediment type. The timing and duration of human activity varies around the world and reflects the advances in agriculture, technology, and culture with the advent of humans in different regions. In some areas, the first anthropogenic landscape changes precede (even by several millennia) the first detectable response in water systems, in others (especially in Europe) the initial landscape modifications are synchronised with the first response of water systems. The results presented here indicate that the influence of successive settlement cultures, including the neolithization process, caused ecological changes in the lake catchments until the early Middle Ages, and the lakes themselves were subject to periodic changes in the trophic status of the waters. In all profiles of the studied lakes, the record of periodic disappearance of species sensitive to a decrease in dissolved oxygen (e.g. Fabaeformiscandona protzi), corresponded to periods of appearance of species tolerating periodic depletion of dissolved oxygen and even anoxic conditions in the water (Candona candida or Cypria ophtalmica). A characteristic element of the results of the conducted studies is the record in the form of fossils of the recovery of Ostracoda fauna after the cessation or during the weakening of anthropogenic influence. Examples of these events recorded in palynological diagrams as phases of forest cover regeneration in Ostracoda results corresponded to the restoration of faunal assemblages that existed before the development of anthropopressure. It was not until the second phase of Early Mediaeval settlement that new groups of fauna developed, defining a new state of lake ecosystems – in the case of Lakes Syczyńskie and Pniówno it was another decrease in the abundance, or even disappearance of the fauna, in Lake Słone species resistant to organogenic pollution remained. Even in the conditions of weakening anthropopressure during the Tatar invasions described for the studied area in archaeological sources, there was a development of fauna assemblages in which species new to the studied objects appeared, adapted to the changed conditions. This is evidenced by the presence of species that tolerate increased water mineralisation, periodic anoxia conditions, or a marked decrease in dissolved oxygen (Candona neglecta, Herpetocypris reptans, Paralimnocythere relicta). In addition, the inconsistent pattern of changes in the abundance and species composition of Ostracoda found in the research indicates that a very important factor differentiating the response of lake ecosystems to human impact was the diversity of the lake and the features of its catchment (especially the size of the catchment area in relation to the lake surface and the presence of a buffer zone in the form of wetlands around the shoreline).
The conducted analysis responds to the still valid postulate of more detailed studies of the reactions of lake ecosystems to anthropogenic pressure (e.g. Dubois et al., 2018; Zhang and Mei, 1996). Individual lakes are a record of changes forced both by natural climatic changes and overlapping anthropogenic changes. Based on the results obtained, it cannot be clearly demonstrated what share of natural climate changes had in the recorded changes in lake ecosystems, and to what extent they were the result of anthropopressure. However, a broader application of Ostracoda analyses in reconstructions of human impact on aquatic ecosystems may provide an appropriate data set for more detailed analyzes in the future.