
Abstract
Healthy aging entails selective losses in visual attention, including the ability to filter clutter, divide attention between inputs, and search for configurations or conjunctions of features. A model of attention as a competition to influence neurons in the visual brain provides a framework for understanding these effects. Under the model, competition is necessary to disambiguate neural responses and resolve object details when multiple stimuli fall within the same visual receptive fields. A pattern of perceptual interference between attended stimuli in close spatial proximity with one another appears to be a psychophysical marker of this competition. Studies of divided visual attention in young and older adults show pronounced age-related increases in the strength of spatial interference between attended items, but only in the presence of clutter. Results suggest that inefficient competition for selection contributes to older adults’ visual attentional difficulties, compromising the ability to resolve details of multiple stimuli within small regions of the visual field. The conceptualization of attention as a competition for selection may thus provide a framework for understanding and assessing age-related attention losses.
Visual attention selects critical information for privileged processing, filtering away less important inputs. This selectivity protects the brain from overload (Broadbent, 1958; Tsotsos, 1990) and is a crucial element of adaptive visual behavior. But losses of visual attention seem to accompany even healthy aging and have been linked to increased risk of traffic crash and other forms of compromised everyday function (Kosnik, Sekuler, & Kline, 1990; Owsley et al., 1998). Age-related attentional changes include difficulty filtering visual clutter (Scialfa, Kline, & Lyman, 1987), dividing attention between multiple competing inputs (Ben-David, Eidels, & Donkin, 2014), and searching for conjunctions of visual features (Plude & Doussard-Roosevelt, 1989). Other forms of visual attentional ability, though, remain intact with age, including the ability to detect salient visual objects (Plude & Doussard-Roosevelt, 1989) and the ability to guide attention on the basis of features such as color (Humphrey & Kramer, 1997).
Developments in computational and neurobiological models of visual attention may help explain this pattern of effects. Current theory explains visual selective attention as a competition between objects for representation by neurons in the extrastriate cortex, a region of the brain responsible for processing complex visual information (Desimone & Duncan, 1995; Luck, Girelli, McDermott, & Ford, 1997). Competition is necessary to disambiguate neural responses when multiple objects fall within the same receptive field. In the absence of selective attention, extrastriate neural responses will tend to conflate the properties of any two or more stimuli within a cell’s receptive field, producing a low-quality representation. Competitive interactions allow a single object to dominate a neuron’s response train (Moran & Desimone, 1985). Bottom-up processes tilt the competition in favor of salient stimuli, and top-down processes bias it toward stimuli that match the observer’s attentional set (Beck & Kastner, 2005; Chelazzi, Duncan, Miller, & Desimone, 1998). Competition is necessary only when task demands require the observer to resolve the details of a target object, and not when the task can be performed on the basis of a coarse representation (Luck et al., 1997; Tsotsos, Rodríguez-Sánchez, Rothenstein, & Simine, 2008). Accordingly, the effects of competition appear to emerge in reentrant processing, as attention feeds back from higher to lower regions of the visual cortex to refine the percept produced by a first wave of activation (Boehler, Tsotsos, Schoenfeld, Heinze, & Hopf, 2009). Various algorithms have been suggested for instantiating competition for selection, including winner-take-all neural interactions (Tsotsos, 2011) and neural response normalization (Maunsell, 2015). We will refer to the broad idea that objects compete for representation in the visual cortex as the competitive-selection model of attention (Knudsen, 2007).
A Psychophysical Marker of Competitive Selection
Although the competitive-selection model has largely been built on neurobiological data and computational analyses, competition manifests in perceptual judgments as well. Under the model, the strength of competition between stimuli depends on the number of receptive fields they share (Luck et al., 1997; Tsotsos, 2011). The ability to divide attention between two objects should therefore depend on their spatial relationship. Items that are distant from one another in the visual field will fall largely within separate pools of extrastriate receptive fields, making them perceptible simultaneously and with little mutual interference. Objects that are very near one another will fall almost entirely within a common pool of receptive fields, necessitating strong competition between them and making them difficult to resolve in parallel. A decrease in perceptual quality that occurs as attended items are moved into closer proximity with one another therefore provides a psychophysical marker of competitive selection.
Figure 1a illustrates the procedure from a study demonstrating this effect in young adult participants (McCarley, Mounts, & Kramer, 2007). On each trial, spatial cues directed the participants’ attention to two circles within a ring of circles; a letter then appeared inside each circle. The two cued letters were always identical, and the participants were asked to simply identify them as quickly as possible on each trial. Performance was analyzed as a function of the spatial separation between targets, and data were compared to those from a control condition in which only a single target item was cued.
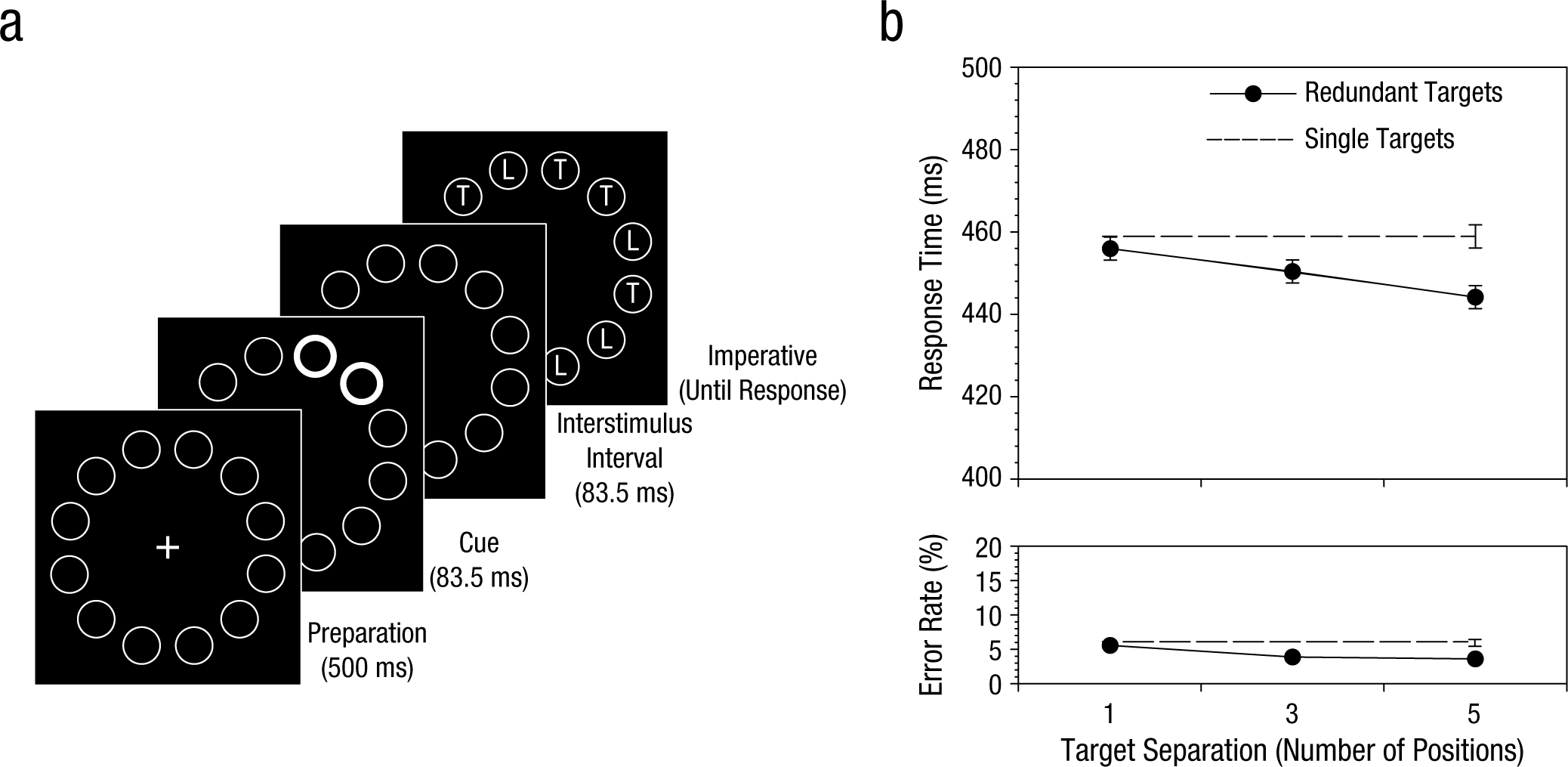
Sample stimuli and procedure (a) and results (b) from a study of divided visual attention (McCarley, Mounts, & Kramer, 2007). On each trial, participants judged the identity of either a single target letter or a pair of redundant target letters. Redundant targets varied in their degree of separation. Adapted from “Spatially-Mediated Capacity Limits in Attentive Visual Perception,” by J. S. McCarley, J. R. W. Mounts, and A. F. Kramer, 2007, Acta Psychologica, 126, pp. 104 (a) and 106 (b). Copyright 2007 by Elsevier. Adapted with permission.
Figure 1b presents mean response times (RTs) and error rates for participants’ judgments. Consistent with the competitive-selection model, responses became slower and error rates increased as the separation between targets decreased. More intriguingly, data showed a redundancy gain—that is, faster responses for multiple-target trials than for single-target trials—when targets were spatially separated, but not when they were in close proximity to one another. A redundancy gain is generally taken as evidence that multiple targets are identified in parallel and with spare processing capacity relative to single targets (Townsend & Nozawa, 1995). The absence of a redundancy gain at small target separations suggests that capacity within small regions of the visual field is fixed and that targets in close proximity are therefore forced to trade off capacity in a nearly one-to-one fashion.
A more recent study (Yamani, McCarley, Mounts, & Kramer, 2013) explored this effect more carefully, using a similar task in conjunction with a diagnostic procedure for distinguishing parallel from serial processing architectures. Results confirmed that processing was parallel when attended items were put in weak competition for selection but became serial when the items to be attended were in close proximity within very dense displays. When competition for selection was strong, participants attended to task-relevant objects one at a time.
Other studies have also demonstrated spatial interference between multiple attended items (e.g., Caputo & Guerra, 1998; Cutzu & Tsotsos, 2003; Mounts, 2000) and explored its characteristics and boundaries. As the competitive-selection model predicts, the effect varies with the relative salience of the attended items (Mounts & Gavett, 2004) and with the observer’s attentional set (Mounts, McCarley, & Terech, 2007). Tellingly, the effect appears to emerge in reentrant processing (Steelman-Allen, McCarley, & Mounts, 2009) and only when observers are asked to resolve the properties of individual objects, not when they are asked to simply detect gross stimulus features (McCarley & Mounts, 2007). Thus, interference appears to arise specifically from the need to disambiguate individual objects. Finally, consistent again with the model, the psychophysical effects of spatial interference between attended items are accompanied by decreases in the amplitude of the N2pc, an electrophysiological marker of attentional selection with its locus in the extrastriate cortex (Hilimire, Mounts, Parks, & Corballis, 2009).
Aging and Competitive Selection
Competition for selection is unnecessary when a target appears in the absence of clutter or distracting objects and is likely to be highly efficient when strong bottom-up or top-down guidance allows a target item to dominate any competing stimuli. Interestingly, these are also the conditions under which visual attentional performance is generally resilient with age. Conversely, competitive interactions are necessary when a target is embedded in competing stimuli and are inefficient when no strong bottom-up or top-down guidance is available to aid selection. These are the conditions under which age differences in visual attention are most evident. All this suggests that older adults’ visual attentional losses may lie, at least in part, in the process of competitive selection. Experiments comparing the strength of spatial attentional interference in young and older adults have lent support to this possibility.
In an early study (McCarley, Mounts, & Kramer, 2004), stimuli were colored letters, one red and one green, embedded among gray fillers. Young adult and older adult participants were asked to make same/different identity judgments of the colored items, with the spatial separation between colored items varying across trials. RTs and error rates for both age groups increased as the target separation decreased, consistent with the results described above. These effects, though, were disproportionately large for the older adults. Whereas the young adults’ error rates increased by roughly 50% from the largest to the smallest values of target separation, for example, the older adults’ increased by approximately 250%. This interaction of age with separation persisted when data were transformed to account for age-group differences in mean RTs, indicating that the effect was not simply a consequence of generalized age differences in processing speed. The interaction disappeared in a control task that used the same displays but asked participants to attend to only one of the two colored items, confirming that the disproportionate costs of target proximity for older adults were tied to the demands of dividing attention between targets.
Recent work extended these results, demonstrating that inefficient competitive selection contributes to older adults’ difficulty seeing in clutter (McCarley, Yamani, Kramer, & Mounts, 2012). Participants again made same/different shape judgments of color-cued characters that varied in separation across trials. On some trials, the targets appeared embedded in gray clutter. On other trials, targets appeared by themselves in an uncluttered display. Expectations were that clutter would compete with targets for perceptual representation, forcing more careful target resolution and engendering stronger interference than seen in uncluttered displays. Displays were again brief, minimizing the chance of eye movements and ensuring that the task was challenging.
Figure 2 presents error rates for participants’ judgments, collapsed across four experimental replications using a variety of stimuli. Data showed pronounced overadditive effects of age, clutter, and target separation. In the absence of clutter, age-group differences were small, and neither group showed large effects of target separation. In contrast, though both groups suffered performance losses and showed costs of target proximity in cluttered displays, these effects were substantially larger for older adults than for young adults. RTs showed a similar pattern. The disproportionate costs of clutter and target proximity to older adults persisted after data were transformed to compensate for differences in average processing speed across age groups, indicating again that the effects were not the result of generalized slowing in the older adults’ performance.
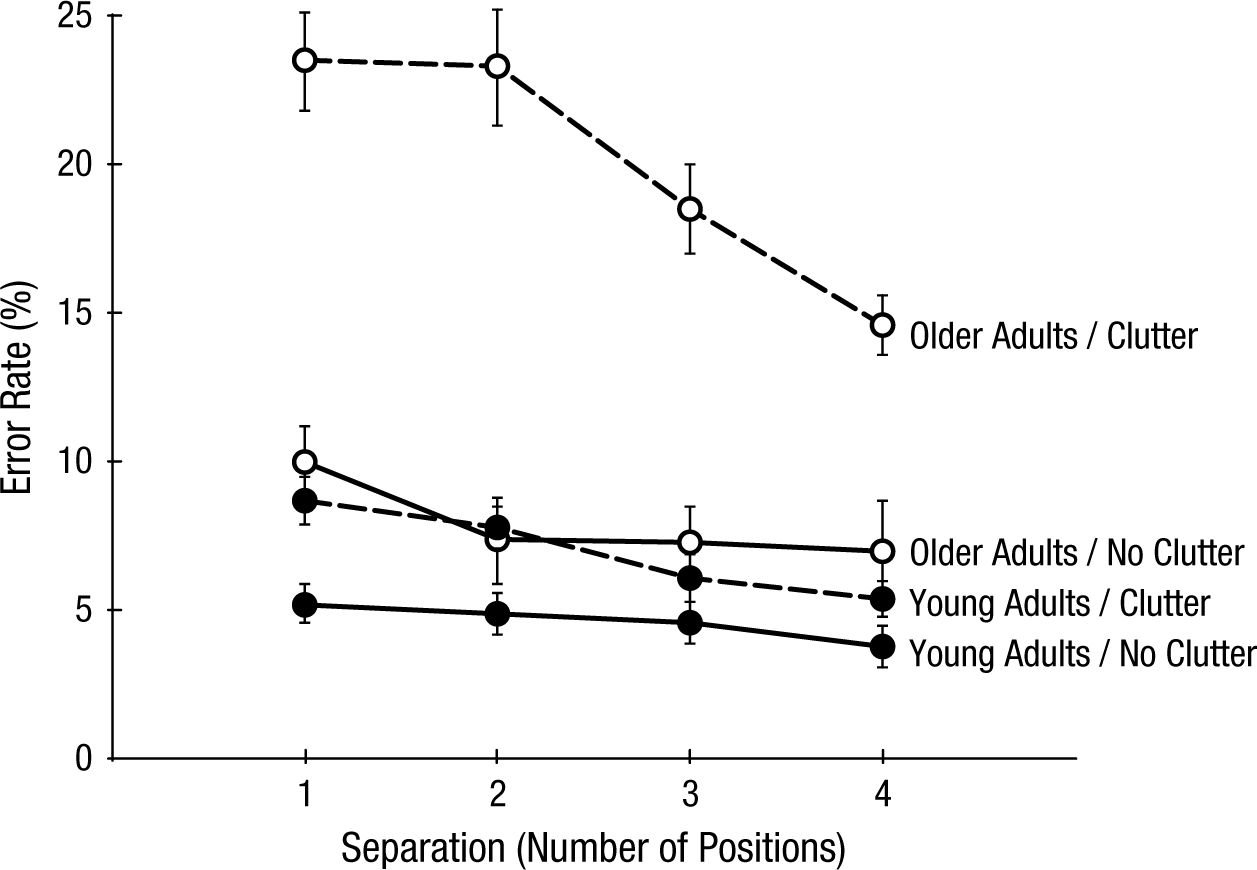
Error rates for shape judgments in a divided-attention task (McCarley, Yamani, Kramer, & Mounts, 2012).
Conclusions
Older adults experience losses in selective elements of visual attention, including the ability to divide attention between objects, the ability to process information in clutter, and the ability to search for conjunctions or arrangements of visual features. These difficulties seem to arise, at least in part, in competition for represen-tation in the visual cortex: A psychophysical marker of competitive selection shows disproportionate age-related changes, but only within cluttered displays and when participants divide attention between multiple stimuli.
An account of visual attention as a competition for selection thus offers a framework for understanding the age-related degradation of some attentional abilities and preservation of others. Under this view, visual performance will be robust in healthy aging when judgments can be based on a rough, feed-forward percept, without need for careful object resolution, or when bottom-up or top-down biases allow a single target object to dominate competing stimuli. Age losses will obtain when non-target stimuli compete for selection with a target or when attention is divided between competing targets, particularly when competing stimuli lie in close proximity.
These conclusions suggest avenues of exploring and remediating visual attentional decline in older adults. Along with psychophysical research to further document the perceptual consequences of age-related changes in competitive selection, electrophysiological studies will be useful to compare neural markers of competition for selection in young and older adults; though markers of visual attentional selection are known to be attenuated and delayed in older adults (Lorenzo-López, Amenedo, & Cadaveira, 2008), these effects have not yet been linked to spatially mediated competition for selection. Age-related changes in the electrophysiological patterns of spatial interference between attended items would provide converging evidence that older adults’ attentional losses arise in the process of competition for selection.
Neurocomputational modeling of age-related changes will be valuable as well—first, to identify the causes of inefficient competitive selection in the aging visual system, and second, to simulate the perceptual consequences of older adults’ impaired visual attention. Potential hints at the mechanisms of inefficient selection come from single-cell data indicating that stimulus selectivity is compromised and signal-to-noise ratios are reduced in the extrastriate cortex of senescent monkeys (Yu, Wang, Li, Zhou, & Leventhal, 2006). As the competitive-selection model anticipates, these effects are disproportionate to losses in earlier visual regions and may be linked to failures of intracortical inhibition. Simulations within a formal computational account such as Tsotsos’s (2011) Selective Tuning model will reveal whether such effects can in fact produce the age-related psychophysical losses described above and, in doing so, may be able to identify more subtle and precise signatures of visual attentional deficits in older adults.
Finally, research will be necessary to test whether inefficient competitive selection contributes to age-related difficulties in everyday perceptual tasks such as finding and recognizing objects among clutter (Kosnik et al., 1990). Existing tests of visual attention predict older adults’ daily functioning well, largely by measuring visual processing speed (Owsley, 2013). As noted above, though, age-related losses in the efficiency of competitive selection appear to reflect deficits beyond those that come from changes in processing speed alone. Research will be valuable to determine if and how these deficits account for additional variance in everyday function. Evidence linking changes in everyday vision to changes in competitive selection would inform the assessment of age-related vision losses and help guide the design of visual displays, interfaces, and environments for older adults.
Footnotes
Declaration of Conflicting Interests
The authors declared that they had no conflicts of interest with respect to their authorship or the publication of this article.