
Abstract
Charcoal shape variations provide insight into past fuel types burned, with charcoal length:width (L:W) being the most popular means of distinguishing fuel types. This paper presents morphometric data (L:W) of charcoal produced from plant taxa (n = 21) native to the southeastern United States of America (USA). These taxa included monocots (n = 3), dicots (n = 12), and gymnosperms (n = 6), which represented a range of growth habits including graminoids, trees, shrubs, and subshrubs. We further divided these taxa into tissues (n = 36 total samples) which included leaves, wood, bark, needles, stems, and a seed pod. Our results show that charcoal produced from plants native to southeastern USA exhibits more variable L:W values than those tested in previous research, but that when aggregated, they broadly agree with published L:W value ranges. Within the dicot group, leaves produced charcoal lower L:W values (median = 1.92, interquartile range (IQR) = 1.33) than wood (median = 2.43, IQR = 3.00), whereas the opposite was true for gymnosperms whose needles produced greater L:W values (median = 2.60, IQR = 3.22) than wood (median = 1.93, IQR = 1.44). The monocot graminoid we tested produced greater L:W values (median = 3.47, IQR = 6.02) than dicot or gymnosperm fuels. Further, our results provide conflicting perspectives on the likelihood of evolutionary relationships as the cause of variations in charcoal L:W values. Last, our results underscore the importance of site-specific experimental charcoal approaches to enable robust paleofire applications of charcoal morphometry.
Introduction
Global fire regimes are changing due to anthropogenic climate change and human land management (Bowman et al., 2009; Flannigan et al., 2009; Moritz et al., 2012). Fire has an integral relationship with vegetation (Bond et al., 2005) and is a key disturbance in many ecosystems (McLauchlan et al., 2020; Thonicke et al., 2001). However, our ability to predict future fire activity is hindered by the complexity of interactions between climate, vegetation, and fire (Bowman et al., 2020; Harris et al., 2016).
Paleofire records provide an informative perspective for the future by characterizing long-term fire relationships in the Earth System (Marlon, 2020). Although many paleofire proxies have been developed (Conedera et al., 2009), sedimentary charcoal is the most commonly used (Power et al., 2010). Incomplete combustion produces charcoal from vegetation which is then dispersed, deposited, and preserved in sedimentary archives (Masiello, 2004; Peters and Higuera, 2007; Snitker, 2018; Vachula and Richter, 2018). Charcoal accumulation has been tied to fire regime characteristics including area burned, severity, and fire frequency (Aleman et al., 2013; Hennebelle et al., 2020; Higuera et al., 2007; Leys et al., 2015; Vachula, 2021; Vachula et al., 2018), and recent work has made considerable headway in characterizing uncertainties and inconsistencies inherent to charcoal reconstructions (Anderson et al., 2023; McMichael et al., 2021; Tsakiridou et al., 2021; Vachula et al., 2022).
Charcoal shape and appearance variations provide a direct means of reconstructing fuel types burned, and allow for the characterization of fire-vegetation relationships (Brown et al., 2023). Early research recognized that different fuel types (e.g. wood vs grasses) produce charcoal particles with differentiable shapes (Umbanhowar and McGrath, 1998). Subsequent work developed morphological keys to distinguish charcoal morphotypes qualitatively and thereby characterize fuel shifts through time from charcoal records (Enache and Cumming, 2006; Frank-DePue et al., 2023; Mustaphi and Pisaric, 2014). Building on earlier work by Umbanhowar and McGrath (1998), quantitative assessments of charcoal morphometric characteristics identified particle aspect ratio, typically referred to as the length:width (L:W) ratio, as a way to distinguish fuel types without the possibility of user-bias affecting fuel interpretations (Crawford and Belcher, 2014).
Experiments producing charcoal from known fuels form the basis of paleoenvironmental interpretations of charcoal L:W data. L:W values of sedimentary charcoal have become a key tool in reconstructing fuel types burned, but the cause of L:W variations between plants remains debated, with some research pointing toward phylogenetically driven plant tissues or biogeochemistry (Umbanhowar and McGrath, 1998; Vachula et al., 2021). Although initial experiments mixed multiple plants to more realistically simulate charcoal production from diverse ecosystems (Umbanhowar and McGrath, 1998; Zhang and Lu, 2006), subsequent experiments combusted taxa individually to resolve potential heterogeneities within plant groups (Ogura, 2007). The roots of paleofire research are in boreal forest systems, and this has methodological implications (Leys et al., 2015, 2018; Rehn et al., 2021; Vachula and Rehn, 2023). Therefore, more recent experiments have sought to characterize fuels native to a wider variety of biomes, biogeographic realms, and climate zones (Feurdean, 2021; Feurdean et al., 2023; Harvey et al., 2023; Pereboom et al., 2020). However, few published experimental studies (n = 7) exist that characterize L:W values of charcoal produced from individual taxa, with the bulk of these studies focusing on the Palearctic biogeographic realm (Table 1). Similarly, few studies have focused on regions with a subtropical climate (Table 1).
Summary of experimental charcoal studies analyzing individual taxa and plant parts.
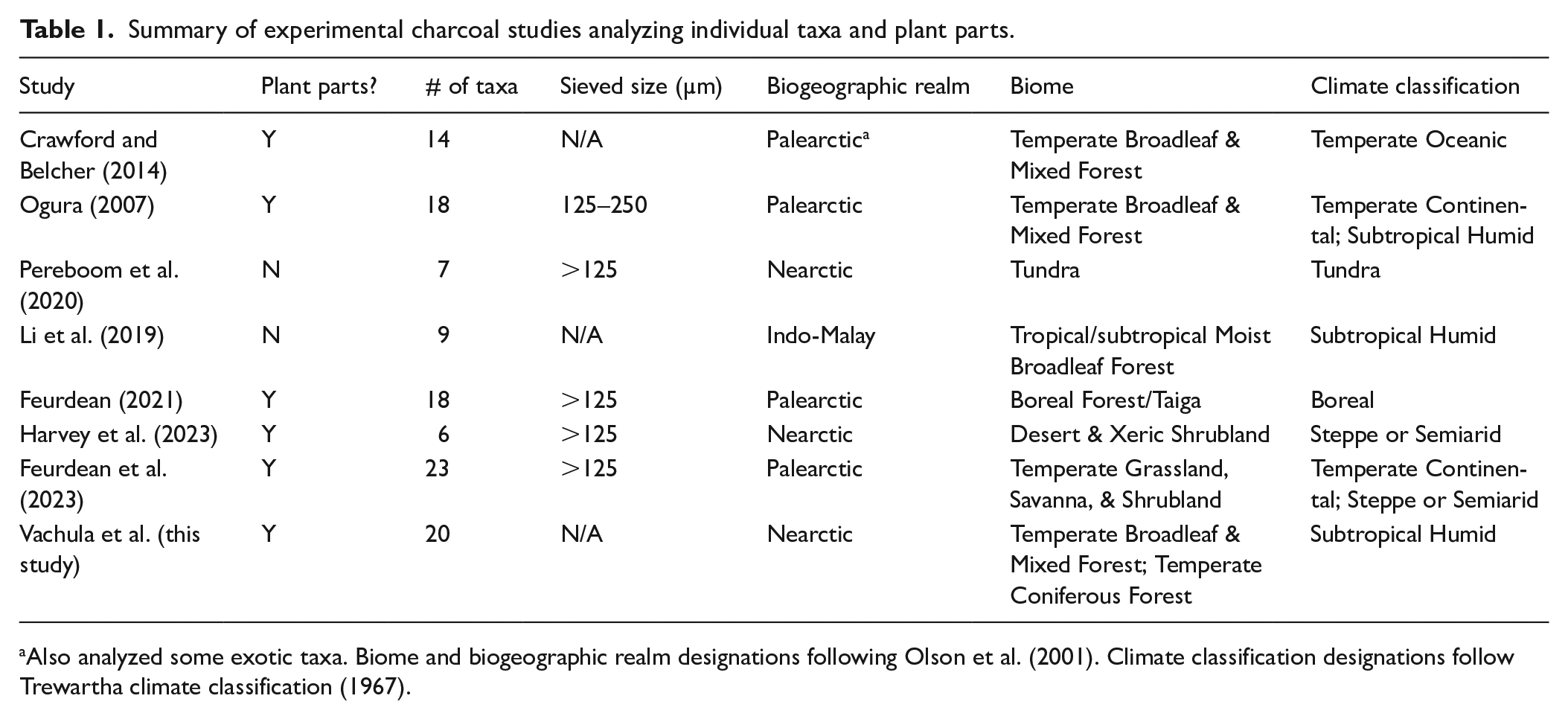
Also analyzed some exotic taxa. Biome and biogeographic realm designations following Olson et al. (2001). Climate classification designations follow Trewartha climate classification (1967).
In this paper, we focus on plants native to the southeastern United States of America (USA), a region with a subtropical climate (Table 1). Understanding fire history and fire-vegetation relationships in this region is particularly important considering ongoing debates regarding the extent of human impacts to Holocene fire regimes in the eastern USA (Oswald et al., 2020; Roos, 2020). Further, prescribed fire is widely applied in this region, beckoning the need to provide long-term fire regime baselines to inform prescribed fire application (Lashley et al., 2014).
To address this gap in experimental charcoal research as well as applied paleofire research in the southeastern USA, we analyzed 21 plant taxa (divided into 36 tissues) native to the region. This dataset comprises the first experimental charcoal dataset from temperate broadleaf and mixed forest, as well as temperate coniferous forest biomes in the Nearctic biogeographic realm. We present these experimental charcoal morphometric data, compare them to previous research from other regions, and discuss their implications for the paleofire field. More directly, this research is guided by the following questions: (1) How do the L:W values of charcoal produced from plant taxa native to the southeastern USA vary and compare to previous findings focusing on other locales? (2) What are the implications of these charcoal L:W values for paleofire applications in the southeastern USA? and (3) What perspective do these data provide for the cause of elongated charcoal and the future of charcoal morphometric analyses?
Materials and methods
Sample collection and preparation
Samples (50–100 g) of 21 plant taxa were collected from leaf litter at two sample locations. First, 15 taxa were collected at the Donald E. Davis Arboretum at Auburn University (32.596 °N, 85.483 °W). The Arboretum hosts a collection of plants native to the state of Alabama and the southeastern USA. Second, 6 taxa were collected from the Graham Creek Nature Preserve in Foley, Alabama (30.345 °N, 87.626 °W). Samples were transported to the lab and stored in open Whirlpak bags for at least 7 days before analysis to air dry. Samples were subdivided into leaf (deciduous), needle (coniferous), wood, bark, and stem tissues. Our use of the term “stem” in our leaf samples refers to the petiole of leaves whereas “wood” refers to the woody tissue of the internode of the plant stem.
The USDA PLANTS database (https://plants.usda.gov/home) was used to assign each plant taxa collected to plant groups (e.g. angiosperm (dicot and monocot), gymnosperm), and growth habits (e.g. graminoid, shrub, tree). Although these plant groups and growth habits are not absolute classifications (in the sense that there is some variability provided by the database), they are consistent with previous research and provide an impartial means of classifying the taxa (Vachula et al., 2021). The PLANTS database differentiates trees from shrubs on the basis of single (trees) versus multiple (shrubs) stems and heights greater than 4 to 5 m. Sub-shrubs are further differentiated from shrubs on the basis of heights <1 m. To compare charcoal morphometry with plant phylogeny, we collected the taxonomic data from the NCBI Taxonomy database (https://www.ncbi.nlm.nih.gov/Taxonomy/CommonTree/wwwcmt.cgi).
In total, we collected samples of 21 plant taxa which were further subdivided by their plant tissues into 36 samples for experimental charcoal production (Table 2). Of these taxa, the majority were angiosperms (n = 15), of which there were 3 monocots and 12 dicots. Six gymnosperm taxa were collected. The majority of plants collected have tree growth habits (n = 12), with lesser numbers exhibiting mixed tree, shrub, and/or sub-shrub growth habits (n = 7), and only one shrub and graminoid growth habit, each. All of the plants collected are perennials.
Experimental charcoal results for plant taxa native to the southeastern United States of America (USA). An average of 50 particles were quantified per sample (tissue).

Groups = Monocot (M), Dicot (D), and Gymnosperm (G)
Tissues = Leaf (L), Wood (W), Bark (B), Needle (N), Stem (S), Seed Pod (SP)
Growth Habit = Graminoid (Gr), Tree (T), Shrub (Sh), Sub-shrub (SSh).
Combustion and sample production
We constructed a combustion apparatus composed of a lidless aluminum can resting atop a petri dish and with its top fitted with a nested wire mesh bowl. We drilled a hole in the side of the can at the height of the base of the wire mesh bowl to allow access of a standard butane stick lighter. Dried samples were placed in the wire mesh bowl and ignited from below with the lighter. Collected charcoal fell into a petri dish at the base of the aluminum can. Samples were reignited to ensure complete combustion of the fuel and that all charred materials fell to the base of the can. Deionized water was used to transfer the resulting charcoal particulates into 15 ml centrifuge tubes for storage. The apparatus was washed thoroughly with water between samples. Samples were gently disaggregated by shaking the centrifuge tubes for about 10 seconds to simulate natural sedimentological breakage (Crawford and Belcher, 2014; Umbanhowar and McGrath, 1998).
Although some previous experimental approaches to characterizing charcoal morphometry have utilized muffle furnaces, charcoal produced in furnaces tends to record higher L:W values (Vachula et al., 2021). Both methods (open burns vs furnaces) likely characterize end-members of oxygen availability (e.g. open burns are aerobic whereas furnaces are anaerobic) during actual biomass burning events. Because previous work has shown that anaerobic combustion in furnaces can inflate L:W values (Vachula et al., 2021), we opted to use open burns to ensure that elevated L:W values reflect fuels and not combustion parameters.
Microscopy and data analysis
Samples were examined and imaged using a Nikon SMZ745T stereo microscope fitted with a DSFi3 color camera. Images were captured and particle aspect ratios (length:width; L:W) were measuring using the CharTool image processing program (Snitker, 2020). For each sample, a minimum of 50 particles was quantified to ensure sufficient characterization of variance of particle L:W (Pereboom et al., 2020).
Sieving of larger particle size fractions can cause inflation of L:W measurements (Crawford and Belcher, 2014), so we did not sieve the samples. To keep track of the equivalent sieved size fraction of charcoal measured, we collected the feret diameter, or the greatest distance been any two points of the edge of the particle, of every particle for which L:W was measured (Snitker, 2020).
We used boxplots to visualize the range of L:W values of charcoal produced from each taxa and plant part. We used two-sample t-tests assuming unequal variances to test the null hypothesis that there was not a significant difference between means of L:W values belonging to groups analyzed in this study. The variances of each group compared via t-test were not equal, thereby necessitating the use of t-tests assuming unequal variances. For statistical comparison of multiple groups, we used single-factor analysis of variance (ANOVA) to test the null hypothesis that the population means of the groups are not different from one another. Next, we used a Tukey’s Honest Significant Test (HSD; using the mean sample size of each group and to the α = 0.05 level) to determine which groups are statistically different from one another.
Results and discussion
Morphometric variations of charcoal produced from plants native to the southeastern United States of America (USA)
Our sampling scheme provides resolution of several plant taxa, growth habits, and tissues for a lesser studied geographic area. Fewer than 100 individual taxa have been subjected to experimental charcoal research (Table 1), and our analyses contribute an additional 20 taxa to this growing dataset. Our analyses also provide charcoal L:W values for monocot taxa that grow as shrubs and trees (Table 2; Figure 1). To our knowledge, all monocots that have been previously subjected to experimental charcoal analysis have graminoid or graminoid/shrub (e.g. bamboo) growth habits (Feurdean, 2021; Feurdean et al., 2023; Vachula et al., 2021).

Boxplots depicting the L:W values of experimental charcoal produced from plant taxa native to the southeastern United States of America (USA). Box outline colors illustrate the growth habits of the plants and the box fill colors depict the plant tissues (Table 2). Horizontal dashed red lines in the background note L:W values of 3.5 and 2.5 (Vachula et al., 2021). Vertical dashed gray lines divide the boxplots by plant group (i.e. monocot, dicot, and gymnosperm). Taxa were only plotted as trees if that is their only growth habit (Table 2). An average of 50 particles were quantified per sample (tissue).
Experimental charcoal L:W measurements varied between plant taxa and tissues (Table 2 and Figure 1). An ANOVA test of the L:W values of monocot, dicot leaves, dicot wood, gymnosperm wood, and gymnosperm needles supports that the population means of the fuel type groups are different from one another (p < 0.001). Likewise, if we remove saw palmetto L:W values from the monocot group before conducting the same analysis, the ANOVA still supports the population means of the groups to be different (p < 0.001). When aggregated by fuel types, the L:W values of monocot graminoids (median = 3.47, interquartile range (IQR) = 6.02) were greater than dicot or gymnosperm fuels (Figure 2). The results of our Tukey HSD test found that monocot charcoal L:W values were statistically distinguishable from those of dicot and gymnosperm woods, leaves, and needles groups (Figure 2). Within the dicot group, leaves produced charcoal with lower L:W values (median = 1.92, IQR = 1.33) than wood (median = 2.43, IQR = 3.00) The Tukey HSD test showed that the difference between the L:W values of dicot leaves and wood is statistically significant (Figure 2). In the gymnosperm group (e.g. wood and needles) the opposite was true with needles producing greater L:W values (median = 2.60, IQR = 3.22) than wood (median = 1.93, IQR = 1.44). The difference between the L:W values of gymnosperm needles and wood was also statistically significant, as supported by the Tukey HSD test (Figure 2).
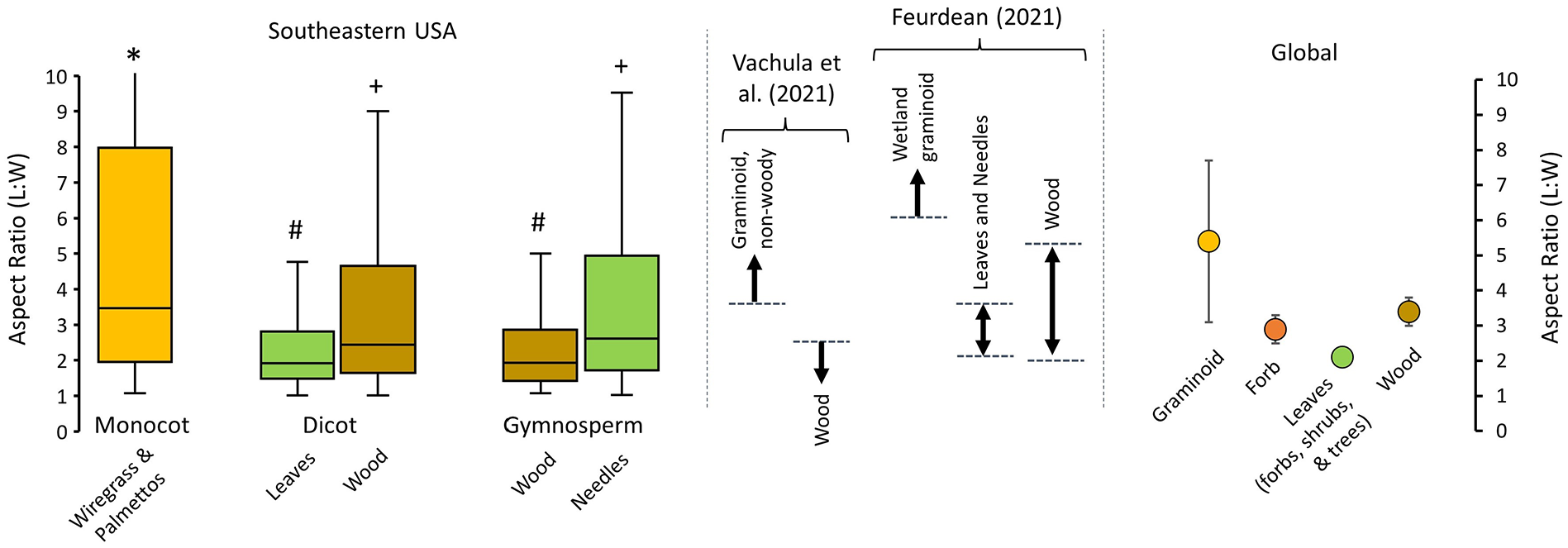
Experimental charcoal L:W values produced from the southeastern USA taxa, aggregated by fuel types (left), are in broad agreement with published mean L:W values of global experimental data (Feurdean et al., 2023; Vachula et al., 2021). The Tukey HSD test results are noted above each boxplot (*, #, and +) by showing the groupings of statistically distinguishable fuel types (α = 0.05 level).
In addition, our analyses provide metrics of charcoal L:W values produced from bark (dawn redwood) as well as seed pods (southern catalpa; Figure 1; Table 2). The bark sample L:W values (3.08 ± 2.30) broadly correspond to dicot wood (median = 2.43, IQR = 3.00) and gymnosperm wood (median = 1.93, IQR = 1.44) tissue samples (Figure 1). In contrast, the seed pod L:W values are relatively elevated (3.92 ± 3.52), highlighting the potential need to examine more seed pods, which can compose a component of litter fuel loads, in future experimental work.
Experimental charcoal produced from individual plants native to the southeastern USA exhibits less consistent L:W values than have been documented in previous research. In a systematic review of published (at the time) experimental charcoal L:W values, Vachula et al. (2021) found that graminoid, non-woody fuels generally have L:W values greater than ~3.5, whereas woody fuels tend to have L:W values less than ~2.5 (Figure 2). Independently and concurrently, Feurdean (2021) experimentally produced charcoal from plants of a diverse array of plant types and growth habits. Feurdean (2021) found that wetland graminoid charcoal L:W values were generally greater than 6.0, leaves and needles were between 2.1 and 3.5, whereas wood ranged between 2.0 and 5.2 (Figure 2). Some of the southeastern USA taxa and tissues we analyzed do not fit the molds that have been developed for charcoal L:W values in these previous experimental studies. For example, our sole graminoid taxa, wiregrass, had median L:W value of 2.92 (Table 2), in contrast to previous findings of graminoid elongated charcoal (Feurdean, 2021; Feurdean et al., 2023; Vachula et al., 2021). Similarly, several of our wood tissue samples, such as southern red oak (3.41 ± 4.51) and pin oak (2.62 ± 3.5), exhibit elevated L:W values that would not fit in the woody fuel category established in previous research (Vachula et al., 2021). Likewise, several of our leaf tissue samples, such as silky dogwood (1.55 ± 0.8) and sweetbay magnolia (1.71 ± 2.61), produced L:W values that fell below the expected intermediate range of previous research showing values of 2.1 ± 0.1 (Feurdean et al., 2023). Several of the gymnosperm needles we analyzed exhibit one of the more conspicuous differences with published charcoal morphometric analyses. Namely, the needles of eastern red cedar (4.55 ± 5.15), slash pine (2.74 ± 2.83), and pond cypress (4.18 ± 5.71) all produced charcoal with elevated L:W values that would typically be diagnostic of graminoid fuels.
Notably, Vachula et al. (2021) identified a methodological bias of charcoal L:W values in the published literature. Namely, they showed that charcoal produced in a furnace tended to have greater L:W values than charcoal produced in open flames. In contrast, Umbanhowar and McGrath (1998) found that open burns produced smaller charcoal with higher L:W values than did charcoal produced in a furnace. This disconnect could be due to the sieving of samples by Umbanhowar and McGrath (1998). As it pertains to the analyses of this study, this bias would mean that the L:W values of charcoal produced from plants native to the southeastern USA have muted ranges relative to if they were produced in a furnace. Therefore, our use of open burning to produce charcoal further underscores the differences observed between the southeastern USA samples and many of those published in previous research (e.g. Table 1).
Despite the heterogeneity of the charcoal L:W values of individual taxa and/or plant tissues, in aggregate our southeastern USA samples broadly agree with published ranges. Indeed, our monocot samples (Figure 2; median = 3.47, IQR = 6.02) generally agree with the recently published global L:W range for graminoids (Feurdean et al., 2023). Likewise, our dicot leaves (median = 1.92, IQR = 1.33) and wood (median = 2.43, IQR = 3.00) samples correspond to the published L:W ranges for global wood (3.4 ± 0.4) and leaf (2.1 ± 0.1; inclusive of leaves from forbs, shrubs, and trees) samples (Figure 2). However, our gymnosperm wood and needle samples are somewhat different from published ranges for global plants. Our gymnosperm wood (median = 1.93, IQR = 1.44) and needle (median = 2.60, IQR = 3.22) samples have lower and higher L:W values than published global ranges, respectively (Figure 2). Notably, however, these global estimates do not distinguish between dicot and gymnosperm wood nor leaves (e.g. leaves vs needles) because differences between these groups and tissues have not been previously observed (Vachula et al., 2021). The differences we observe between our gymnosperm charcoal and global L:W ranges have (1) potential paleofire utility for the southeastern USA, (2) provide perspective on the cause of elongated charcoal, and (3) highlight the need for more experimental work. We will discuss each of these implications in the following sections.
Implications of our experimental work for paleofire applications in the southeastern USA
Our analyses provide an informative perspective for paleofire research in the southeastern USA. The bulk of experimental charcoal studies analyzing individual taxa have focused on the Palearctic biogeographic realm (Table 1), and our study is the first to focus on taxa from temperate broadleaf and mixed forest as well as temperate coniferous forest biomes in the Nearctic realm. Similarly, this is one of few studies to focus on a subtropical climate region. Although previous research has used L:W values of charcoal to interpret fuel type changes from paleofire records in the Nearctic (Blarquez et al., 2018; Brown et al., 2017; Frank-DePue et al., 2023; Leys et al., 2017), they have relied on the experimental charcoal insights derived from aggregate mixtures of plants (Umbanhowar and McGrath, 1998) or other regions of the world (Vachula et al., 2021). Therefore, our data provides novel resolution of taxa-level charcoal L:W values and affords empirical basis for paleofire interpretations in the southeastern USA.
Our findings of elevated L:W values produced by non-graminoid taxa serve as a cautionary tale for paleofire interpretations in the region. When aggregated by fuel types (Figure 2), our data show that the wood of dicots and gymnosperm needles produce L:W values which can fall into the range of L:W values that is typically diagnostic of graminoids (Feurdean et al., 2023). To this end, paleofire practitioners working with samples from the southeastern USA should be especially cautious of interpreting high L:W values as solely representative of graminoid fuels. Qualitative charcoal morphological appearances can thereby provide an additional perspective and means of identifying graminoid fuels (Enache and Cumming, 2006; Frank-DePue et al., 2023; Krause et al., 2019; Mustaphi and Pisaric, 2014). Our finding of high L:W values for the wood of dicots (median = 2.43, 75th percentile = 4.66) and gymnosperm needles (median = 2.60, 75th percentile = 4.95) indicates that roughly half of the charcoal particles produced from these fuels have L:W values greater than 2.5. The L:W thresholds of 1.7, 2.0, 2.5, and 3 have each been used to distinguish grass versus woody fuels in paleofire research in other regions of the world including the Tibetan Plateau, central Africa, Quebec, Italy, Malta, Iberia, and southern Africa (Vachula et al., 2021). This juxtaposition means that practitioners should be cautious when interpreting paleofire L:W values, especially on long timescales when vegetation and ecosystem changes which could mimic the modern biogeographical heterogeneities of L:W values. Additionally, paleofire practitioners should consider other proxy data to further inform their interpretations. For example, comparison of charcoal L:W values with sedimentary pollen, phytoliths, or biomarkers could provide invaluable additional perspective with which to interpret sedimentary charcoal data (Jensen et al., 2007; Lupien and Sliwinska, 2024; Wen et al., 2020).
The differences of charcoal L:W values between dicot and gymnosperm tree tissues observed in our data, combined with qualitative morphology changes, could provide a basis for paleofire fuel interpretations that would be uniquely informative in the southeastern USA. Although this region was ice-free during the last glacial period, its Holocene vegetation development is generally characterized by a transition from conifer to deciduous dominated forests in association with the retreat of the Laurentide Ice Sheet (Bernabo and Webb, 1977; Krause et al., 2019; Overpeck et al., 1992). In combination with knowledge gained from pollen data, the charcoal L:W differences between the woods and leaves/needles of deciduous (dicots) and coniferous (gymnosperm) taxa (Figure 2) could thereby provide a means of distinguishing these Holocene fuel types in paleofire records. Although the L:W value ranges do overlap (Figure 2), additional qualitative morphological characteristics (e.g. charcoal morphological classification, plant tissue trait identification) would make it relatively straightforward to distinguish deciduous leaves, coniferous needles, and wood (Enache and Cumming, 2006; Mustaphi and Pisaric, 2014). In this way, charcoal morphometry could provide a more nuanced understanding of the role of fire in the Holocene shift from coniferous to deciduous forest in the southeastern USA. Further, this approach demonstrates the previously explained utility of multi-proxy approaches.
Getting at the cause of elongated charcoal
Previous research has speculated whether charcoal L:W values are controlled by growth habits, phylogenetic relationships or associated structural and chemical differences between plants of shared/differing ancestry. Previous experimental charcoal L:W values have generally supported a phylogenetic root to differences between plants, with Vachula et al. (2021) demonstrating that elevated L:W values (i.e. >4) were related to their shared possession of predominantly unidirectional veins and/or vascular bundles, as well as the dominance of cinnamyl in their lignin (Hedges and Mann, 1979; Umbanhowar and McGrath, 1998), all likely representing monocot synapomorphies. Notably however, not all monocots exhibit high L:W values. Feurdean et al. (2023) identified the disconnect of high and relatively low L:W values in graminoid charcoal to correspond to wetland and dryland taxa, respectively, which could also reflect evolutionary differences in physiology and tissue composition.
The plant taxa and tissues that we sampled provide two unique but conflicting perspectives on the root cause of elongated charcoal particles. First, our sampling of monocots shows that regardless of growth habit (shrub vs tree morphotypes), monocot charcoal can exhibit high L:W values (Table 2; Figure 1). However, as our Wiregrass sample and previous research shows, not all monocot charcoal exhibits high L:W values (Feurdean, 2021; Feurdean et al., 2023; Vachula et al., 2021). Therefore, these data suggest that elongated charcoal is likely at least somewhat phylogenetically driven and could be independent of plant growth habit.
Second, the elevated charcoal L:W values produced from several conifer needles provide a unique opportunity to test the role of evolutionary history on elongated charcoal. Although several experimental studies have previously noted L:W values produced from non-monocot, non-graminoid taxa that were higher than might be expected (e.g. greater than ~2.5), they have been overlooked as isolated anomalies within individual studies (Crawford and Belcher, 2014; Feurdean, 2021; Ogura, 2007). In contrast, our experimental burns identified several non-monocot, non-graminoids that produce charcoal with L:W values exceeding ~2.5, beckoning further investigation. Our analyses, in conjunction with published L:W data, allow for assessment of whether phylogenetic signal may influence the elongated nature of conifer needle charcoal. When we compare conifer needle L:W values, we find little relationship with known evolutionary relationships (Figure 3). This finding stands in contrast to previous support for phylogenetic control of elongated charcoal produced from monocots. Further work is therefore needed (and will be discussed in the following section) to fully disentangle and understand the cause of elongated charcoal.
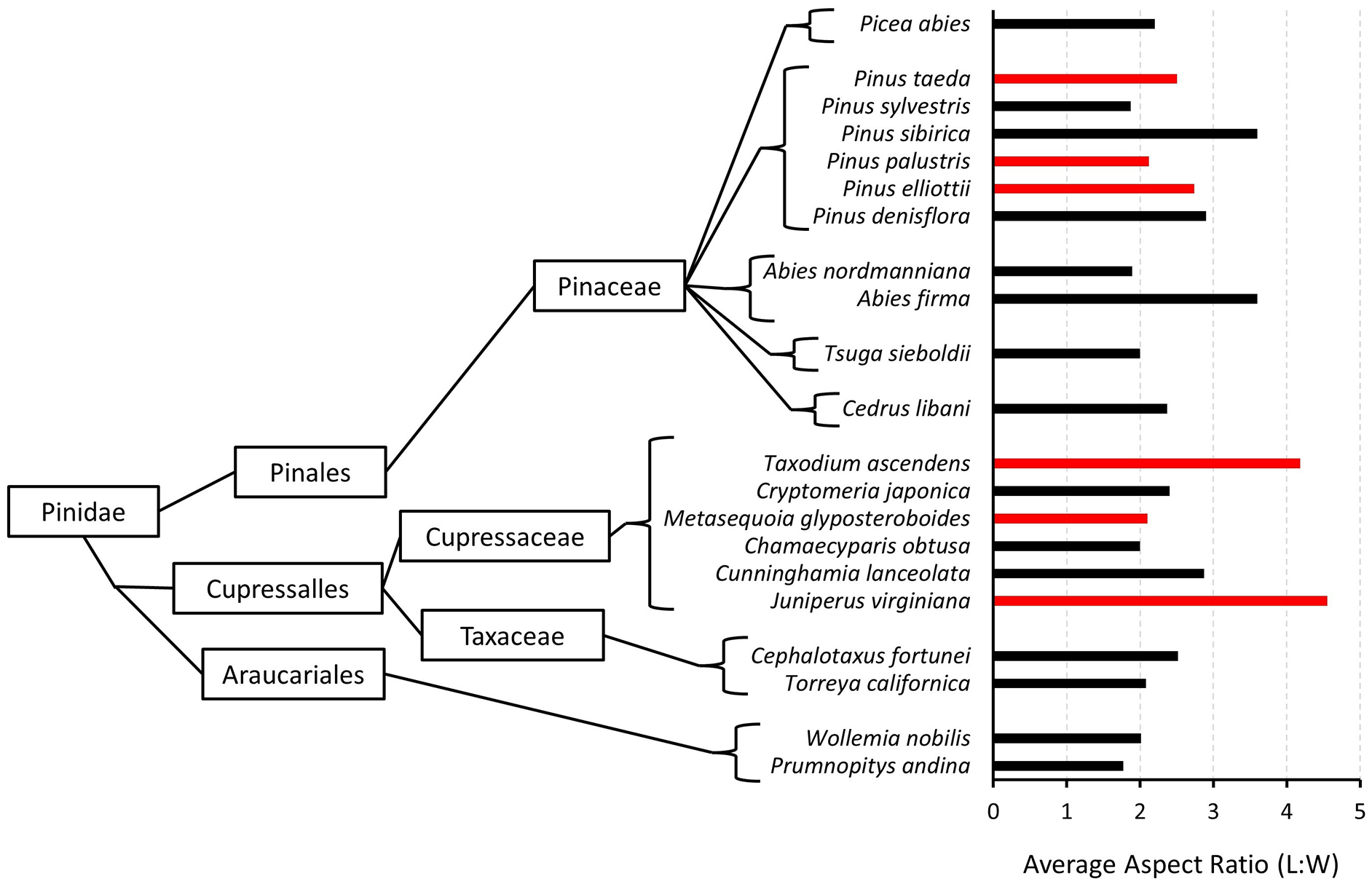
Cladistic relationships between conifer taxa and the L:W values of charcoal produced from their needles. Red bars note samples analyzed in this study and black bars note those from other publications (Crawford and Belcher, 2014; Feurdean, 2021; Ogura, 2007).
Paths forward for charcoal morphometric analyses
A notable result from our experimental burns was that despite the variety of plant types (e.g. dicot, gymnosperm) and growth forms (e.g. shrubs, trees) burned in our experiments, the resulting charcoal particle sizes were generally consistent, with the average feret diameter of charcoal particles for each sample ranging between 73.2 and 109.9 µm (Table 2). Further, there were not significant differences between the mean feret diameters of monocots, dicots, or gymnosperms (p > 0.1). However, the median L:W values and mean feret diameter of each sample were positively correlated (r = 0.43; p < 0.01). Although not directly analogous to a particle size bound that would result from sieving particles, the feret diameter (greatest distance been any two points of the edge of the particle) provides a relatable metric to sieved size fractions (Crawford and Belcher, 2014; Snitker, 2020). Therefore, our findings of average feret diameters less than 125 µm is striking as this size range would literally be washed down the drain during what is typically termed macroscopic charcoal analysis (Vachula, 2019). Sieving samples is notably the norm in experimental charcoal production studies (Table 1), as is the case for applied paleofire research (Vachula, 2019). Indeed, sieving of sediment samples in paleofire research is often conducted to isolate more locally-sourced, coarser charcoal particles from more regionally-sourced, smaller particles (Clark et al., 1998; Patterson et al., 1987). Similarly, larger particles are generally easier to quantify while recording similar variations as smaller size fractions (Whitlock and Millspaugh, 1996). Our data show that these methodologies could be missing out on the bulk of the size distribution of charcoal produced by open burning, which has important implications for relating measured charcoal production rates to sieved sedimentary charcoal records (Feurdean, 2021; Pereboom et al., 2020).
Further experimental charcoal research is thereby needed to fully characterize the impact of combustion parameters on charcoal morphometrics. Previous work has shown that L:W measurements can be impacted by sieving (Crawford and Belcher, 2014), as have the relative production rates of charcoal by taxa (Feurdean, 2021; Feurdean et al., 2023; Pereboom et al., 2020). Our finding of a positive relationship between L:W values and feret diameter further reinforces the need to consider particle size as a complication for interpreting downcore L:W values. However, there has been little examination of how analytical size foci compare to the size distributions of charcoal produced by plants.
Although our analyses provide a needed addition to the body of published experimental charcoal morphometric data, important sampling gaps persist. Indeed, despite the addition of 20 new taxa, in addition to new types of plants and tissues, further work is needed to more comprehensively examine the possible variability and sources of charcoal that could be encountered in paleofire records. For example, experimental studies have only examined plants from three of the eight global biogeographic realms (Table 1). To this end, we echo the call made by Feurdean et al. (2023) for future experimental charcoal work focusing on tropical grasslands. We would also underscore the need to experiment with plants from tropical, subtropical, and warm temperate zones more broadly, as their diversity might be expected to produce similarly diverse charcoal morphometric characteristics. In addition, any future efforts to extend these fire proxy records beyond the Quaternary and to deeper-time contexts may depend on more robust sampling of plant taxa from such environments, given the frequent intervals of greenhouse climate conditions and relaxed latitudinal temperature gradients during those older time periods (Scotese et al., 2021). Our results show that additional analyses of diverse plant taxa and tissues provides resolution of charcoal morphometric variability which undermines potentially assumed universality.
Our results emphasize the need for site-specific experimental charcoal approaches to enable robust paleofire applications of charcoal morphometry. Akin to previous findings of experimental charcoal studies focusing on new ecosystems (Feurdean et al., 2023; Pereboom et al., 2020), our analyses show that by characterizing the charcoal produced by previously untested taxa, additional understanding can enable paleofire interpretations. As we show for charcoal produced from dicot and gymnosperm wood, leaves, and needles of taxa native to the southeastern USA, new insights can be provided for paleofire applications through the examination of experiment charcoal.
Conclusion
This paper presents morphometric data (L:W) for charcoal produced from plant taxa native to the southeastern USA. In addition, we provide novel resolution of several plant taxa, growth habits, and tissues that have not before been analyzed in experiment charcoal research. We find that our charcoal samples from plants native to the southeastern USA exhibit more variable L:W values than have been documented in previous research, but that when aggregated, they broadly correspond with published L:W value ranges. Notably, our data show that there are some non-graminoid taxa which produce charcoal with elevated L:W values, which serves as a cautionary tale for paleofire interpretations in the region. We also note that differences of charcoal L:W values between dicot and gymnosperm tree tissues could provide a basis for paleofire fuel interpretations that would be uniquely informative in the Holocene of the southeastern USA, namely the transition between conifer to deciduous dominated forests. More broadly, our data provide conflicting perspectives on the cause of elongated charcoal particles, highlighting the need for further research on this topic. Additional experimental analyses of plant taxa from previously overlooked regions are needed to more comprehensively catalog the variability and diagnostic ability of charcoal morphometric data. Last, our study underscores the need for site-specific experimental charcoal approaches to enable robust paleofire applications of charcoal morphometry.
Research Data
sj-xlsx-1-hol-10.1177_09596836241274975 – Research Data for Morphometric characteristics of charcoal produced from plants native to the southeastern United States of America (USA)
Research Data, sj-xlsx-1-hol-10.1177_09596836241274975 for Morphometric characteristics of charcoal produced from plants native to the southeastern United States of America (USA) by Richard S Vachula, Thomas M Cullen, Matthew R Galinger, J Conner Welch, Jeremiah Battaglia, Dallyn Smith and Matthew N Waters in The Holocene
Footnotes
Author Contributions
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project was supported by start-up funds from Auburn University. DS was supported by the NSF CASE-REU program (Award 1950304). MG was supported by NSF-NRT (Award 1922687). We thank two anonymous reviewers for their helpful comments and improvements to this manuscript.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.