
Abstract
Taora (RIU), a rockshelter located on the northern coast of New Guinea, was first occupied shortly after sea level stabilisation ~6800 cal BP. Re-excavation of this site in 2004 to investigate claims of early pig domestication and pottery-use uncovered an uncharacteristically large assemblage of bone technology for the region. Here we present the analysis of that osseous artefact assemblage—including some 464 tiny bi- or unipoints alongside six plant fibre-working tools and numerous bone blanks for the manufacture of the projectile points. Analysis of the morphology and use wear present on the points indicates that they likely tipped multi-pronged fishing spears. This collection, and its association with abundant fish remains, supports previous notions of changing lifeways during the mid-Holocene and provides insight into the central role of organic technologies in Australasia during that period.
Introduction
Archaeological investigation of New Guinea’s rich human past continues to lag behind the intensive research interest seen in the smaller Indonesian islands to the east. Consequently, our understanding of the peopling of this large island, along with the technological and cultural diversity and change which was enacted across its diverse and often challenging landscapes remains patchy at best. This situation is particularly true of New Guinea’s coastal areas, which underwent considerable changes following deglaciation at the end of the Pleistocene, followed by rising sea levels reaching a highstand between 8000 and 6000 calBP and tectonic events causing uplift during the later Holocene. Little is known about deep human history across the island’s vast coastal lowlands relative to the Highlands, in part due to these restructuring events which obscured old landscapes and their archaeological sites as well as creating new coastal areas.
The north coast of New Guinea is centrally important to understanding Holocene developments in New Guinea and nearby Melanesia. Archaeological research along the coast of the Sepik region has given rise to the ‘sleeping giant’ model of regional prehistory (Golitko et al., 2024; Terrell, 2004). Briefly, this theory posits that the sea-level rises of the Holocene restructured the rocky, unattractive coastal landscapes of northern New Guinea into highly productive mangrove, estuary, and coral reef environments — leading to massive population growth, sedentism, increased technological complexity, and ultimately, intensified social organisation that formed the basis of the long-distance networks observed historically throughout the region.
Near Vanimo in western Sandaun Province of Papua New Guinea (PNG), archaeological research within caves and rockshelters has revealed a similar sign of local human activity during the Holocene (Gorecki et al., 1991; O’Connor et al., 2011). Here, sea levels reached their maximum level around 6800 years ago before stabilising, creating new distributions of marine or estuarine environments in close proximity to sites which had been inhabited for tens of thousands of years prior. Preliminary analyses of fauna from some of these sites, such as Lachitu which was inhabited prior to the Last Glacial Maximum (LGM), suggest a strong and increased influence of marine animal foods—molluscs, fish, and crustacea—in the Vanimo Coast diet at this time.
To date, however, very little is known about the procurement of these resources or the technological basis for doing so. Here we present the analysis of a large assemblage of bone artefacts recovered from Taora Shelter located on the Vanimo Coast. Numbering some 526 bone artefacts in total, this deposit is unique in its abundance and offers fresh insight into mid-Holocene osseous technologies and lifeways within these local coastal environments and the marine economy of people who lived within them.
Taora Shelter
Taora shelter (PNG National Museum & Art Gallery Site code RIU) is located not far from the present-day town of Vanimo (Figure 1). It lies 5 km west of the better known site of Lachitu, both of which sit within karstic limestone terrain at the foot of the Oenake Range. The area is tectonically active with net uplift rates >1.2 m/1000 years and likely movement associated with Holocene seismic events (Tudhope et al., 2000). Currently, Taora is about 9–12 m above sea level and 450 m inland on the seaward side of a large limestone block outcrop. Excavations at the site have demonstrated that the main phase of human occupation coincided with an active shoreline and back-beach abutting the outcrop around 6800 years ago (Gorecki et al., 1991; O’Connor et al., 2011).
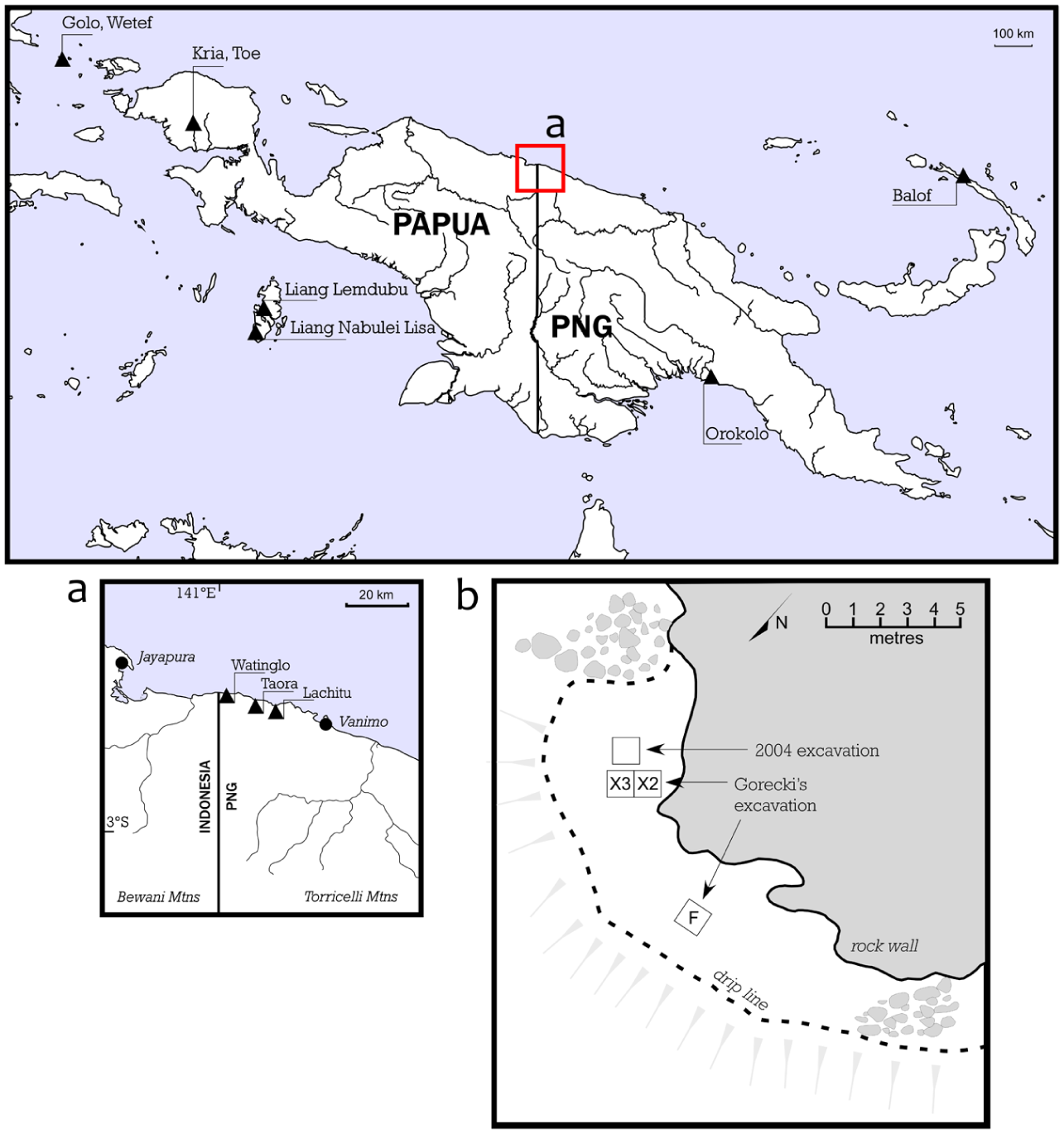
Location of Taora on the north coast of Papua New Guinea (PNG) and other sites mentioned in text; (b) location of 2004 and Gorecki excavations at Taora.
Taora shelter was first excavated by Paul Gorecki and team in 1990, who opened a 1 m2 pit (F4) on one side of the site and a 2 × 1 m trench (X2 and X3) on the other (Gorecki et al., 1991; Figure 1). Gorecki et al. (1991) obtained charcoal ages of 5860 ± 90 yr BP (ANU 7606) and 6120 ± 190 yr BP (ANU 7605) from a hearth directly overlying beach sand at the base of trench X2 and shell dates from 90 cm above the base of 5370 ± 90 yr BP (ANU 7702) for X2 and 5410 ± 90 yr BP (ANU 761 1) for F4 demonstrating that the deposit accumulated very rapidly. The upper deposit containing ceramics in X2 was dated on marine shell to 2250 ± 70 yr BP (ANU 7701). Significantly, Gorecki et al. reported that pig remains were present throughout the deposit.
Taora was re-excavated in 2004 by Sue O’Connor and her team in order to investigate the validity of the claim for a Mid-Holocene appearance of pig and pottery within the sites deposit (O’Connor et al., 2011). This more recent excavation was restricted to a 1 m2 pit close to Gorecki’s squares X2-3 (Figure 1) which extended down to 178 cm below the surface where sterile beach sands were reached. Both stratigraphy and dating were found to be broadly consistent with that reported by Gorecki et al. (1991). This excavation was able to confirm that Taora was first occupied shortly after sea levels began to recede and stabilise from ~6800 cal BP. It also found the presence of intermixed beach sand and terrestrially-derived sediments with cultural material in the lowest levels indicating that the site was occupied as soon as it was uplifted, and may have been subject to occasional marine incursions during the earliest phase of its occupation.
Stratigraphy in the 2004 excavation features shelly well-sorted beach sands in lower levels, indicating over-wash on a narrow cliff-base berm. Cultural material is present in the beach sand units, co-associated with coral blocks, suggesting initial occupation coincident with the maximum Holocene transgression on this up-lifting slope. Subsequent relative sea level fall and tectonic uplift has left the site separated from the modern shoreline by forest. Upper units are more clastic and shelly throughout and had accumulated through cultural and natural deposition including sediment from rock-fall and in-wash. Small scale bioturbation involving crabs and other animals was noted during excavation, and there was bioturbation where tree and vine roots had reached several metres into the deposit, but such deep bioturbation is uncommon and localised.
Radiocarbon dating demonstrated that the Taora deposit accumulated rapidly, and predominantly over a very short period of time between 6800 and 6000 cal BP. A total of 17 AMS dates were obtained, five on marine shell, one on charcoal and one on Canarium nut and all fall into a narrow time range between c. 6800 and 6000 calBP (Table 1). A date on charcoal from Spit 4 obtained subsequently by Beaumont et al. (2018) aimed at dating the densest concentration of pottery in the Taora (Wk 47060) and returned an age of 495–323 cal BP.
Distribution of bone artefacts from 2004 excavation alongside sediment weight excavated and bone, lithic and potsherd finds (after O’Connor et al., 2011).
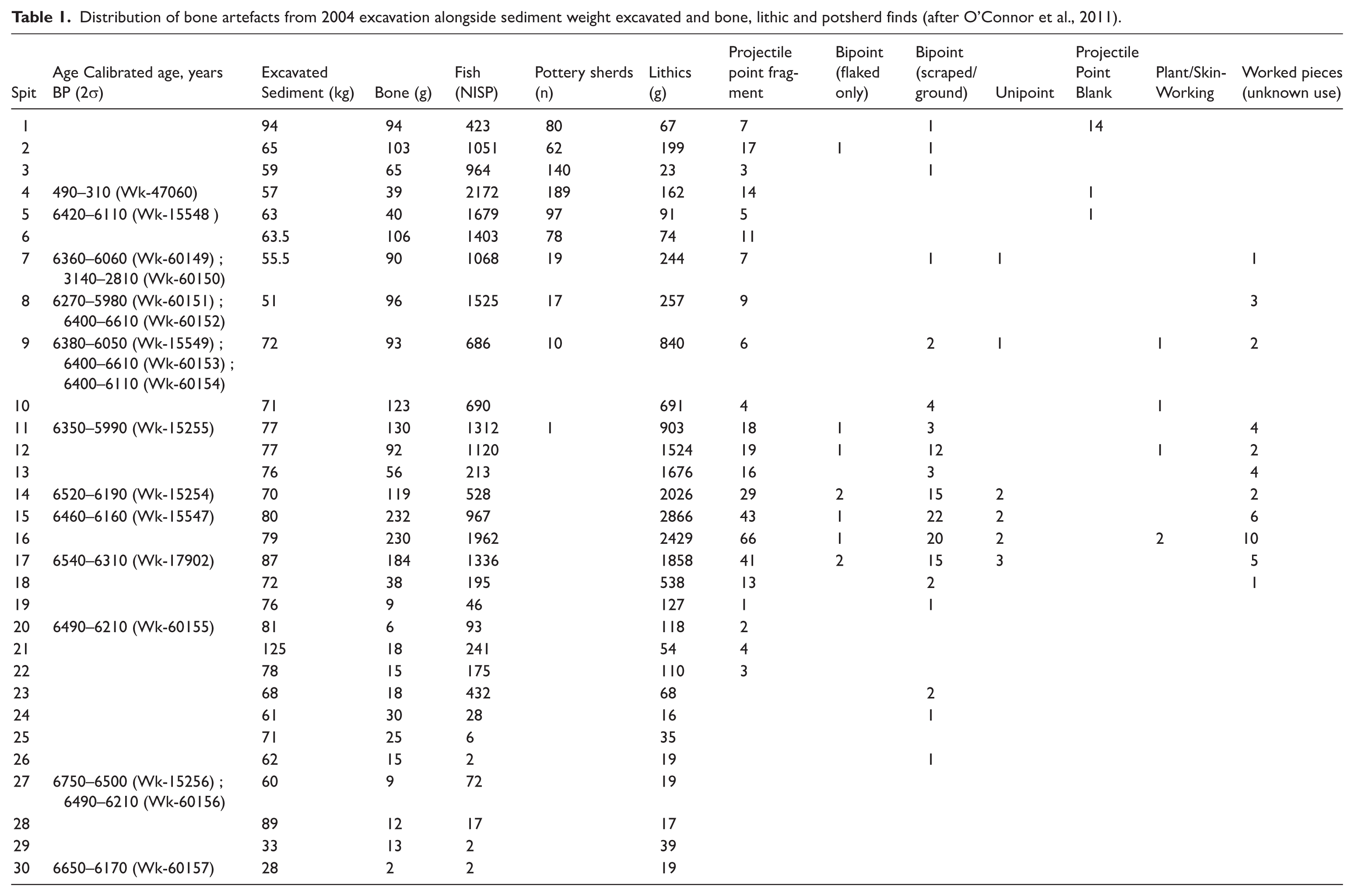
Significantly, the small 2004 excavation also recovered a very large assemblage of bone artefacts, an artefact category not mentioned by Gorecki et al. (1991) as being found at Taora. These artefacts are the subject of the following analysis.
Methods
The 2004 excavation proceeded using excavation units which averaged between 2 and 5 cm in depth (Figure 2). Samples of shell and charcoal were collected for dating from in situ locations within excavation units and from the sections so that units could be correlated with any stratigraphic changes in the section undetected during excavation. All excavated deposit was wet-sieved through a 1 mm mesh in combination with flotation to the 0.25 mm level which eliminated any possibility that tiny sherds, bones, or teeth could be overlooked and also to ensured maximum recovery of charcoal and other macrobotanic material. All material from both the heavy fraction and flot were then dried and a preliminary sort on the heavy fraction undertaken in the field. The residue was then returned to the Australian National University for final laboratory sorting (see Fairbairn, 2005). All marine shell samples submitted for dating were tested for recrystallisation.
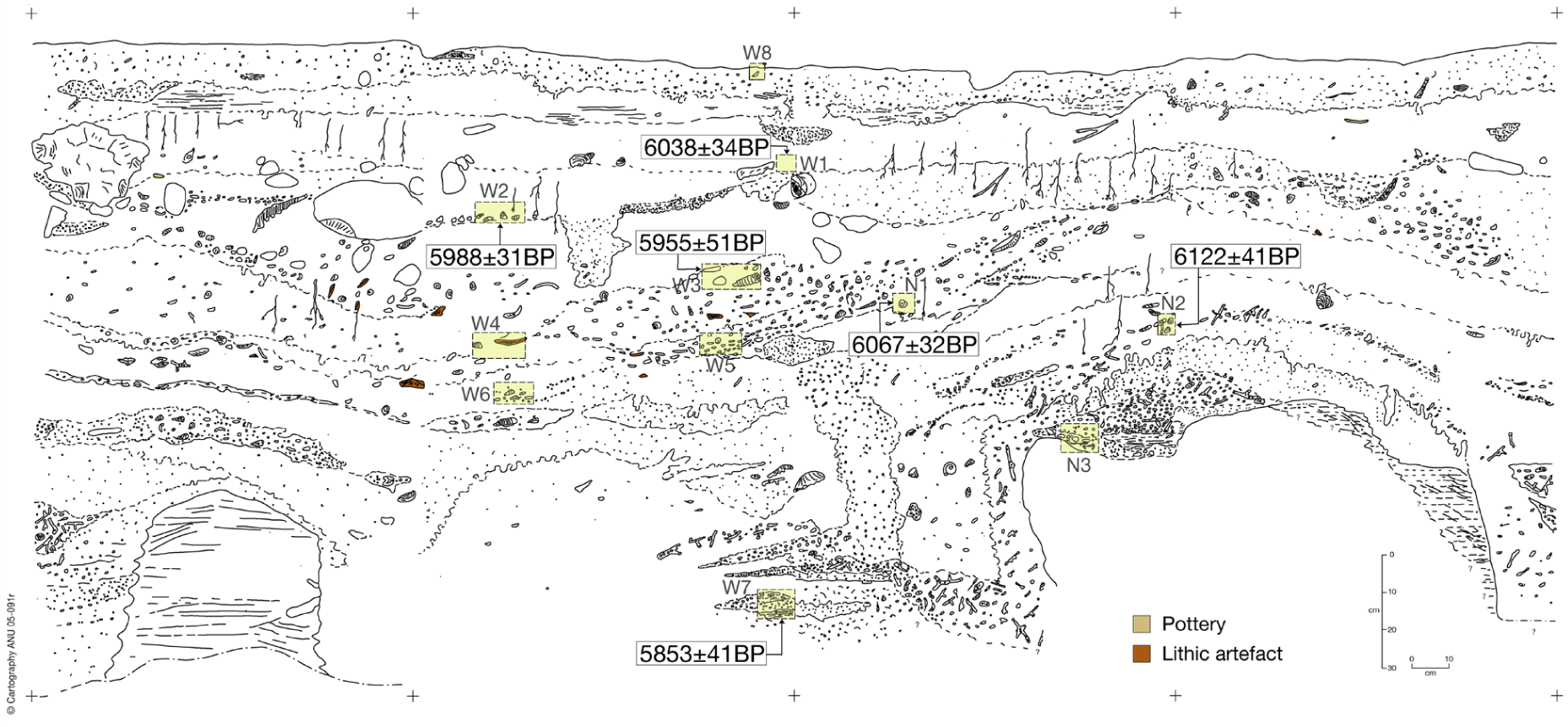
Section of the 2004 excavation at Taora, with positions of the dated in-situ samples. Image drawn by Anthony Barham.
Both shellfish and fish bone is abundant throughout the deposit, including the presence of crab and urchin, though vertebrate fauna (including tree kangaroo) is most abundant in the upper half of the deposit. Pig (Sus scrofa) bone was limited to the top two spits. Fragments of human bone were present in most spits down to spit 10. Lithics were present throughout the stratigraphy, while pottery sherds (n = 693) were mostly restricted to the top six spits. The few which were found within the lower mid-Holocene levels are believed to have moved from the denser deposit above (Beaumont et al., 2018; Table 1). As such, the Late-Holocene occupation, indicated by the abundance of potsherds in the upper 9 spits, sits directly on the older ~6800–6000 year old materials, and the previous interpretation of pigs and pottery in the Mid-Holocene is erroneous (O’Connor et al., 2011).
In terms of the analysis of the bone artefacts, metrics for each artefact were collected using Mitutoyo digital callipers, the jaws covered in a layer of plastic coating to prevent damage to the objects (this thin coating does not alter the measurements taken as the callipers are able to be zeroed before use). Metrics gathered follow standards set out in previous analysis of osseous pointed technologies (e.g., Knecht, 1991;, 2023Pétillon, 2006; Langley 2018, Langley 2023). Artefacts were classified as ‘intact’ when both distal and proximal extremities were present in their majority, or otherwise as ‘fragments’ with the section (distal, mesial, proximal, etc.) recorded. Artefacts were classified according to their overall morphology in the first instance, and as belonging to an ‘unknown’ type if too fragmentary to be sure of their form. We prefer to err on the side of caution, so pieces which may have been flaked but did not present enough evidence of working to be certain of their technological status are not included in the statistics below.
Taphonomic alterations, manufacturing traces, and use wear observations were based on comparison with published works (e.g., Álvarez et al., 2014; Bach et al., 2023; Buc, 2011; Buc and Loponte, 2007; Bradfield, 2015a; Campana, 1980; Desmond, 2022; Fernandez-Jalvo and Andrews, 2016; Fisher, 1995; Houmard and Sidéra, 2024; Langley et al., 2022; Legrand and Radi, 2008; LeMoine, 1991, LeMoine,1994, 1997; Newcomer, 1974; Pal et al., 2020). Terminology for description of these features follows standards set out in this archaeological literature. Interpretations of artefact functions were based on the combination of morphology, use wear distribution and type, residue distribution and type and comparison to experimentally produced and ethnographically collected tools of known function.
Results
In total, 526 bone artefacts were identified from the 2004 excavated 1 m2 pit (Table 1). Of these pieces, 464 are identifiable as either intact or fragmented pieces of small projectile points. Six fragments of possible plant- working tools were also identified, along with 16 possible blanks for the manufacture of the projectile points. Another 40 pieces were found to display substantial traces of working in the form of flaking and scraping or use (chipping, crushing, rounding, polish, etc.) but were too fragmentary to confidently assign them to a tool form or possible function.
The bone technology is largely restricted to between spits 9 and 19 with a few found both below and above this central section of the deposit. Comparison of the stratigraphic distribution of bone artefacts to lithics shows that spits 9–19 preserved the bulk of material culture (Table 1). Spit 8 and above appears—from both the presence of much later dates and relative quantities of artefacts—to present a palimpsest of some mid-Holocene amongst a majority of more recent Late-Holocene material (i.e., pottery sherds). Furthermore, as O’Connor et al. (2011) have previously surmised, most of the deposit at Taora accumulated quickly between about c. 6800 cal BP and 6000 cal BP followed either by an occupation hiatus of several thousand years or the removal of deposit dated between c. 6000 cal BP and 2000 cal BP. As such, the bone artefacts described below primarily originate from the mid-Holocene use of the site. Owing to the short period of time represented (approximately 500 years) and the identified small scale bioturbation below spit 8, the bone artefact assemblage is treated here as a single entity. Having said that, several artefacts indicate from their manufacture that they are more recent in origin, indicating that the creation of bone points and tools at Taora certainly continued into the last few hundred years.
Before moving into describing the bone technology, it should be noted that overlying the manufacturing and use traces are several forms of taphonomic damage, the most extensive of which has been caused by insects (Figure 3). The main culprit appears to be termites of which there are 45 species in New Guinea (Bourguignon and Roisin, 2011), but may also include some large ant, beetle, or other insect activity based on the comparison of the observed damage to that in the published literature (Backwell et al., 2021; Fernandez-Jalvo and Andrews, 2016; Parkinson, 2023; Watson and Abbey, 1986). As pointed out by Backwell et al.’s (2021) review of insect damage to bones, relatively little attention has been paid to the specific traces of distinct species’ damage, this situation also unfortunately the case in the Australasian region. As such, while we can be certain that the marks are the product of insect activity, and most likely termites and beetles, being more specific is currently impossible.
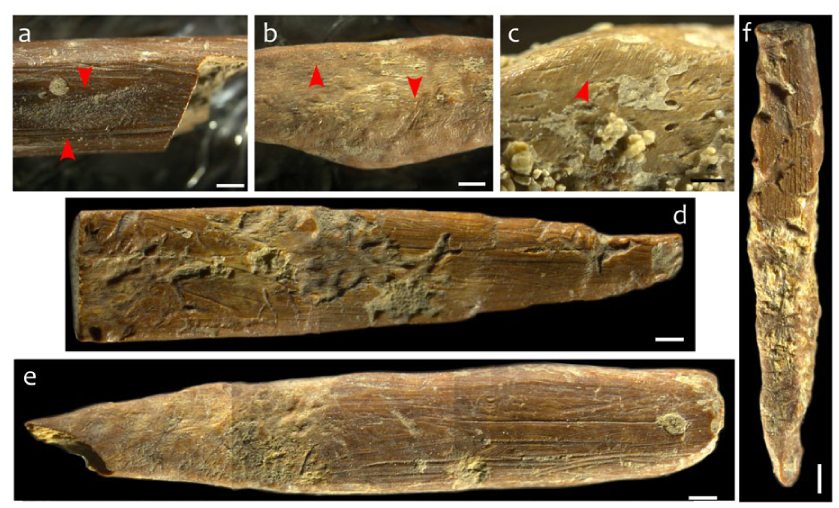
Insect damage to the Taora bone artefacts. (a) Surface damage from unknown beetle(?). (b and c) Fine parallel striations caused by termites, beetles or other insects. (d–f) Examples of damage caused by termites. Scale bar = 1 mm.
Figure 3 provides examples of the identified insect damage which ranges from affecting only small sections of an artefact surface, through to extensive alteration of the surface (Figure 3d–f). To summarise the observed traces, the damage appears in the form of star-shaped and curvilinear grooves which begin in one location and then spreading out from that initial contact site, stripping away the outer layer of bone (Figure 3d–f). Within these sections where a layer of bone has been stripped away, the surface is covered by tiny, shallow, sub-parallel striations (Figure 3b). On other artefacts, slightly wider but still shallow parallel striations are visible (Figure 3c), and may be the result of dermestid beetles though further analysis is required to confirm (Backwell et al., 2021).
Projectile points
Some 88.2% of the bone artefact assemblage comprises small points which average around 27.1 mm in length (range between 13.6 and 46.9 mm) when intact (Figure 4). The interpretation of these artefacts as projectile points is based on the combination of morphology, size, evidence for fixed hafting of the proximal extremity, and use wear which is consistent with having been acquired via impact between a target and the projectile shaft (detailed below). Alternative interpretations, such as fishing gorges which are similar in their form, can be ruled out owing to the differences in hafting and use wear traces (Bradfield, 2019).

Examples of intact and near intact bone points recovered from Taora (2004 excavation). The four artefacts in the top row are examples of possible unipoints, while the remainder are examples of the bipoints.
The points have been made on fragments of mid-sized terrestrial bone, though a few appear to be on large avian or fish bone. Sources for this bone likely include white-striped dorcopsis (Dorcopsis hageni), along with other members of the Macropodidae and Phalangeridae as these species dominate the faunal assemblage. Manufacture appears to have occurred on site, with targeted flaking resulting in bone flakes (blanks) whose shapes mirror finished artefacts (compare finished points shown in Figure 4 with possible blanks in Figure 5). In fact, large numbers of bone splinters of this approximate shape and size were identified during faunal analysis but are not considered further in this work. Owing to the extent of modification and size of the pieces, it is impossible to identify the anatomical element utilised with certainty, though long bones appear to be preferentially selected and other elements which have thicker sections may also have been used.

Possible bone blanks for the manufacture of bone projectile points at Taora.
Creating a finished point from a blank could be restricted to flaking (at least nine points present clear traces of use but no other method of shaping), though the vast majority also present striations indicative of having been scraped into the desired form. Stone edges were evidently used, the striations varying in width, depth and cross-section owing to the uneven and rapidly changing edges of the stone tool as the scraping is undertaken (Figures 6a, b and 7g–j); Newcomer, 1974). Interestingly, two points have been shaped using a metal blade rather than stone (Figure 6c and d). The use of a metal blade is indicated by the long, straight, and fine longitudinal striations which entirely cover the interior surface of the shave scar (particularly evident in Figure 6d). The striations are regularly spaced and morphologically comparable across the shave scars, metal blades changing significantly less rapidly than those made on stone (Christidou, 2008; Cristiani and Alhaique, 2005). Both examples come from higher in the stratigraphic sequence (spits 1/2 and 6).

Shaping of bone points using lithic (a and b) and metal (c and d) blades. Scale bar = 1 mm. (a) RIU469, Spit 4. (b) RIU480, Spit 3. (c) RIU467, Spit 6. (d) RIU476, Spit 1/2.

Traces of manufacture and use observed on the Taora projectile points: (a–f) impact fractures and wear to distal extremities, (g) distal-mesial fragment of a bipoint with lithic scrape marks, (h) intact bipoint, (i) bipoint with bevel fractures to both distal and proximal extremities and (j) mesial-proximal fragment of a bipoint. Scale bar = 1 mm.
Bipoints are the dominant form, with 116 examples (intact enough to be confident of their shape) found alongside 11 points which might be more traditionally classified as unipoints (Figure 4). These latter 11 artefacts display horizontal or near-horizontal proximal extremities as opposed to the more or less acutely pointed proximal ends of the bipoints, however, these morphological differences are very minor and ultimately insignificant. All of these small points represent the same functional artefact, their morphology being the product of personal preference for size, symmetry and finish, and then the vagaries of use wear and rejuvenation cycles.
As is characteristic of projectile point assemblages (e.g., Arndt and Newcomer, 1986; Bergman, 1987; Knecht, 1997; Langley, 2015; Roper, 1979; Titmus and Woods, 1986), the vast majority of the Taora projectile point collection are mesial-proximal sections where the distal portion has fractured in use and is now missing (61.6%). A further 20.2% of the assemblage is made up of mesial or extremity fragments. Fractures evident on the distal extremity range between comparatively ‘minor’ crushing, rounding, and bevel and step fractures (e.g., Figures 7d, e and 8) through to larger splinter, bevel, snap, and step fractures (Figure 7a–c, f, i and j). Damage to proximal extremities are typically less destructive, usually restricted to chipping, crushing, and rounding (e.g., Figures 7h and 8), however, larger bevel and snap fractures are also well-represented (e.g., Figure 7i and j). This combination of damage to both the distal and proximal extremities of points is consistent with use as hafted projectiles (e.g., Bradfield, 2012; Langley, 2015; Langley et al., 2022; Pétillon, 2006; Pétillon et al., 2016).

Forms of use damage from less to more severe wear to distal and proximal edges using examples drawn from the Taora assemblage. Note that the proximal edge of fixed projectile points typically accrue wear slower than that of the opposing active surface.
Evidence for fixed hafting is found in edge-rounded, compression, and polished proximal halves, along with notches formed through the use of ligatures near the mid-sections and the aforementioned damage to the proximal tip (crushing, rounding, chipping; Figure 7h; Rots, 2005). Furthermore, fracture across the mid-section has been noted as a common fracture pattern where the point fractures at the height of the haft (Arndt and Newcomer, 1986; Bergman, 1987; Guthrie, 1983; Knecht, 1997; Pokines, 1998; Tyzzer, 1936).
The diminutive size of these points suggest that these pieces tipped either multi-pronged hand-thrown spears (which may or may not have had a line, the former then technically being a harpoon) or bow-launched arrows. Certainly, very similar bone bipoints have been identified as components of multi-pronged fishing spears in the Australian context (Langley et al., 2022, 2023) and comparisons to ethnographically-known Indigenous Australian fishing spears have previously been drawn in interpreting the much smaller collections of bone points found elsewhere on New Guinea and surrounding islands (e.g., Olsen and Glover, 2004; Pasveer and Bellwood, 2004). Similarly, bone-tipped arrows (including multi-pronged arrows) for taking fish, birds, and arboreal game are documented in this region for the recent past (Bulmer and Bulmer, 1964; Comrie, 1877; Hornell, 1923; Simmonds, 1959; Woodford, 1888).
From examination of Taora’s points themselves, it is impossible to differentiate between these two possibilities (hand-thrown spear or arrow tips). Examination of recent Papuan bone arrow-tips tend to be much longer and more sinuous, often with a hypodermic form that utilises the exposed hollowed medullary canal. Others are intricately carved from larger pieces of bone cortical section to feature multiple barbs themselves; and both varieties are typically used in warfare rather than hunting or fishing. However, this does not mean that earlier arrows could not have used smaller bipoint forms, since similar artefacts are believed to have tipped arrows in the wider Indo-Pacific region from a very early period (e.g., Langley et al., 2020). Conversely, while the dominant point forms at Taora strongly resemble those mounted in recent Australian Aboriginal multi-pronged fishing spears, equivalent ethnographic examples from mainland New Guinea seem to be essentially absent despite the island-wide use of hand-thrown spears in fishing. It is worth noting that a multi-pronged leister type of fishing spear (where each prong tip also has a sharp internal-facing barb) is used in this area of north-central New Guinea (Idris et al., 2021) as well as on Biak and in the Moluccas. Additionally, such fishing spears have also been seen in use ethnohistorically not far from Taora itself, on Wuvulu, an island north-northeast of the site (Edge-Partington, 1896). Each prong and its barb is today made from a single piece of metal, but in the past it is conceivable that bone was used to achieve a similar configuration.
Consideration of Taora’s faunal assemblage offers a potential way forward past this issue of launch method. While white-tailed dorcopsis (Dorcopsis hageni) and other medium-sized mammals are present (Thylogale browni and Phalanger orientalis in particular) within the site—fish, crab, and urchin dominate the faunal assemblage at Taora. These marine resources are particularly dense between spits 9 and 18, which coincides with the distribution of bone projectile points (compare Table 1 to Supplemental Table 1). Preliminary analysis of the fish remains found that a range of fishes were caught, including but not restricted to Balistidae (triggerfish and relatives), Scaridae (parrotfish), Acanthuridae (tangs, surgeonfish), Labridae (wrasses), Lethrinidae (emperor breams), Serranidae (sea bass and groupers), Muraenidae (moray eels), Elasmobranchi (rays, skates), Diodontidae (porcupinefish), and Mugillidae (mullets). Based on the size of these remains, and those of the numerous unidentified vertebral elements, the fish processed at Taora are estimated to have been between about 20 and 30 cm in length, though some larger specimens are also present. Given the dominance and range of near shore fish present at Taora, a technology—such as multi-pronged fishing spears—which was central to their capture should be represented within the archaeological material culture. Furthermore, such multi-pronged weapons can and are used to capture other forms of prey in New Guinea, three- or four-pronged spears recorded as being used to hunt small birds and mammals (Bulmer, 1968: 307–308). Consequently, we feel that the most parsimonious interpretation of the exceptionally large (for this region) collection of small bone projectile points is that they once tipped tip multi-pronged spears which were primarily used to catch near shore fish in shallow water or from canoes—but not necessarily only fish.
Possible plant-working tools
Also found were six artefacts which display morphological characteristics and use wear which differ from the projectile points (Figures 9 and 10). While also pointed, these distal-medial fragments of once longer tools are significantly wider and thicker than the projectile tips described above. Each has a sub-circular to oval cross-section at the distal tip which moves into either the natural morphology of a long bone (no’s 132, 179 in Figure 9 and no 166 in Figure 10) or a concavo-convex cross-section if made on a fragment of long bone (no 6 in Figure 9 and no’s 31 and 61 in Figure 10).

Three of the pointed tools possibly used in working plant materials. Numbers underneath each artefact is their unique analysis identifier. Scale bar = 1 mm.

Other three pointed tools which appear to have been used in working plant materials. Numbers underneath each artefact is their unique analysis identifier. Scale bar = 1 mm.
Each has fractured in antiquity with either a bevel, splinter, or snap fracture resulting in only the distal extremity being recovered (or identified). These artefacts feature the undulating surfaces and coarse striations produced by scraping the extremity into a point with a lithic edge and appear to be more ‘expedient’ in their creation than the often (but not always) carefully shaped projectile points described above.
Each of these artefacts displays pronounced crushing, chipping, and rounding to the distal edge with rounding and a high polish continuing from the tip down towards the mesial section for between 1.5 and 3 cm. Artefact 6 additionally presents a series of sub-parallel striations down the right side (Figure 9) which is reminiscent of use wear known to develop when working with plant materials and possibly rubbing coarse hide (Bradfield, 2015b; Buc, 2011; Buc and Loponte, 2007; Gates St-Pierre, 2007). While superficially similar to some of the termite damage observed in this assemblage (Figure 3), the striations on this artefact are deeper and more widely spaced suggesting they are the result of anthropogenic behaviour rather than insects. Overall, these few artefacts are indicative of plant- and possibly also hide-working at Taora site.
That the manufacture of plant-based objects was occurring at Taora should be expected as in New Guinea, as elsewhere, a multitude of containers, bags, nets, clothing, and more are recorded in ethnohistoric times and no doubt these all have a deep antiquity. Of particular interest here is Bulmer’s (1968: 315) observation in his ethnography of hunting in New Guinea, that ‘although all New Guinea peoples have quite elaborate techniques of netting used in the construction of bags and, often, personal garments, the use of nets in hunting seems mainly restricted to groups who also use nets for fishing, or at least live within relatively short distance of the coast’. It may be then, that the larger bone implements found at Taora were used in the construction of plant-fibre netting used in fishing.
Sillitoe (1988) further describes ‘needles’ made of hardwood or bone used by the Wola of the Southern Highlands of PNG which are pointed at one end, rounded at the other and usually perforated at the blunt end. These tools are up to 15 cm in length and are used for stitching rain capes and making string bags. Similarly, amongst the Meybrat in the Ayamaru region of western Papua, hollow unipoints made on bat bones are cited as being used for ‘sewing’ pandanus leaves (Elmberg, 1955: 14), as well as for sewing bags made of bark and umbrellas of palm leaves which were used for shelter, to sit or sleep on, and to carry foods or other things (Pasveer, 2005). Ethnographic accounts also provide background to the manufacture of plant-working tools. Sillitoe’s (1988: 65) ethnographic account of the Wola, noted that ‘for bone implements they prefer the bones of pigs, mapun wallabies (Thylogale or Dorcopsulus spp.) and the wings of tagem black flying foxes (Dobsonia magna), as these are particularly strong’ and that these bones are simply broken ‘to produce jagged points, sometimes paring them to give sharp tips’. This approach is exactly that seen on the Taora artefacts, including the presence of a bat ulna with evidence of scraping, though this example may be from butchery rather than tool manufacture and so was not included as an example of worked bone.
Another form of material culture that should be considered are decorative hairpins which have been ethnographically recorded as being made of wood, bamboo, and bone. These are pointed at both ends and usually quite long, reaching up to 15 cm and also made on flying fox wing bones, wallaby long bones, and pig ribs. Sillitoe (1988: 67) notes that the hairpins are ‘quickly made and just as quickly discarded after use’. It is worth noting that many of the worked bone artefacts that could not be categorised beyond having been worked (displaying scraping marks down their surfaces) are mesial fragments of long bones and may represent fragments of further plant-working tools or items such as these ethnohistorically described hairpins.
Discussion
The past 20 years has seen the publication of bone technology from a handful of Australasian archaeological sites (Table 2). Not one of these contexts, however, have produced the sheer abundance of osseous artefacts that was recovered from Taora (and this number recovered from a single 1 m × 1 m square). Such density of osseous artefacts is indeed unusual for this region, though whether this difference is a product of sampling methods and recognition, preservation conditions, or site use is yet to be demonstrated. However, the ongoing analyses of fauna from the other Vanimo Coast sites—Lachitu, Watinglo, and Paleflatu—(Helgen et al., 2010; O’Connor et al., 2011) are now sufficiently advanced to confidently identify that the number of points from Taora is exceptional also for the local area. Since Gorecki et al. (1991) make no mention of bone artefacts being found at Taora, it remains possible that the 2004 excavation simply happened to come upon a bone-working locality within the shelter. Alternatively, and perhaps more plausibly, is the difference in recovery methods used during the two excavations. The 2004 excavation at Taora utilised a 1 mm sieve diameter in the wet screening of all deposit from the excavation (O’Connor et al., 2011). Gorecki et al. (1991) do not discuss their excavation or recovery methods but it is likely that much larger meshed sieves were used and slender bone artefacts likely passed through the sieve.
Previously reported bone technology from Australasia.

Number of artefacts includes those reported as ‘complete’ or ‘fragmented’ (or similar) in the original publications. The maximum width and thickness for the Taora points are included here for comparison to the ‘mesial width and thickness’ reported by Pasveer and Bellwood (2004) and similar measurements by other reports. As the widest and thickest points on bipoints are usually their midpoint, these metrics should be broadly comparable.
The closest archaeological comparison of the Taora material is that found in Kria Cave, located on the Bird’s Head of New Guinea. From this location, Pasveer (2004, 2005) examined 92 bone artefacts along with eight utilised macropod teeth dating to between c.7000 and 4300 BP. As with the Taora artefacts, the majority of the assemblage was made up of small bipoints made on slivers of cortical bone or shaft bone fragments from mid-sized mammals (Dorcopsis wallabies, tree kangaroos, and cuscus), which were shaved, scraped, and/or ground into their final shape. Pasveer found essentially the same range and distribution of use wear, including crushing and small fractures to the distal and proximal edges, along with polish on the surfaces, though she interprets these artefacts as having been hafted onto handles as engravers or perforators for use on plant materials. She also identified five cortical bone fragments which she interpreted as possible blanks for the unfinished bipoints. Again, these blanks are very similar to those identified within the Taora assemblage. Another 16 unipoints are reported from Kria which are interpreted as barbs or spear or arrow tips, while the eight teeth displayed use wear consistent with having been used as drill bits. At nearby Toé Cave, only four osseous artefacts were recovered. Three of these artefacts may be small points for multi-pronged spears, and another utilised tooth makes up the fourth find.
Similar artefacts have also been described by Pasveer for sites located in the Moluccas, west of New Guinea. The largest assemblage was recovered from Golo Cave on Gebe Island, and includes 75 bipoints of similar size, morphology, manufacture, and use wear to the Taora, Kria, and Toé examples (Pasveer and Bellwood, 2004; Table 2). Additionally, Golo Cave also produced 18 spatulate-tipped points, along with 19 ‘attenuate unipoints’, these latter items being similar to the Taora artefacts identified as plant-working tools. As with the Taora tools, these 19 unipoints are larger than the small bipoints and display more morphological variability, possibly owing to the use of various bones left over from butchery.
Likewise, Pasveer and Bellwood (2004) report that small bipoints were recovered from Siti Nafisah on Halmahera, which again were made on small fragments of cortical bone and shaped using scraping and grinding. The authors comment that the bipoints from this site are more homogenous in their morphology than those at Golo Cave, but display very similar use wear, including polish on their surfaces. These same tiny bipoints were also identified at Daeo Cave and Uatamdi Rockshelter, though in much lower numbers (Table 2), along with larger unipoints which could be plant- or hide-working tools owing to their morphology and reported use wear traces (Pasveer and Bellwood, 2004). Most recently, Gaffney et al. (2025) described nine bone bi- or unipoints along with another five possible bone artefacts found in terminal Pleistocene through to mid-Holocene layers at Mololo Cave on Waigeo Island, just off the bird’s head of New Guinea. These artefacts are also similar in material, size, manufacture, and use wear to those described here for Taora, but appear in contexts without fish remains.
To the east, at Balof on New Ireland, Downie and White (1978) recovered five intact and 20 fragments of bone points in levels IV to VIII with an associated age between 7000 and 2500 BP. These artefacts are once again small bipoints (along with one unipoint) of similar size and manufacture to those at Taora, and two points are reported as appearing ‘to be unfinished, having been cut to shape but with the tips remaining unground’ (Downie and White, 1978: 777). The authors conclude that ‘it is likely that the points were spear or arrow barbs’ based on comparison to ethnographic specimens from the Bismarcks and Bougainville and note the use of the site as a hunting base.
On the other side of New Guinea, several bone and tooth artefacts are reported from the sites of Marea Ita 1 and Maivipi 1 in Orokolo Bay in the Papua Gulf (Basiaco et al., 2020). No measurements are given for these tool fragments, though one elongated point is interpreted as a possible awl, while three others are suggested to perhaps have functioned as projectile points. Examination of the provided photographs finds that these three latter pieces are likely mesial-proximal fragments from small bipoints such as those found at Taora and elsewhere in New Guinea. Such an interpretation may provide an explanation for the possible ochre resides seen on one example if colourants were utilised in hafting.
Further west, on the Aru Islands, the cave sites of Liang Lemdubu and Liang Nabulei Lisa preserved small collections of bone implements (Pasveer, 2005). Liang Nabulei Lisa preserved 15 bone artefacts, most of which date to roughly between 13,000 and 9000 BP (Pasveer, 2005). Liang Lemdubu had a larger assemblage of 37 artefacts, these dating between c.1830 ± 60 to >25,700 ± 460 but with the majority in spits dating to the Late-Holocene (Pasveer, 2005). Unlike the northern New Guinea sites, only one bipoint was among the 52 artefacts analysed by Pasveer from Liang Lemdubu and Liang Nabulei Lisa, though for Lemdubu Pasveer does report 21 short unipoint fragments that ‘are parts of broken uni- or bipoints’. It is unclear what size these points are, though from the included photograph of sectioned artefacts, at least one may be similar to the small Taora points. The remaining artefacts are similar to the plant- or hide-working implements, some being pointed (as the Taora examples) and others being spatulate (as seen at Golo Cave). Again, the use of cortical bone or shaft pieces of mammal bone was found to be used to make these artefacts. Most were ground into shape, with some being shaved/scraped prior to grinding. Interestingly, while Pasveer (2005) suggested that lightly burnt bone seems to have been deliberately selected for manufacturing bone implements on the Aru Islands, we didn’t find evidence for this approach at Taora.
Each of the studies cited above have provided an interpretation for both the small uni- and bipoints and larger unipoints. It is interesting to note that while the larger points appear universally to have been interpreted as plant-working tools in the main, the small (usually bipointed) pieces attracted a few different ideas. Initially, Pasveer and Bellwood (2004: 329) argued that the ‘the high manufacturing effort represented by each of these delicate and highly modified artefacts. . ..are likely to have relatively specialised and specific functions’, which they surmise were part of a composite tool, and owing to their similarity to recorded ethnographic items in the Indo-Pacific region, were either fish-gorges (Anell, 1955; Codrington, 1891; Massola, 1956) or multi-pronged fishing spears (McCarthy, 1940; Osmond, 1998). In her later analysis of the Kria and Toé Cave examples, however, Pasveer (2005) changes her mind about interpreting the small points as the tips of multi-pronged fishing spears. In this work, she instead suggests that they were used for drilling holes or in engraving activities, an interpretation supported by an ethnographic account of the Wola of the Southern Highlands (Sillitoe, 1988). There hafted bone points and animal incisors were noted in such activities. While we would agree with the interpretation of the utilised macropod teeth as drilling or engraving tools, with such uses being observed across the Australian continent (Langley, 2023 and references therein), we argue that the small bone uni- and bipoints were more likely the tips of multi-pronged spears. While bone points of this kind might be used, on occasion, for carving or drilling, generally these artefacts are not very effective in this kind of work—unlike teeth which are very effective.
It is worth reiterating that multi-pronged spears and arrows were not restricted to catching fish, though they are exceptionally well-suited to this function. As was cited above, such weapons are also known to be useful for arboreal and flighted prey, which goes towards explaining why such artefacts are found in near-coastal sites with abundant fish remains (Taora, Daeo Cave 2) and those firmly inland with little or no fish (Kria, Toé, Golo Cave, Siti Nafisah, Mololo). Furthermore, if people are moving around the landscape to take advantage of various resources and visit with neighbouring communities, they may take tools which are primarily used in one circumstance (i.e., fishing) into different contexts where they may find use (i.e., in capturing arboreal prey) but to a lesser degree.
From these data it appears that the small bipoints—identified as having tipped fishing and/or hunting multi-pronged weapons—are found in greater numbers across the northern half of New Guinea and nearby islands, while sites situated across the south have very few or none of these artefacts owing to their later antiquity. Whether this apparent geographical distribution continues to become apparent as excavations continue across the region is a matter of interest as the developing pattern may indicate a difference in Mid-to-Late-Holocene technologies.
Finally, Taora records human behaviour across the dramatic shift recorded around 6500 cal. BP previously noted as one of the most striking features of the archaeological record on the north coast of New Guinea. Swadling and Hide (2005) have suggested that sea level stabilisation created hitherto unprecedented rich marine and lacustrine environments adjacent to fertile land which led to new settlement foci, supported by a new subsistence strategy involving locally developed horticultural strategies and marine or estuarine resource use. Allen (2000) goes further, suggesting that there was a transition from a predominantly mobile hunter-gatherer lifestyle to a socioeconomic system based substantially on food production with greater dependence on spatially fixed resources, increasing sedentism and ultimately, a more spatially restricted and stronger sense of territoriality (e.g., Denham and Haberle, 2008). Certainly, Taora reflects an intense focus on the exploitation of marine resources which would support this theory, which is joined by the large quantities of fish and shellfish remains found at the nearby Lachitu cave (Gorecki et al., 1991; O’Connor et al., 2011) and Watinglo rockshelter (O’Connor et al., 2011). All three of these caves record a similar depositional hiatus between the late Middle and early Late-Holocene, adding to a growing sense of the widespread abandonment or sediment erosion in sites across northern New Guinea starting in the mid-Holocene and continuing until about 2000 cal. BP.
Conclusion
With excavation of mid-Holocene sites across New Guinea remaining sporadic and few in number compared to the more intensive work being undertaken on the Indonesian islands to the west, documenting the antiquity, diversity, and change in New Guinean toolkits remains in its infancy. This situation is particularly so for organic technologies, though some patterns are beginning to emerge in terms of the use of tiny flaked and scraped bipoints for tipping multi-pronged spears across the north. Further investigation of sites across this large island will determine how ‘real’ this apparent distribution is for reflecting past behaviour, but for now, we have the beginnings of understanding their role in a period of transition from mobile hunter-gatherer-fisher to, perhaps more territorial and focused exploitation of the coastline.
Supplemental Material
sj-xlsx-1-hol-10.1177_09596836261450782 – Supplemental material for Itty-bitty bone points from New Guinea: Bone artefacts and fishing between 6800 and 6000 cal BP at Taora
Supplemental material, sj-xlsx-1-hol-10.1177_09596836261450782 for Itty-bitty bone points from New Guinea: Bone artefacts and fishing between 6800 and 6000 cal BP at Taora by Michelle C. Langley, Loukas Koungoulos, Rintaro Ono and Sue O’Connor in The Holocene
Footnotes
Acknowledgements
The fieldwork in PNG was carried out under the auspices of the National Museum and Art Gallery (NM&aG) and the Sandaun Provincial Government. We would particularly like to acknowledge our gratitude to the late Baiva Ivuyo (NM&aG) who assisted with the excavation at Taora in 2004. Rose Singadan, Biology Department, UPNG gave us access to the faunal collections from Gorecki’s earlier excavations at Lachitu. The chiefs, councillors and communities of the villages of Fichin and Musu are thanked for granting permission and assisting with the excavation at Taora.
Author contributions
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Fieldwork and dating for the original Taora excavation were supported by an Australian Research Council Discovery Grant DP0451043. This research on the legacy collection was supported by The ARC Centre of Excellence in Australian Biodiversity and Heritage (CABAH) CE170100015.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.