
Abstract
The disappearance of Nesophontidae represents one of the few family-wide mammalian extinctions documented in recent history. Previous radiometric studies from Hispaniola suggested that this extinction occurred during European colonization, coinciding with significant landscape transformation and the introduction of invasive species, including the black rat (Rattus rattus). However, direct evidence for the temporal overlap between nesophontids and invasive rats has remained elusive. Here, we present three new direct AMS radiocarbon dates from a cave assemblage in southwestern Hispaniola (Greater Antilles), including the Last Appearance Date (LAD) of Nesophontes hypomicrus, an endemic small-bodied insectivorous mammal, and two post-Columbian dates for the invasive black rat (Rattus rattus). Our results narrow the temporal gap between the last occurrence of Nesophontes and the introduction of invasive rats, and are consistent with a brief regional temporal overlap during the early colonial period. When considered alongside existing radiometric evidence for black rat introduction, these findings are consistent with the hypothesis that the invasive species introduced following European colonization may have contributed to the final decline of Hispaniolan nesophontid populations.
Introduction
The West Indies have experienced exceptionally severe mammalian losses during the Holocene, with approximately 84% of native non-volant mammal species becoming extinct (Cooke et al., 2017; Dávalos and Turvey, 2012; MacPhee, 2009; Orihuela et al., 2020a, 2020b). This extinction hotspot therefore provides a unique framework for studying the dynamics and drivers of extinction processes in insular faunas (Myers et al., 2000). However, the scarcity of stratified subfossil assemblages, combined with poor collagen preservation typical of tropical environments, has hindered precise determination of extinction timing for most endemic West Indian mammals (Cooke et al., 2017; Morgan and Woods, 1986; Orihuela et al., 2020a, 2020b; Woods et al., 2020).
Among the recently extinct Caribbean mammals are the Nesophontidae, an endemic family of small (c. 10–125 g) plesiomorphic eulipotyphlan insectivores comprising eight nominal species that formerly occurred on Cuba, Puerto Rico, Hispaniola, Grand Cayman and Cayman Brac (e.g. Miller, 1929; Morgan, 1994; Orihuela et al., 2020a, 2020b; Orihuela León, 2023; Turvey and Fritz, 2011; Woods et al., 2020). Three nesophontid species have been described from Hispaniola, the second largest Caribbean island: Nesophontes paramicrus (39–47 g), N. hypomicrus (21–24 g), and N. zamicrus (~ 10 g), differentiated primarily by body size (Miller, 1929). Recent morphometric and genetic studies, however, suggest that species boundaries among Hispaniolan Nesophontes—particularly between N. hypomicrus and N. zamicrus—may be less distinct than previously assumed, and potential conspecificity has been proposed (Buckley et al., 2020; Orihuela León, 2023; Woods et al., 2020). All three species have been reported to co-occur in recent subfossil deposits across Hispaniola (MacPhee et al., 1999), suggesting a singular case of insular adaptive radiation driven by niche partitioning among size classes (Orihuela et al., 2020a).
The factor(s) responsible for nesophontid extinction remain a subject of considerable debate. Natural or anthropogenic environmental change, introduced predators, competitors, diseases, or combinations thereof have all been proposed as possible causes (MacPhee et al., 1999; Morgan and Woods, 1986). However, landscape transformation and the introduction of invasive species, such as the black rat (Rattus rattus), following European colonization are widely regarded as among the most plausible drivers of this lineage’s extinction (Borroto-Páez and Mancina, 2017; Guiry et al., 2024; MacPhee et al., 1999). Notably, this extinction event represents one of the few family-wide mammalian extinctions documented in recent history (Turvey et al., 2007; Turvey and Fritz, 2011).
Radiocarbon dating has been crucial for testing extinction hypotheses by establishing temporal frameworks. Last Appearance Dates (LADs), representing the most recently dated specimens, for Nesophontes on Puerto Rico, Hispaniola, the Cayman Islands, and Cuba generally predate European colonization by several centuries (Brace et al., 2016; MacPhee et al., 1999; Morgan et al., 2019; Orihuela et al., 2020a, 2020b; Woods et al., 2020), whereas First Appearance Dates (FADs) for black rats on Hispaniola coincide with their introduction around the time of initial European contact (Cooke and Erin Crowley, 2022; MacPhee et al., 1999). This apparent temporal offset of roughly 300 years has hindered efforts to directly link rat arrival with nesophontid extinction.
While such gaps are expected due to inherent biases of the fossil record (the “Signor-Lipps effect”; Signor and Lipps, 1982), they are further exacerbated for microfauna (Semken et al., 2010). The minute skeletal elements of small animals yield very limited collagen, often making it difficult to obtain enough material for 14C dating. This in turn limits the number of datable specimens. Recent studies, however, have shown that these chronological gaps can be narrowed through targeted sampling of late-surviving assemblages (e.g. Bover et al., 2016) and advances in AMS radiocarbon dating techniques applicable to small collagen samples (Cersoy et al., 2017). Thus, obtaining more direct radiocarbon dates has become both feasible and essential for refining the chronology of taxon persistence at specific localities and for overcoming longstanding preservation and sampling biases in the Caribbean (Cooke et al., 2017).
Here, we present three new direct AMS radiocarbon dates for Nesophontes spp. and Rattus rattus from Hispaniola, refining extinction timing and providing a chronological framework to evaluate hypotheses concerning the role of biological invasion in this process.
Material and methods
Fieldwork was conducted in July 1996 as part of a multidisciplinary campaign jointly led by J.A. Alcover and J.A. Ottenwalder within the framework of the Spanish Ministry of Education and Science’s Ibero-American Cooperation Program (Trias et al., 1997). The survey targeted the karstic littoral region surrounding Laguna de Oviedo, within Jaragua National Park (Pedernales Province, southwestern Dominican Republic). A total of 12 caves were explored on the western shoreline of the lagoon, with vertebrate and crustacean remains documented in at least seven of them. Among these, Pozimán Cadena was identified as a site of particular interest due to a distinctive pellet-derived accumulation of small vertebrate remains, which included both endemic and introduced taxa (Figure 1).
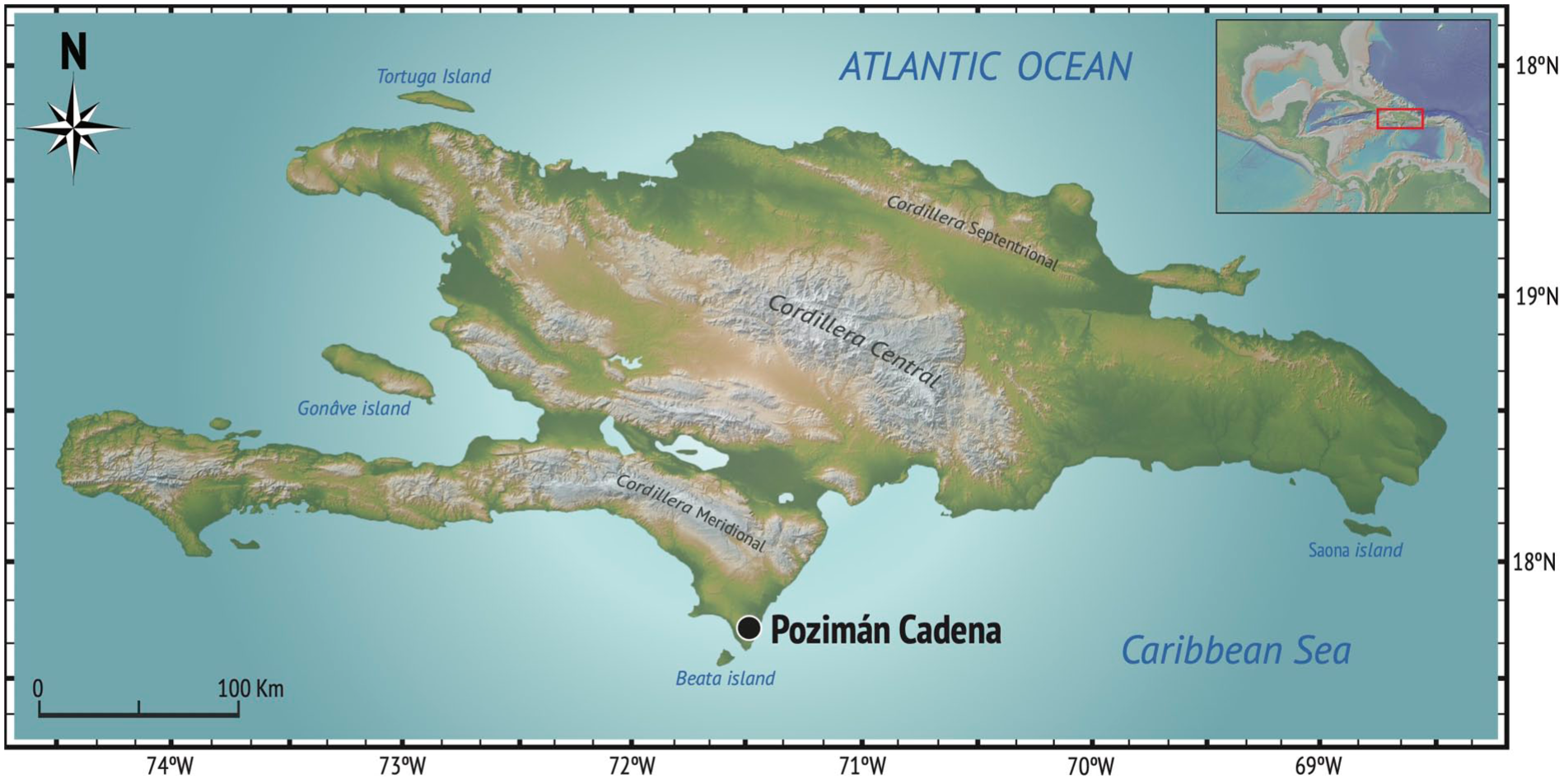
Geographic location of the Pozimán Cadena site on Hispaniola.
Pozimán Cadena is a shallow collapsed sinkhole (cenote) located near the southwestern shore of Laguna de Oviedo (17.744975°N, 71.402884°W; WGS84). The region is characterized by a tabular Oligo–Miocene limestone platform with intense hydrologic erosion (León, 1989), further described by Jaume and Wagner (1998) regarding anchialine cave systems in the area. The cenote, situated at <80 m a.s.l., connects with the adjacent hypersaline lagoon and contains brackish water in its lower reaches.
The upper zone of the cave contains a well-preserved accumulation of small vertebrate bones in superficial dry sediments. The assemblage likely originated from pellet deposition by the ashy-faced barn owl (Tyto glaucops) and includes abundant remains of Nesophontes spp., Rattus sp., and several bat taxa (Artibeus jamaicensis, Brachyphylla nana, Stenoderma sp., cf. Macrotus waterhousei), as well as unidentified avian and reptilian taxa. Although no clear stratification was observed, the Rattus remains were located in the uppermost part of the deposit, slightly offset vertically from the bulk of the Nesophontes material. Representative cranial and mandibular material used for taxonomic assessment is illustrated in Supplemental Figure S1.
Identification of Rattus remains was based on osteological criteria traditionally applied in archeological and zooarchaeological contexts, including mandibular and cranial morphology and metric traits (e.g. Armitage et al., 1984; Cuenca-Bescós and Morcillo-Amo, 2022; Miller, 1912; Wolff et al., 1980). While isolated mandibles alone may not always allow unequivocal distinction between R. rattus and R. norvegicus, the assemblage includes both mandibular and cranial elements, none of which showed characters attributable to R. norvegicus; the combined evidence from dental morphology, cranial characters, and osteometric ranges consistently supports attribution to R. rattus. This identification is further reinforced by the historical and biogeographic framework of rat introductions in the West Indies, where R. rattus represents the earliest invasive species associated with European colonization, whereas R. norvegicus is generally regarded as a later arrival, typically from the 18th century onward (Borroto-Páez and Woods, 2012).
Taxonomic identification of Nesophontes specimens was based on morphological and morphometric comparison with published reference data from Hispaniola (Morgan et al., 2019; Orihuela León, 2023). Four diagnostic measurements were recorded for each mandible (i.e. mandibular length, molar crown length from M1–M3, depth of the horizontal ramus, and height of the coronoid process) and compared to established metric ranges for N. hypomicrus, N. paramicrus, and N. zamicrus to confirm species-level attribution (Table 1).
Morphometric characters (minimum, maximum, and mean) of the dated Nesophontes spp. hemimandibles from Pozimán Cadena compared with the published metric ranges of the three Nesophontes species identified on Hispaniola (N. paramicrus, N. hypomicrus, N. zamicrus).
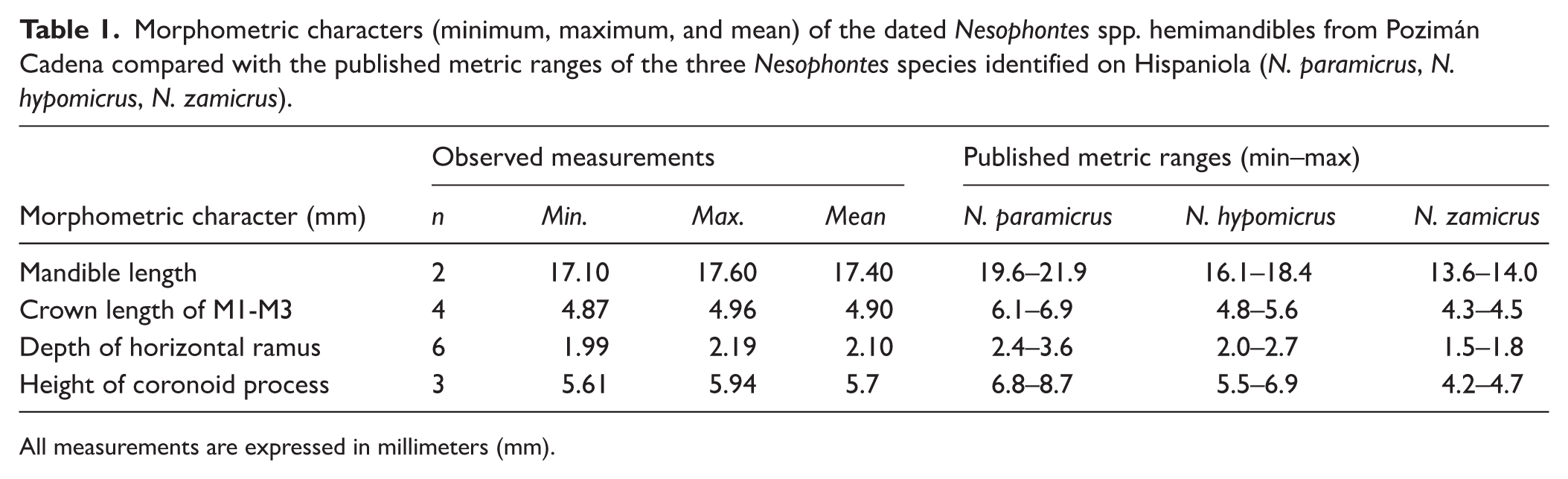
All measurements are expressed in millimeters (mm).
From this deposit, we selected specimens for radiocarbon dating based on unambiguous taxonomic identification and good macroscopic preservation. Specimens were pooled by taxon from the same assemblage context because individual skeletal elements of Nesophontes and Rattus yield insufficient bone mass for reliable AMS dating under the KIK-IRPA protocol (minimum ~0.4–0.5 g; Wojcieszak et al., 2020). Importantly, any time-averaging from pooling biases dates toward older values, making our LAD for Nesophontes a conservative minimum bound on last occurrence. Direct AMS 14C dating was performed at the Radiocarbon Laboratory of the Royal Institute for Cultural Heritage (KIK-IRPA, Brussels), following the modified Longin collagen extraction protocol (Longin, 1971), described by Wojcieszak et al. (2020), which involves mechanical surface cleaning, demineralization in HCl, removal of humic acids with NaOH, collagen solubilization at pH 3 and 90°C, and freeze-drying of the extracted gelatin. Conventional radiocarbon ages were calibrated using OxCal v4.4 (Bronk Ramsey, 2009) with the IntCal20 calibration curve (Reimer et al., 2020). Calibrated age ranges are reported at 68.3% (1σ) and 95.4% probability (2σ). Stable isotope ratios (δ13C and δ15N) were measured in duplicate by isotope ratio mass spectrometry (IRMS) on a Thermo Flash EA/HT elemental analyzer coupled to a Thermo DeltaV Advantage IRMS via ConfloIV interface at the Department of Earth and Environmental Sciences, KU Leuven (Belgium), to assess collagen integrity and evaluate potential dietary effects on 14C ages (Lanting and van der Plicht, 1998; van Klinken, 1999). Collagen quality indicators (%C, %N, atomic C:N ratio, and collagen yield) were determined on the extracted collagen of each pooled sample and are reported in Table 2; all values fall within accepted thresholds for reliable radiocarbon and isotopic analysis (van Klinken, 1999).
New radiocarbon dates from Pozimán Cadena (Parque Nacional de Jaragua, Oviedo).
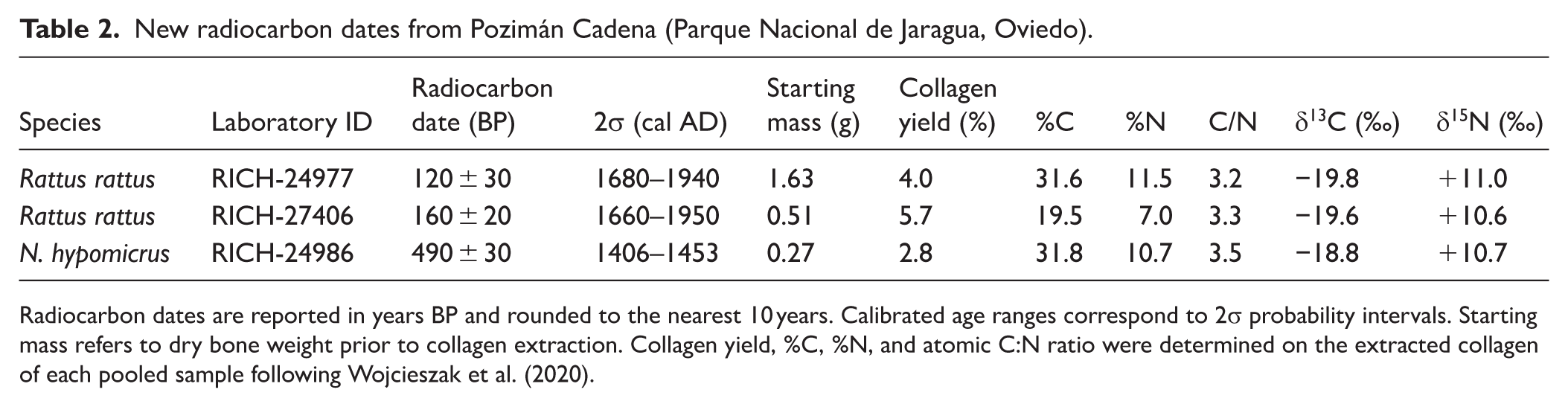
Radiocarbon dates are reported in years BP and rounded to the nearest 10 years. Calibrated age ranges correspond to 2σ probability intervals. Starting mass refers to dry bone weight prior to collagen extraction. Collagen yield, %C, %N, and atomic C:N ratio were determined on the extracted collagen of each pooled sample following Wojcieszak et al. (2020).
Results
All recorded measurements fall within the diagnostic range of N. hypomicrus and outside the ranges of both N. paramicrus and N. zamicrus as defined by Morgan et al. (2019), confirming the taxonomic attribution of the dated specimens. Specifically, mandible length ranged from 17.10 to 17.6 mm (n = 2), consistent with the N. hypomicrus range (16.1–18.4 mm) and clearly below that of N. paramicrus (19.6–21.9 mm). Similarly, molar crown length (M1–M3) ranged from 4.87 to 4.96 mm (n = 4), aligning with N. hypomicrus (4.8–5.6 mm) and exceeding that of N. zamicrus (4.3–4.5 mm). The horizontal ramus depth measured 1.99–2.19 mm (n = 6), within the N. hypomicrus range (2.0–2.7 mm) and above that of N. zamicrus (1.5–1.8 mm). Finally, coronoid process height ranged from 5.61 to 5.94 mm (n = 3), also matching that of N. hypomicrus (5.5–6.9 mm).
A total of three sets of bone samples from the Pozimán Cadena assemblage were successfully radiocarbon dated (Table 2; Figure 2). In all cases, dates were obtained from taxonomically homogeneous sets of elements pooled to ensure sufficient collagen yield and to provide conservative estimates for the timing of taxon presence.
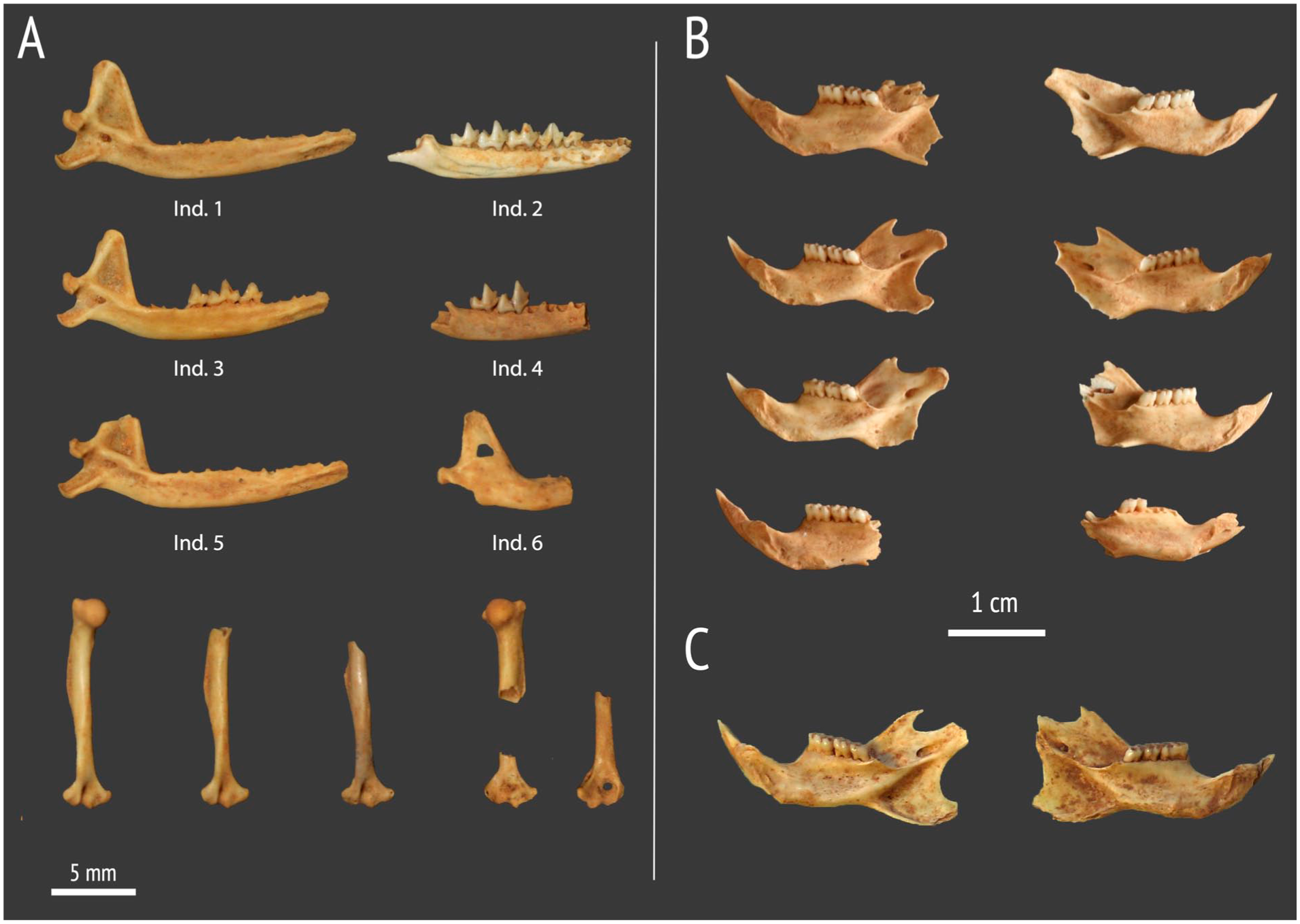
Material from Pozimán Cadena selected for radiocarbon dating: (a) sample RICH-24986, comprising six fragments of hemimandibles and six fragments of humeri of Nesophontes hypomicrus, (b) sample RICH-24977, eight partial hemimandibles of R. rattus, and (c) sample RICH-27,406, two hemimandibles (one from each side) of R. rattus.
The first sample (RICH-24986; Figure 2a) was composed of Nesophontes hypomicrus mandibles and humeri. The humeral fragments were attributed to N. hypomicrus on the basis of their overall dimensions, which exclude the substantially larger N. paramicrus and are consistent with the mandibular material from the same pool. This sample yielded a conventional radiocarbon age of 486 ± 29 BP, corresponding to a calibrated 2σ range of 1406–1453 cal AD, and represents the most recent directly dated occurrence for this extinct insectivore, slightly refining the known LAD of Hispaniolan nesophontids.
The other two samples consisted of grouped R. rattus mandibles: one (RICH-24977; Figure 2b), pooling eight elements, produced an age of 120 ± 28 BP (1680–1940 cal AD), while the other (RICH-27406; Figure 2c), comprising two mandibles, yielded 160 ± 22 BP (1660–1950 cal AD). These results align with a post-Columbian introduction of black rats following European contact (MacPhee et al., 1999). The internal consistency between the two R. rattus samples and their chronological separation from the Nesophontes date indicate a later depositional phase for the rat remains. Stable isotope analysis revealed δ13C values of –19.8‰ and –19.6‰ for R. rattus, and –18.8‰ for N. hypomicrus, consistent with predominantly terrestrial carbon sources. Nitrogen isotope values (δ15N) ranged from +10.6‰ to +11.0‰ for R. rattus, and +10.7‰ for N. hypomicrus. All dated samples exhibited atomic C:N ratios between 3.2 and 3.5, indicating good collagen preservation.
Discussion
Recent applications of direct AMS radiocarbon dating to microfaunal remains have improved the chronological resolution of late Holocene extinctions in the Caribbean, although significant uncertainties persist. The timing of Nesophontes extinction relative to European arrival and the introduction of invasive species has been particularly difficult to resolve, owing to the scarcity of well-stratified assemblages and the inherent challenges of dating microfaunal remains in tropical contexts (e.g. Borroto-Páez and Mancina, 2017; MacPhee et al., 1999; Morgan and Woods, 1986). While several endemic Caribbean rodents are now directly dated into the late-Holocene or even post-Columbian period (Cooke and Erin Crowley, 2022; Viñola-López et al., 2022), direct radiocarbon determinations for Hispaniolan Nesophontes have remained scarce (e.g. Brace et al., 2016); existing LAD compilations (Orihuela et al., 2020a, 2020b) document the broader Greater Antillean chronology but highlight the difficulty of constraining extinction timing from time-averaged cave deposits, where dated specimens constrain minimum persistence rather than actual extinction timing. It was unclear whether these small insectivores survived until European contact or had disappeared earlier, leaving a temporal gap between their LADs and the FADs of commensal Old-World rats on Hispaniola (MacPhee et al., 1999). Our new results narrow this gap by documenting a later-than-previously-reported persistence of Nesophontes, and are consistent with a possible regional-scale temporal overlap with the earliest R. rattus invasion.
Previous radiocarbon studies dated two of the three known Hispaniolan Nesophontes species (N. paramicrus and N. hypomicrus) to the centuries immediately preceding European colonization. For instance, a N. paramicrus from Constanza yielded a calibrated age of 1294–1425 cal AD (Beta-106566; MacPhee et al., 1999), while specimens from Cueva Jung (Beta-104812; MacPhee et al., 1999) and Cueva de Bosque Húmedo (OxA-19721; Brace et al., 2016) produced similar 13th–14th century ages. No specimen of the smallest species, N. zamicrus, has yet been directly dated. Collectively, these results indicate LADs within roughly 500–700 14C years BP (c., 13th–15th centuries AD), broadly preceding the island’s first Spanish contact in 1492.
Our new AMS radiocarbon determination for N. hypomicrus from Pozimán Cadena refines and slightly extends this survival window. The pooled Nesophontes sample yielded a conventional age of 486 ± 29 BP, calibrating to 1406–1453 cal AD (95.4% probability). This mid-15th-century LAD currently represents the most recent directly dated occurrence of any West Indian nesophontid, while acknowledging that the Signor-Lipps effect (Signor and Lipps, 1982) implies that this specimen is unlikely to represent the final individual of the species (Figure 3).
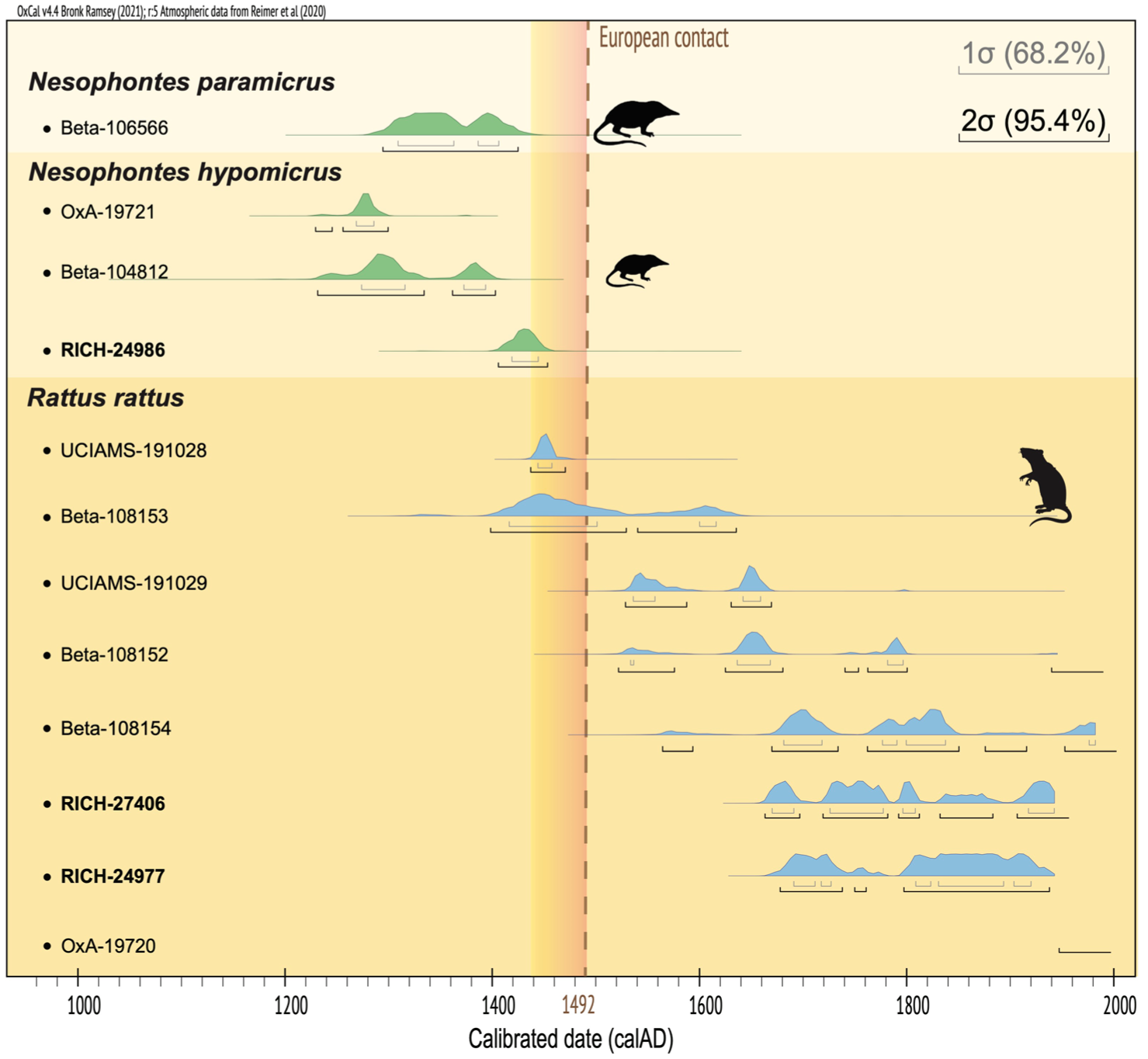
Chronology of the extinction of endemic West Indian nesophontids (Nesophontes spp.) and the introduction of black rats (Rattus rattus) on Hispaniola.
This result aligns with associated chronologies from archeological contexts. Notably, at the El Flaco site in northwestern Dominican Republic, N. paramicrus remains were recovered from late pre-Columbian deposits dated to 1328–1490 cal AD (Shev et al., 2023). Although the Nesophontes bones at El Flaco were not directly dated, their stratigraphic association with ~15th-century layers (Unit 69) is consistent with, though not direct evidence of, survival until near the time of first European contact. In pellet-derived and cave assemblages, spatial or stratigraphic association alone is insufficient to establish contemporaneity, as time-averaging and mixing are common even within single beds or concentrations (Orihuela et al., 2020a; Semken et al., 2010). Together, the El Flaco evidence and our new date from Pozimán Cadena are consistent with the persistence of at least two Hispaniolan Nesophontes species well into the 15th century AD. This significantly narrows the chronological gap between nesophontid extinction and the onset of European colonization. While a precise terminal date remains unknown (no specimens are yet found in clearly post-1492 contexts), the data imply that Nesophontes persisted on Hispaniola up to the brink of European arrival, if not overlapping briefly with the earliest colonial period. Importantly, the radiocarbon dates presented here constrain population-level chronologies rather than individual extinction events and therefore do not demonstrate direct causal relationships between invasive species and nesophontid extinction, but instead provide a refined temporal framework within which competing extinction hypotheses can be evaluated.
In contrast, radiocarbon evidence for invasive black rats on Hispaniola points to an introduction coincident with, or shortly after, European contact. Both of our R. rattus samples from Pozimán Cadena produced post-Columbian ages (~1660–1950 cal AD), suggesting that, despite their spatial association with the endemic fauna, they belong to a later depositional phase. This implies that the assemblage likely represents multiple periods of accumulation; a pattern well documented in karst cave contexts, where faunal remains often reflect time-averaged deposits spanning centuries (e.g. Cooke and Erin Crowley, 2022; MacPhee et al., 1999; Orihuela et al., 2020a, 2020b; Viñola-López et al., 2024). These R. rattus dates are entirely consistent with the known post-1492 dispersal of black rats by Europeans, though they do not, in themselves, clarify the initial timing of invasion.
Previous studies have provided direct radiocarbon dates for Rattus rattus on the island. MacPhee et al. (1999) reported three AMS dates on R. rattus femora from a Dominican cave (Monte Culo de Maco), the earliest of which (Beta-108153, 440 ± 60 BP) calibrates to 1398–1635 cal AD, with the highest probability (72.6%) between 1398 and 1529 cal AD. The other two samples (Beta-108152 and Beta-108154) were clearly post-Columbian, with 95.4% ranges spanning 1522–1940 cal AD and 1526–1950 cal AD, respectively, their main probabilities clustering between the 17th and 19th centuries.
A more recent study reported two additional AMS radiocarbon dates for Rattus sp. from Hispaniola (Cooke and Erin Crowley, 2022). One falls within a calibrated 2σ range of 1521–1662 cal AD (UCIAMS-191029; 280 ± 20 BP), while the other produced an unexpectedly early range of 1435–1468 cal AD (UCIAMS-191028; 435 ± 15 BP), potentially predating the arrival of Europeans in 1492. This latter result is best regarded as a conservative terminus post quem for rat arrival on Hispaniola rather than evidence of pre-Columbian presence, consistent with the authors’ own interpretation. The absence of any published anatomical description, illustration, or stable isotope values (δ13C) for this specimen limits the evaluation of possible biases. Radiocarbon calibration during the colonial interval is complicated by the multi-intercept structure of the IntCal20 calibration curve (Reimer et al., 2020), which can produce ambiguous or apparently early age ranges for samples of this period. Taken together, the available radiocarbon evidence is consistent with a post-1492 introduction of black rats, in line with historical and biogeographic expectations.
In this context, stable isotope data (although based on a limited number of samples) are used not as evidence of ecological interaction per se, but as an independent line of evidence to assess potential dietary biases affecting radiocarbon chronologies. Carbon isotope values (δ13C) for both Nesophontes hypomicrus and Rattus rattus fall within the range expected for terrestrial C3-based food webs, indicating that marine-derived carbon is unlikely to have influenced the reported ages. In the case of Nesophontes, a strict insectivore, a marine dietary contribution can be confidently excluded. Although some commensal populations of R. rattus may occasionally consume aquatic resources (Puckett et al., 2020), the isotopic values reported here fall well within the terrestrial range, suggesting that marine reservoir effects did not significantly bias the radiocarbon results. Nitrogen isotope values (δ15N) indicate protein-rich diets for both taxa; however, given the limited sample sizes (n = 1–2), these data do not allow robust inference of dietary overlap, trophic competition, or direct ecological interactions.
Taken together, the available evidence is consistent with a scenario in which the European colonization introduced a suite of novel pressures that exceeded the resilience of nesophontid populations on Hispaniola. While the precise mechanism remains uncertain, the introduction of black rats likely constituted one of several important contributing factors. It is important to note that European settlement brought multiple stressors to West Indian ecosystems, and Nesophontes extinction cannot be attributed to a single factor with absolute certainty. Widespread habitat alteration (e.g. deforestation, agriculture) and other introduced biota (cats, dogs, mongooses, commensal house mice, etc.) likely contributed to faunal turnover in complex ways (Borroto-Páez and Mancina, 2011; Morgan and Woods, 1986). Nonetheless, black rats emerge as a particularly plausible contributor to the decline of nesophontid populations. Rattus rattus is among the most pervasive and ecologically destructive invasive mammals, implicated in numerous insular extinctions via predation, competition, and disease transmission (e.g. McCreless et al., 2016; Smith and Banks, 2014). On many oceanic islands, invasive rats have devastated native bird colonies through egg and chick predation and have decimated lizard and small mammal populations through direct predation and competition (Shiels et al., 2014; Siracusa et al., 2012). Global meta-analyses identify rats as leading contributors to island extinctions (Jones et al., 2008, 2016). Caribbean ecosystems were no exception: indigenous land animals evolved without evolutionary exposure to introduced, ground-dwelling mammalian predators, rendering them naïve and highly vulnerable upon rat arrival (Morgan and Woods, 1986; Orihuela et al., 2020b; Viñola-López et al., 2022, 2024). For Nesophontes (i.e. small, ground-dwelling insectivores) rat predation would likely have represented a direct threat, while competition for invertebrate prey or shelter could also have reduced nesophontid viability. Moreover, invasive rats are known carriers of pathogens and parasites that could have been novel to nesophontids, and disease-mediated impacts cannot be discounted. Such effects have caused extinctions elsewhere, notably the loss of Christmas Island’s native rats due to a trypanosome introduced by R. rattus fleas (Wyatt et al., 2008). Together, these interacting pressures associated with European colonization may have exceeded the adaptive capacity of nesophontid populations, ultimately contributing to their extinction on Hispaniola. These mechanisms are not directly demonstrated by the present dataset, but are well documented in comparable insular systems and provide a plausible ecological context for interpreting the observed chronological patterns.
Conclusions
Our findings highlight a close temporal proximity between the final persistence of Nesophontes on Hispaniola and the onset of European-mediated ecological change. The new LAD of ~1400–1450 AD for Nesophontes hypomicrus, when considered alongside the earliest robust evidence for the introduction of black rats shortly after 1492, indicates that nesophontid extinction occurred at, or immediately prior to, the threshold of European arrival.
At the same time, the available radiocarbon evidence remains limited. No nesophontid remains have yet been directly dated to the post-contact period, and the precise timing of black rat arrival on Hispaniola still relies on a small number of dates that warrant further confirmation. Consequently, the dates presented here constrain the chronology of taxon presence at Pozimán Cadena rather than the precise timing of species-level extinction, and they do not, by themselves, establish a direct cause-and-effect relationship.
Although it does not provide definitive proof, this study narrows the temporal window within which competing extinction scenarios must be evaluated. Within this framework, invasive species introduced during European colonization—particularly black rats—emerge as especially plausible contributors to the rapid decline and eventual loss of nesophontid populations, acting in concert with other anthropogenic stressors such as habitat transformation and broader ecological disturbance.
More broadly, this work underscores the importance of integrating high-resolution paleoecological chronologies with invasion biology in insular systems. By tightening the temporal relationship between endemic persistence and invasive species arrival, our results contribute to a growing body of evidence showing that many island extinctions were not slow, attritional processes, but abrupt ecological transitions closely tied to human-mediated introductions during the Late-Holocene.
Supplemental Material
sj-docx-1-hol-10.1177_09596836261458223 – Supplemental material for New direct radiocarbon dates constrain the timing of Nesophontes spp. : Extinction and the arrival of Rattus rattus on Hispaniola (Greater Antilles)
Supplemental material, sj-docx-1-hol-10.1177_09596836261458223 for New direct radiocarbon dates constrain the timing of Nesophontes spp. : Extinction and the arrival of Rattus rattus on Hispaniola (Greater Antilles) by Alejandro Valenzuela, Enric Torres-Roig and Josep Antoni Alcover in The Holocene
Footnotes
Acknowledgements
Fieldwork and initial specimen recovery were carried out within the framework of the Ibero-American Cooperation Program of the Spanish Ministry of Education and Science and the project ICEX 472/95RD “Zonas cársticas insulares mediterráneas y caribeñas: aspectos espeleológicos, paleontológicos y carcinológicos”, led by J.A. Alcover and J.A. Ottenwalder. . We thank Dr. José Alberto Ottenwalder and Bienvenido Martínez for their logistical support during the fieldwork at the Laguna de Oviedo caves, and Juan José Enseñat (SEB, IMEDEA) and Damià Jaume (IMEDEA) for their assistance in locating and contextualising the Pozimán Cadena site. We are also grateful to J. Orihuela, S. Cooke and K. Hensley for helpful discussions and constructive review comments that substantially improved this manuscript. A.V. was supported by a Ramón y Cajal contract (RYC2022-037791-I) from the Spanish Ministry of Science and Innovation (MICIU/AEI/10.13039/501100011033) and E.T.R is financed by a postdoctoral grant (Programa Margalida Comas) from the Conselleria d’Economia, Hisenda i Innovació of the Govern de les Illes Balears (PD-042-2023).
Author contributions
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: A.V. was supported by a Ramón y Cajal contract (RYC2022-037791-I) from the Spanish Ministry of Science and Innovation. E.T.R. is financed by a postdoctoral grant (Programa Margalida Comas) from the Conselleria d’Economia, Hisenda i Innovació of the Govern de les Illes Balears (PD-042-2023). Fieldwork and initial specimen recovery were supported by the Ibero-American Cooperation Program of the Spanish Ministry of Education and Science and by the project ICEX 472/95RD “Zonas cársticas insulares mediterráneas y caribeñas: aspectos espeleológicos, paleontológicos y carcinológicos,” led by J.A. Alcover and J.A. Ottenwalder.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
All data supporting this study are included within the article. No additional datasets were generated or analyzed.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.