
Abstract
Our ability to remember past events requires efficient encoding and successful “re-collection” of event features, such as the time, place, people, thoughts, and feelings associated with a past experience. Neuroimaging studies have demonstrated that episodic memory depends on specific brain mechanisms within the prefrontal cortex, medial temporal lobe, and ventral posterior parietal cortex that interact with the cortical sites that store event features. In this article, I present an analysis of the neural correlates of episodic memory, along with a theoretical framework, Cortical Binding of Relational Activity (CoBRA), that integrates the dynamic links between efficient encoding and successful retrieval of episodic memory.
What did you eat for dinner last night? If you were able to retrieve the information (paella, in my case), it is likely that you also remembered other features, such as the time, place, people, sights, and sounds surrounding the event. Recollections of such episodic (i.e., autobiographical) memories have been likened to mental time travel, as they allow us to reenact or replay past events in our minds (Tulving, 2002). Such experiences connect us to our past—both personally and culturally—and, as such, have been celebrated as an essential part of what makes us human.
Episodic Encoding
To remember a past event, it is of course necessary to have registered and stored the experience in the first place. Thus, at the time of an event (last night’s dinner, in our example), the sensations, thoughts, and feelings that were encountered had to be encoded (i.e., processed and linked together) as an encapsulated set of event features. The cortical representation of these features—the various sights, sounds, ideas, and emotions associated with the event—is presumed to be distributed throughout the brain and held together on-line by an active working memory process. At any given moment, we cannot hold every sensation or thought in mind, and therefore we need to select, filter, and maintain certain features while dismissing others. Studies of brain-injured patients and functional neuroimaging analyses have shown that the prefrontal cortex (PFC) acts as an executive-control mechanism that supervises what is held in working memory (Miller & Cohen, 2001; Shimamura, 2008).
The landmark neurological case of Henry Molaison (Corkin, 2013; Scoville & Milner, 1957) highlighted the critical nature of the medial temporal lobe (MTL) in the storage of episodic memories. As an attempt to relieve intractable epileptic seizures, Molaison underwent an experimental operation that included bilateral removal of the anterior MTL, which included most of the entorhinal cortex and anterior hippocampus. 1 The operation mitigated Molaison’s seizures, but it left him with a debilitating impairment in his ability to remember events that occurred after his surgery. For example, after eating lunch, he could not remember what he had eaten or even if he had eaten at all. Like other MTL patients, he exhibited intact executive-control processes as well as normal language and social skills. MTL patients’ primary deficit is in storing new experiences, as a notable feature of MTL amnesia is relatively spared memory for personal information and public events that occurred before the onset of amnesia (Squire et al., 2010). Controversy still exists, however, over the time course of MTL contributions to episodic retrieval, as amnesic cases vary considerably in the degree to which retrieval of remote past memories is spared (Squire & Wixted, 2011; Winocur & Moscovitch, 2011). Neuroimaging studies have confirmed and clarified the important role of the MTL in memory formation. In particular, increased MTL activity during encoding is a strong predictor of subsequent memory performance (Paller & Wagner, 2002; Ranganath, 2010). Most theoretical accounts of MTL function point to its role in establishing long-lasting links that connect event features active during encoding (Cohen & Eichenbaum, 1993; Davachi, 2006; Ranganath, 2010; Shimamura, 2010; Squire & Wixted, 2011; Winocur & Moscovitch, 2011).
Taken together, a general processing framework can be developed that describes how the PFC, the MTL, and cortical regions that process and store event features interact during encoding to establish long-lasting episodic memories. Initially, bottom-up sensory processes, such as visual processing along the dorsal and ventral paths, are activated in response to sensory signals. PFC executive-control processes select, maintain, and update information in working memory, which comprises the ensemble of sensory, conceptual, and emotional features that make up an episodic memory. The MTL binds coactive features stored in disparate neocortical regions, thereby linking them as an encapsulated memory. Binding is established by way of a rapid associative process known to occur in MTL neurons (Lynch, Rex, & Gall, 2007). In computational terms, this process facilitates pattern separation, whereby relational bindings help to make each episodic memory distinct or separable from others (Norman & O’Reilly, 2003; Yassa & Stark, 2011). After initial encoding, reactivation or replay acts to consolidate or solidify episodic memories as strongly connected representations. Because MTL patients can retrieve personal memories of their past, it has been argued that episodic memories are ultimately established as cortical-cortical bindings and thus without the need of the MTL links, though controversial questions exist about the nature of such memories, such as whether they are qualitatively different from the kind of recollections we consider as “mental time travel” (see Nadel & Moscovitch, 1997; Shimamura, 2010; Squire & Wixted, 2011; Winocur & Moscovitch, 2011). In any event, such neocortical bindings are presumed to take longer to establish but, when formed, create long-lasting and well-integrated memory ensembles.
Episodic Retrieval
Retrieving an episodic memory often begins with a simple request, such as a friend asking “What did you do last night?” If memory serves, such a cue will evoke associative links to people, events, and experiences pertinent to the request. You may have first tried to search around a specific time period or activity (e.g., dinner) and from there come up with features surrounding the event. In computational terms, the process of “re-collecting” an ensemble of event features through relational bindings is called pattern completion (Norman & O’Reilly, 2003). In vernacular terms, we describe this process as reminiscing, which does have the quality of mentally traveling back in time (Tulving, 2002).
Neuroimaging studies have implicated a broad network of brain regions that is activated during episodic retrieval. Retrieval typically starts with PFC executive-control functions’ facilitating the search through memory and activating pertinent event features (Cabeza, Locantore, & Anderson, 2003; Wagner, 2002). The MTL contributes by activating event features through relational bindings (i.e., pattern completion; Ranganath, 2010; Shimamura, 2010). A rather surprising finding is that the ventral posterior parietal cortex (vPPC) is particularly involved during episodic retrieval. On recognition memory tests, this region is more active when individuals correctly identify items as “old” (hits) compared with when they correctly identify items as “new” (correct rejections, or CRs). This increased activation in response to hits relative to CRs is termed the successful-retrieval effect, and the vPPC is now recognized as one of the most active regions associated with the phenomenon (for a review, see Cabeza, Ciaramelli, & Moscovitch, 2012; Rugg & Vilberg, 2013; Shimamura, 2011; Uncapher, Gordon, & Wagner, 2014). The vPPC successful-retrieval effect has been observed using a variety of stimuli (pictures, faces, words, natural sounds, music clips) and tested with a variety of measures (recognition, cued recall, feeling of knowing, implicit memory). False recognitions, such as false alarms on recognition tests (identifying new items as “old”), also activate the vPPC to some degree, which suggests that this region signals subjective feelings of remembering, even when such feelings are invalid (Wheeler & Buckner, 2003).
The unexpected importance of the vPPC to episodic retrieval is attributed in part to the fact that lesions in this region do not produce severe impairment in memory retrieval. Patients with vPPC lesions exhibit a variety of other cognitive disorders, including deficits in visuospatial attention, sensorimotor integration, and verbal short-term memory (see Baldo & Dronkers, 2006; Pause, Kunesch, Binkofski, & Freund, 1989; Warrington & Shallice, 1969). In recent studies, however, patients with bilateral vPPC lesions have been found to exhibit impoverished autobiographical memories and to be less confident in their retrievals (Berryhill, Phuong, Picasso, Cabeza, & Olson, 2007; Hower, Wixted, Berryhill, & Olson, 2014; Simons, Peers, Mazuz, Berryhill, & Olson, 2010). Anatomical studies and multivariate analyses of fMRI data have shown that the vPPC is widely connected to many cortical regions, including the MTL, the PFC, and sensory regions, making it a useful hub for connecting multimodal features (Anderson, 1987; Jones & Powell, 1970; Seghier, 2013; Sestieri, Corbetta, Romani, & Shulman, 2011).
Several theories have been proposed that describe vPPC processes in terms of activating, maintaining, consolidating, or accumulating event features for the purpose of retrieving an encapsulated episodic memory (Cabeza et al., 2012; Hutchinson et al., 2014; Rugg & Vilberg, 2013; Shimamura, 2011; Uncapher et al., 2014). One theoretical framework, Cortical Binding of Relational Activity (CoBRA), suggests that the vPPC acts as a convergence zone that binds event features within the neocortex (Shimamura, 2011). This integrative process is established after encoding and creates cortical-cortical bindings that link event features related to an episodic memory. These links are thus functionally similar to MTL bindings, though MTL bindings are established quickly at the time of stimulus encoding, whereas vPPC bindings develop more gradually and are subject to inadvertent association (e.g., false memories). I propose that in order to establish or consolidate long-lasting episodic memories, it is necessary to form these vPPC links to event features stored in distributed regions in the neocortex (see Fig. 1). During retrieval, vPPC bindings facilitate the coactivation (i.e., pattern completion) of an ensemble of event features that make up an episodic memory.

Lateral and ventral view of the neocortex. According to the Cortical Binding of Relational Activity (CoBRA) framework, episodic retrieval is initiated by top-down search of stored event features in the prefrontal cortex (PFC). This search is facilitated by relational bindings in the medial temporal lobe (MTL). The ventral posterior parietal cortex (vPPC) acts as a convergence zone that links event features within the neocortex. These links are formed through reactivation or replay of episodic ensembles after initial encoding and later facilitate in the reinstatement (i.e., pattern completion) of episodic ensembles during retrieval.
The vPPC is in a prime location for binding event features, as it is located between visual ventral and dorsal paths that are known to represent object and spatial features, respectively (Anderson, 1987; Seghier, 2013). It is also near cortical regions that represent phonological and semantic information stored in the temporal lobes. Recent findings point to the angular gyrus, located in the posterior part of the vPPC, as the focus of retrieval-related activations (see Elman, Marian, Klostermann, Verstaan, & Shimamura, 2012; Hutchinson et al., 2014; Nelson, McDermott, Wig, Schlaggar, & Petersen, 2013; Uncapher et al., 2014).
Linking Encoding and Retrieval Processes
Scientists often prefer stark boundaries between psychological processes, such as between perception and memory or between encoding and retrieval. Yet with advances in research, these boundaries often become blurred, and critical distinctions that seemed rather obvious at first become muddled (for further elaborations on this issue, see Shimamura, 2010). As mentioned above, successful recollection requires both efficient encoding and successful retrieval of event features, and it is important to consider how episodic encoding and retrieval are related. Commonalities in regional activity during study and tests suggest that, to some extent, retrieval involves the reinstatement of activity patterns that occurred during encoding. For example, in an fMRI study (Johnson, McDuff, Rugg, & Norman, 2009), individuals were presented with words (e.g., “STAPLER”) while completing one of three tasks: (a) an artist task, in which they had to determine how well the object represented by the word could be drawn; (b) a function task, in which they had to determine how many ways the object represented by the word could be used; and (c) a read task, in which they had to determine what the word would sound like when spelled backward. At retrieval, they were presented with the study words intermixed with new words and asked to determine if each word was presented earlier and, if so, how strongly they remembered the word. For a strongly remembered item, the pattern of cortical activity during retrieval could be used to identify the study task (artist, function, or read) that was used to encode the item. This finding suggests that retrieval depends on the reinstatement of the processes and features used during encoding, a phenomenon often described as transfer-appropriate processing (Morris, Bransford, & Franks, 1977; Roediger, Gallo, & Geraci, 2002).
Other findings, however, suggest a disconnect between activity patterns during encoding and retrieval. In particular, encoding activity in the vPPC, as well as in the posteromedial cortex (PMC), which includes the retrosplenial cortex and posterior cingulate gyrus, is often negatively correlated with subsequent memory performance (Daselaar, Prince, & Cabeza, 2004; Uncapher & Wagner, 2009). That is, efficient encoding depends on these regions being suppressed or deactivated, despite the fact that heightened activity in these same regions during retrieval is commensurate with stronger memories. The difference between study- and test-phase patterns of activity has been called the encoding-retrieval flip (Cabeza et al., 2012), and various theories have been proposed to explain the phenomenon.
In particular, a neural circuit described as the default-mode network (for a review, see Buckner, 2012) has been implicated in both regional deactivations during encoding and increased activations during retrieval. The default-mode network, which includes the vPPC, the PMC, the MTL and the medial PFC, was christened as such because the network is more active during resting (default) states than in moments that require attending to sensory stimuli. It has now been established that these regions are recruited for a variety of mindful events, such as mind-wandering, episodic retrieval, perspective taking, and future planning (Buckner, 2012). Presumably, during encoding, it is better to focus on the sensory stimulus itself than to engage in mindful processes that may be distractive. Thus, if at times during stimulus presentations one mind-wanders (i.e., activates the vPPC and other default-network regions), then it is likely that subsequent memory for stimuli presented during those times will be poor. To the extent that the vPPC is involved in establishing critical links between event features, as described by CoBRA (Shimamura, 2011), it is predicted that soon after initial encoding, vPPC activity should be positively correlated with successful memory.
A recent study confirmed the positive role of the vPPC soon after initial encoding (Elman, Rosner, Cohn-Sheehy, Cerreta, & Shimamura, 2013). Individuals were presented with photographs of scenes and, immediately following stimulus presentation, were given one of two cues—“remember” or “ignore.” As predicted in earlier studies, during stimulus presentations, the angular gyrus was more deactivated for subsequently remembered items (hits) compared with subsequently forgotten items (misses). Yet immediately following the stimulus, when participants were given the “remember” cue, vPPC activity was positively correlated with subsequent memory (see Fig. 2). Thus, a flip was observed in vPPC activity between the stimulus and cue phases. This finding demonstrates that even during encoding (when a subject is asked to remember an item), increased vPPC activity is conducive to better memory. Given these new findings, the “encoding-retrieval flip” is a misnomer, as the flip can occur during encoding between stimulus presentation and elaborative cuing. One could argue that during elaborative cuing, the vPPC is active because individuals are merely “retrieving” information. Yet whenever individuals access a memory—during stimulus presentation, elaborative cuing, and long-term retrieval—they are “retrieving” information in this sense. What is clear is that vPPC activity is mediating successful memory formation even at the time of encoding.
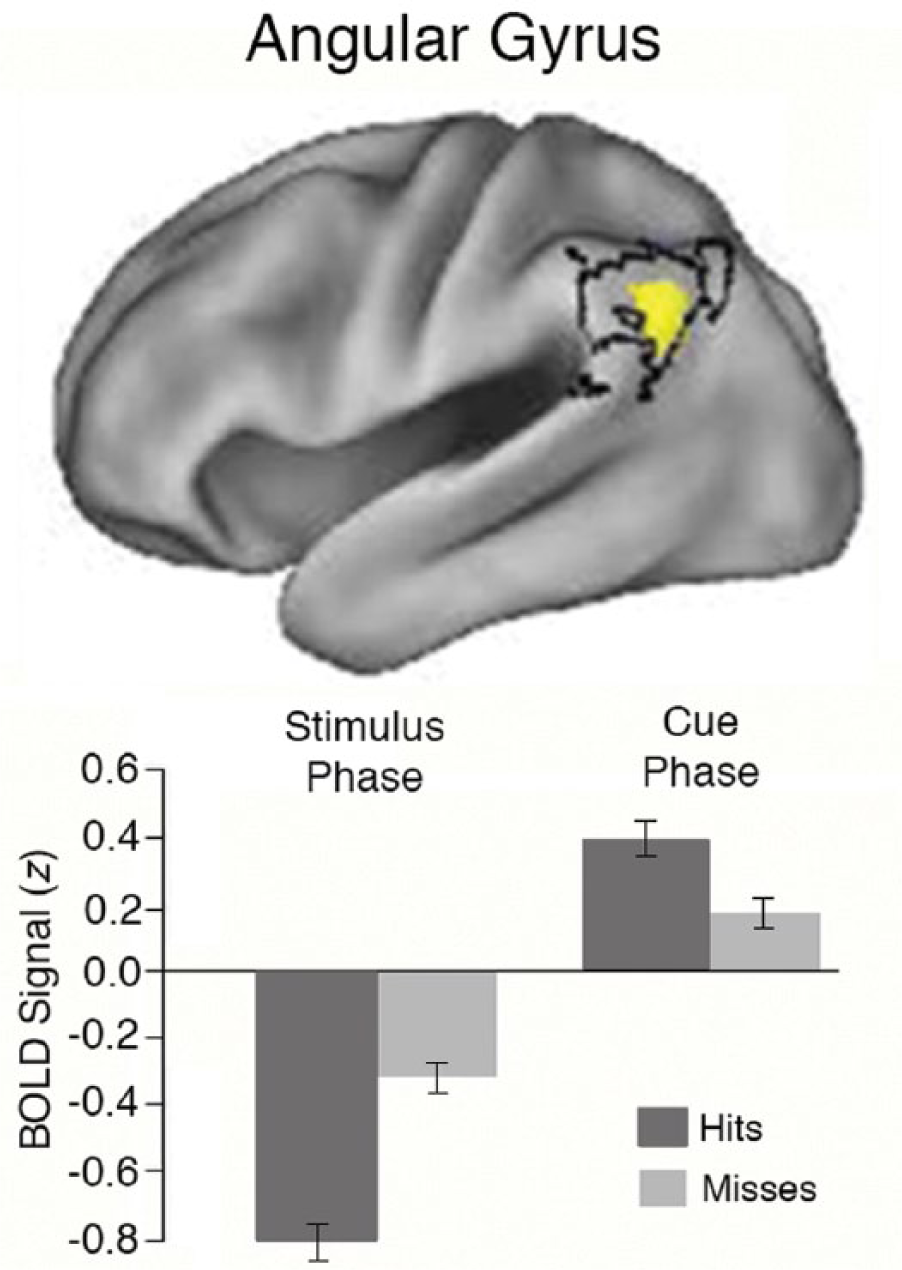
Lateral view of the neocortex showing decreased activity in the angular gyrus during stimulus presentation for subsequently correct items. As shown in the graph, this effect (hits – misses) flips immediately after stimulus presentation when subjects are instructed to remember the item (data shown here are drawn from Elman, Rosner, Cohn-Sheehy, Cerreta, & Shimamura, 2013). Error bars represent standard errors. BOLD signal = blood-oxygen-level dependent signal.
In many respects, cortical binding in the vPPC acts as a primer after initial encoding by reinforcing durable and long-lasting episodic memories. It is therefore predicted that the contribution of the vPPC in episodic retrieval will increase with the age of the memory. Although vPPC retrieval activity can be observed following short delays, it may be that with time (and more replay), long-lasting episodic memories become fully bound by vPPC bindings. Interestingly, in an electroencephalography (EEG) study, Saletin, Goldstein, and Walker (2011) found that sleep-spindle activity recorded over the left parietal cortex during a nap between study and test phases was correlated with subsequent retrieval. This finding suggests that a possible function of sleep-enhanced consolidation may be to facilitate vPPC bindings after encoding.
Given the critical role of vPPC activity during memory encoding and retrieval, why don’t individuals with vPPC lesions exhibit severely impaired episodic memory? As mentioned above, such patients do exhibit less-detailed autobiographical recollections (Hower et al., 2014; Simons et al., 2010). Yet their impairment certainly pales in comparison to that of patients with MTL lesions. This seeming discrepancy may be attributed to the fact that MTL bindings must be established during initial encoding for an episodic memory to be established. Thereafter, vPPC bindings contribute to the reinforcement of these memories. For most episodic memories, retrieval through pattern completion would occur through both MTL and vPPC bindings. It is therefore predicted that a combined lesion of both the MTL and vPPC regions would result in severe deficits in episodic retrieval. The neurological cases who most closely resemble patients with such combined lesions are patients with Alzheimer’s disease, who do exhibit severe episodic-retrieval deficits (Kopelman, 1989).
It’s a Whole-Brain Issue, Stupid!
Considering the many brain regions involved in the encoding, storage, and retrieval of episodic memories, it is clear that our ability to remember the past is a whole-brain experience. Regions within the PFC, the MTL, and the vPPC are critical, yet they must interact with the many cortical sites that store the sensory, semantic, and emotional features that make up an episodic memory. It is also clear that these brain circuits engage in a highly dynamic and integrative orchestration of activations during both encoding and retrieval. As investigations and technologies advance, it is likely that the functional properties of subunits within these cortical regions will be found to be more specifically linked to our ability to remember the past.
In this period of neurocognitive research, rather than simply attributing one brain region to one mental process, it is incumbent on the psychological scientist to consider multiple properties of any given circuitry. Indeed, it is clear that the networks described above are not only recruited for the purposes of remembering the past, but are also involved in other mindful processes, such as when we engage in perspective taking or future planning. Analyzing how multiple brain regions interact under multiple task demands has the immense potential of breaking away from the simplistic “phrenological” (i.e., one brain region, one function) perspective that has been used as a criticism of biological approaches to cognitive function.
Footnotes
Declaration of Conflicting Interests
The author declared no conflicts of interest with respect to the authorship or the publication of this article.