
Abstract
Binocular rivalry (BR) refers to the spontaneous, unpredictable fluctuations in visual awareness provoked by dissimilar stimulation of the two eyes. Reports of the phenomenon date back several centuries, but interest in BR has exploded in recent years as researchers in diverse disciplines—psychology, neuroscience, medicine, philosophy—have found reasons to study it. New ideas about BR have emerged, sparking controversies about its neural bases, which may be resolved thanks to new methodological developments. This essay provides a synopsis of some key empirically determined aspects of BR as well as an overview of theoretical developments in this field. Work published during the past decade or so is emphasized (and explicitly referenced); earlier key findings are mentioned and referenced in the annotated bibliography included in the Supplemental Material.
Over the five decades spanning my professional career in experimental psychology, I have maintained an enduring fascination—indeed, obsession—with the visual phenomenon known as binocular rivalry (BR). It was the topic of my dissertation at Vanderbilt University, and it has maintained my fascination ever since. Now, being on a glide path to retirement, I have an opportunity to reflect on what researchers have learned about BR and to surmise why it has generated broad interest within diverse fields, including psychology, neuroscience, medical science, and philosophy. This essay emphasizes recent findings and ideas about BR; the annotated bibliography in the Supplemental Material available online lists earlier work on BR. I start by describing my first encounter with this beguiling phenomenon.
The Mind’s Visual Disappearance Trick
It was Robert Fox who introduced me to BR in 1967, when I toured his Vanderbilt University lab as an entering graduate student. The experience remains vividly etched in my memory: With me seated in front of a peculiar optical device with a separate viewing port for each eye, Fox illuminated the pictures in the two ports and asked me what I saw. Unaware that my two eyes were being shown different pictures, I opined that he was showing me a sequence in which the picture of a tree dissolved into the picture of a person’s face, and vice versa, over time. Each picture remained present for several seconds before giving way to the other, but the precise exposure durations were not always the same. Chuckling at my description, Fox next instructed me to alternately open and close my two eyes, thus revealing the origin of the trick that fooled me: It was my mind—not the optical device—that was generating these alternating states of visual perception. My awareness, in other words, was fluctuating even though the physical stimulation remained unchanged. And, indeed, that is the hallmark feat of magic that fascinates everyone upon first experiencing BR. I am deeply grateful to Fox for serving as the magician who sparked my curiosity about BR and who, then, taught me how to explore this and other aspects of perception using psychophysical techniques.
Binocular Rivalry and Perceptual Multistability
BR belongs to a family of visual phenomena characterized by fluctuations in perception despite unchanging physical stimulation: What you are seeing, in other words, is not what you are viewing. This fascinating family of phenomena collectively exhibit what is called multistable perception (see Fig. 1).
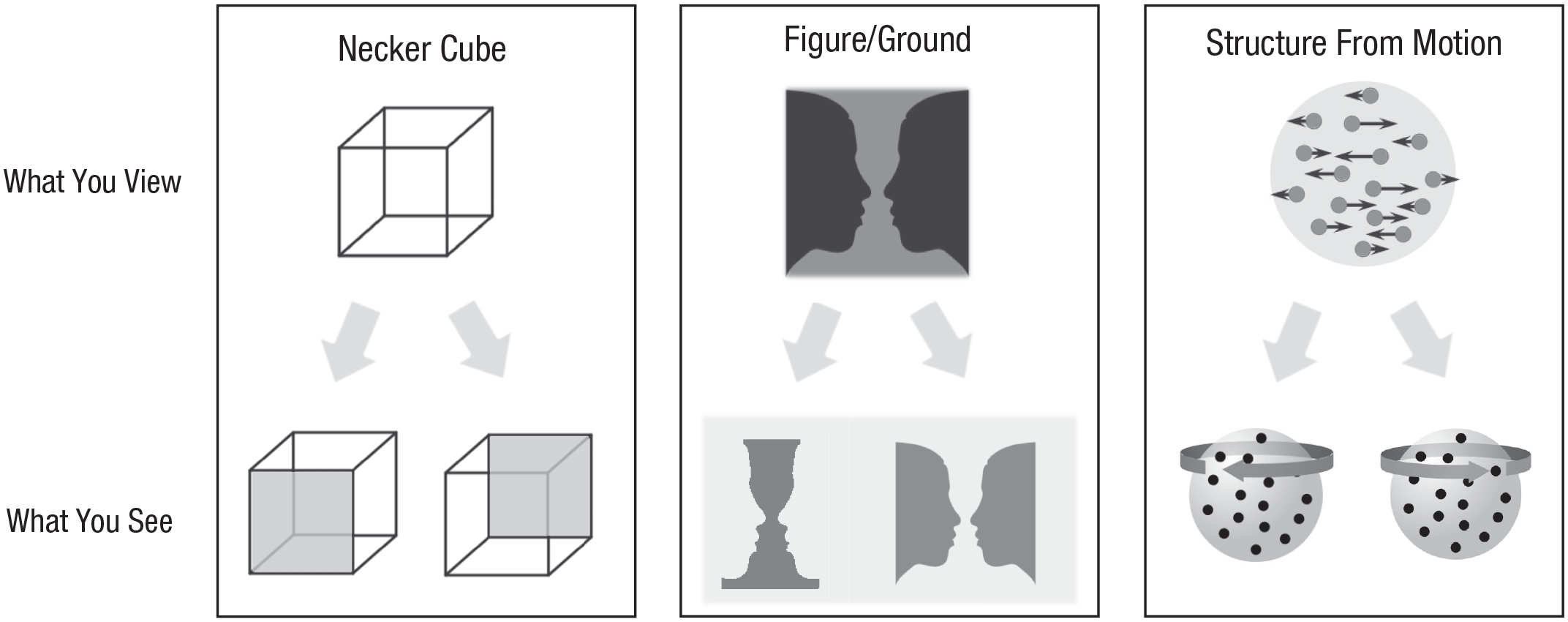
Examples of visual figures and viewing situations in which an ambiguous figure or ambiguous animation provokes multistable perception. In the case of the Necker cube, the absence of cues indicating which lines are in front of others results in alternating perception of two orientations of the cube. The gray shading indicates the perceived front surface of the cube. The middle panel illustrates ambiguity in what is figure and what is ground. Viewers of this illustration may perceive it as alternating between the picture of a wine goblet and the picture of two faces. In the right panel, the schematic at the top denotes one frame of an animation in which dots on a globe-shaped object move laterally to the left or to the right (the arrows denote the directions of the dot motion). The bottom two schematics denote the two alternative interpretations a person may experience when viewing this animation, that is, rotation of a globe about its vertical axis in either the clockwise direction (left-hand globe) or the counterclockwise direction (right-hand globe). The thick gray arrows superimposed on the globes represent perceived direction and are not actually present in the animation. With continued viewing, the globe’s direction of rotation appears to change unpredictably over time because the “true” direction is ambiguous. Illustration by Alexander Pastukhov.
But in one important respect, BR stands apart from nearly all other members of the family. Specifically, most classic examples of multistability entail viewing a single figure that can be interpreted in one of several ways because of figure/ground or perspective ambiguity: One’s interpretation wavers, but the figure itself remains visible continuously. During BR, however, two dissimilar figures undergo reciprocal periods of complete invisibility, taking turns moving into and out of visual awareness (Fig. 2a). Interspersed between these fluctuating states of exclusive monocular awareness (i.e., dominance periods in BR parlance) may be brief transitions when one sees a dynamic mosaic-like episode when one figure sweeps the other out of visual awareness (Wilson et al., 2001). This dynamic competition for visual dominance defines the remarkable phenomenon of BR, an increasingly popular topic of study over the past 30 years (see Fig. S1 in the Supplemental Material).
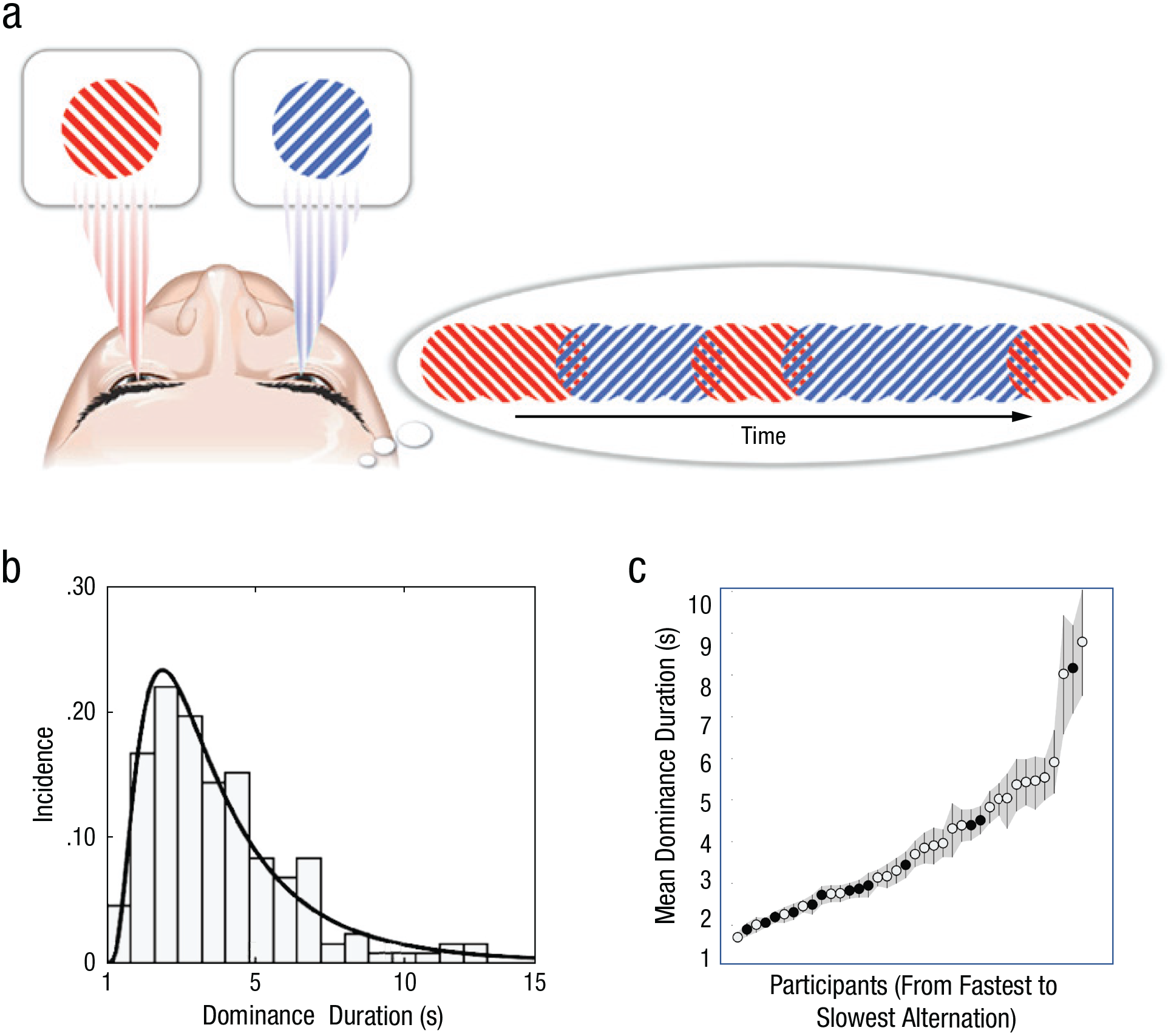
Binocular rivalry (BR) dynamics. As illustrated in (a), when the two eyes view dissimilar monocular figures (dichoptic viewing), visual awareness of the stimuli (dominance) fluctuates unpredictably over time. The timeline on the right illustrates schematically what an individual might be experiencing and reporting in an experimental study. Illustration on the left adapted from Mitchell et al. (2004), p. 411. The histogram in (b) is an example of the distribution of dominance durations measured during extended viewing of BR. This particular histogram comes from one of 38 people tested in a recent study (Blake et al., 2019). The solid line shows the best-fitting log normal curve, the canonical shape for duration distributions in BR and other forms of multistable perception (Brascamp et al., 2005). The graph in (c) shows the average dominance duration for each of the 38 participants in the same study, in order from the participant who experienced the fastest alternations of dominance (left) to the participant who experienced the slowest alternations (right); the shading denotes 95% confidence intervals around those means. Open versus filled circles represent the two locations at which data were collected. The rate at which rival states alternate over time is inversely related to the durations of those states: Participants who perceived faster alternations experienced briefer state durations, on average. Graphs adapted from “Individual Differences in Continuous Flash Suppression: Potency and Linkages to Binocular Rivalry Dynamics,” by R. Blake, R. Goodman, A. Tomarken, and H.-W. Kim, 2019, Vision Research, 160, p. 14 (https://doi.org/10.1016/j.visres.2019.04.003). Copyright 2019 by Elsevier. Adapted with permission.
Two well-established features characterize BR: (a) the unpredictability of the durations of visual dominance and suppression as BR unfolds over time and (b) the wide range of individual differences in the rate at which people experience reversals in dominance during BR. These two characteristics are exemplified in the graphs in Figures 2b and 2c. Measured over extended periods of viewing dissimilar monocular images (i.e., dichoptic stimulation), successive periods of left-eye and right-eye dominance and of mixture states define the dynamics of BR. From an ecological perspective, this breakdown in stable visual perception can be construed as the consequence of violation of a fundamental physical constraint: Two different objects cannot exist at the same location in space at the same time, and yet that is exactly the information that the two eyes are sending to the brain. Figuratively speaking, the brain reacts to this impossibility by selecting one of the two alternatives and updating that selection from time to time, thus producing a time series of periods of exclusive monocular visibility interspersed with brief periods of transition from one alternative to the other as the previously suppressed image emerges into dominance, supplanting the previously dominant stimulus in a wavelike fashion.
One might construe the irregularity of BR state durations and the large range of individual differences in alternation rates as distracting nuisances when trying to develop a causal account of BR dynamics, but that attitude may be shortsighted. In fact, the within-subject consistency among session-to-session measures of BR is sufficiently high to enable one to distinguish a given person’s data from another person’s with exceedingly high reliability (Sy et al., 2017). This reliability allows investigators to exploit an individual-differences approach to examine links between BR dynamics and physiological measures such as alpha oscillations in electroencephalography (EEG) signals (Katyal et al., 2019) and to estimate the contribution of genetics to alternation rates (Miller et al., 2010).
Factors Influencing BR Dynamics
Thanks to decades of research, it is now possible to enumerate a host of factors that reliably govern BR dynamics and the incidence of mixtures. Those factors can be categorized as intrinsic (i.e., inherent within the stimuli themselves) and extrinsic (i.e., supplemental perceptual information accompanying those stimuli or factors inherent to the viewer). They are summarized below, and published articles documenting these influences are listed in the Supplemental Material (Appendix S1).
Intrinsic determinants of BR dynamics include (a) relative strengths of the dissimilar monocular images (e.g., low vs. high contrast patterns), (b) figural complexity of the images (e.g., natural scenes vs. 2D spatial noise), (c) the temporal character of the patterns (e.g., flickering vs. stationary elements), (d) large differences in chromaticity of dissimilar patterns viewed by the two eyes (e.g., red vs. green patterns), and (e) differences in the affective valence of competing figures (e.g., neutral vs. angry faces). Among the extrinsic factors influencing BR dynamics are (a) contextual cues, such as shadows or movement within the vicinity of ongoing BR that favors one stimulus interpretation over the other; (b) the presence of accompanying sounds, tactile sensations, or odors congruent with one of the two rival stimuli; (c) the relevance of a given stimulus established during previous performance of a non-BR task (e.g., visual search); (d) the viewer’s history of rewards or penalties associated with a given rival stimulus; and (e) explicit, focused attention on one of the two competing stimuli. The potency of these extrinsic factors encourages the view that BR is modulated by top-down influences (e.g., Wolf & Hochstein, 2011), including ones arising within nonvisual areas of the brain. It remains arguable, however, whether those influences include abstract conceptual information (e.g., word associations), and if so, whether that influence is attributable to increased attention that is engaged only when the conceptually meaningful stimulus is dominant. The next section revisits this issue in the context of suppression of awareness.
Observer-related factors also play a role in determining BR dynamics. For example, alternation rate tends to slow as people get older, just as it does when people have ingested alcohol, the hallucinogen ayahuasca, or the serotonin inhibitor psilocybin. On the other hand, the stimulant caffeine temporarily accelerates BR alternation. Alternation rates measured while a viewer is walking are a good bit slower than alternation rates measured when that individual is sitting, even though the BR patterns are equivalent in the two conditions. There is mixed evidence whether alternation rates are generally reduced in people within the autism spectrum, but consistent evidence that people diagnosed with schizophrenia, bipolar disorder, major depression, or obsessive-compulsive disorder exhibit slowed alternation. It remains to be learned how those conditions relate to neural events underlying BR. One suggestive finding is that normal adults exhibiting intrinsically slower switch rates during BR tend to have higher concentrations of the inhibitory neurotransmitter gamma-aminobutyric acid (GABA) within the visual cortex.
One particularly curious, revealing feature of BR dynamics deserves mention. When dissimilar monocular stimuli are viewed for periods of time much longer than the conventional 1- or 2-min trial duration, the initially high incidence of phases of exclusive dominance of one eye’s view or the other eye’s view declines steadily as those exclusivity phases give way to longer periods of mixed dominance (see Fig. S2 in the Supplemental Material). Moreover, recovery from this prevalence of mixtures back to conventional BR requires extended viewing under normal binocular conditions. These observations implicate experience-based plasticity within inhibitory processes mediating BR suppression (Klink et al., 2010).
What Is Suppressed During Rivalry?
This is the question that inspired my dissertation work on BR (Blake & Fox, 1974), and it continues to be a popular topic of investigation. Stated succinctly, during BR, a normally visible, potentially engaging, and informative visual stimulus can remain suppressed from awareness for seconds at a time: To what extent does information embodied in the neural representations of such a subliminal stimulus continue to affect visual processing?
Remarkably, a large range of visual processes remain at least partially effective when the mediating visual stimulus is erased from awareness by BR (see the references in Appendix S1 in the Supplemental Material). For example, visual exposure to oriented contours, to visual motion, or to surface color can induce lingering, measurable aftereffects of that exposure even though the inducing stimulus itself has been suppressed from awareness courtesy of BR. In a similar vein, brain systems responsible for generating eye movements still manage to shift fixation to the location of a task-relevant visual stimulus or to bias the direction of tracking eye movements even when the stimulus mediating those eye movements is suppressed from awareness. On the other hand, registration of high-level semantic information associated with facial expressions, scene context, or threatening objects seems to be nullified during suppression. Two lessons learned from attempts to answer this seemingly simple question about what survives suppression are that (a) it is important to acknowledge contributions of low-level feature elements defining objects and (b) during BR, a complex stimulus can be fractionated, such that some features (e.g., color) remain visible while others (e.g., contour orientation) succumb to suppression (Hong & Blake, 2009; Moors et al., 2017).
Theoretical Work on Binocular Rivalry
The characteristics just summarized provide essential grist for developing and testing ideas about possible mechanisms underlying BR. The products of those efforts have evolved over the years, starting with two distinct theoretical branches that more recently have evolved into a single, hybrid view of BR (see Fig. S3 in the Supplemental Material).
One theoretical branch treats BR as the culmination of reciprocal excitatory and inhibitory interactions between sensory representations of the left- and right- eye images; adaptation and/or neural noise contributes to the stochastic nature of BR. Some of these models view BR as a concomitant of the neural machinery mediating binocular fusion (i.e., blending of the two monocular images) and stereopsis (i.e., keen sense of 3D vision that arises when one is viewing the world through two laterally separated eyes). As a group, those theories constitute what has been termed eye-based models of rivalry. Occupying the other theoretical branch are models that construe BR as the product of competition between plausible but incompatible interpretations of what is being viewed (object rivalry), initiated within putative high-level brain areas beyond visual cortex. A good example of this class of models is the predictive-coding account developed by Hohwy et al. (2008). In that account, neither of two interpretations engendered by dissimilar monocular stimulation enjoys a greater likelihood of being correct based on current sensory evidence and expectation grounded in past experience. The resulting conflict is temporarily resolved by adoption of one interpretation over the other, but the continued physical presence of the monocular image favoring the unadopted interpretation constitutes residual error signal that thwarts stability. The conflict remains unresolved, which eventually prompts a switch in dominance. In this narratively appealing Bayesian-flavored model, error signal serves the same role as do inhibition, adaptation, and noise in eye-based models (i.e., triggering alternations in dominance over time).
At the turn of this century, ideas about BR began trending toward an amalgam of the two theoretical branches (Blake & Logothetis, 2002). Moreover, this hybrid view of BR comported comfortably with what was being learned about the neural bases of BR (Tong et al., 2006; see Fig. S4 in the Supplemental Material). Still, there is continuing debate concerning the directions of causality within the hierarchy of brain areas purportedly involved in BR (Brascamp et al., 2018). Dynamic causal modeling applied to functional MRI measurements from humans offers one strategy for assessing sequential neural activity across brain areas during perceptual bistability (Weilnhammer et al., 2021). An even more precise way to distinguish the directions of feedforward and feedback causality may be to measure neural response propagation at the millisecond scale within different layers of the cortex simultaneously throughout distributed areas of the brain (e.g., Bastos et al., 2020). Indeed, intracranial recording is a technique that can be utilized under limited conditions within awake humans performing a visual task (de Jong et al., 2020). One final thought on this emerging view combining the two branches: The distinction between low-level, eye-based accounts of BR and high-level, object-based accounts of BR has probably served its purpose but is now outdated. Within this emerging conceptual view, the notions of “low” and “high” lose their meaning once neural information begins reverberating among areas within the cortical hierarchy (e.g., Heeger, 2017).
Methodological Advances
Complementing the theoretical advances are more refined psychophysical procedures for generating and analyzing BR data. One of those procedures, called flash suppression, entails presentation of a monocular pattern followed a second or so later by the abrupt appearance of a competing pattern presented to the other eye. This staggered presentation to the two eyes guarantees initial dominance by the second pattern. Another variant is the interocular swap procedure, wherein two dissimilar monocular patterns are repetitively exchanged between the two eyes at a very rapid rate (Logothetis et al., 1996), the idea being to discern whether prolonged dominance of one pattern or the other survives eye swapping. Indeed, something resembling BR may be experienced, but only under more limited conditions than when BR is evoked without eye swapping (Lee & Blake, 1999). More recently, a third BR procedure was introduced, one that generates unusually long periods of invisibility (i.e., suppression) of one eye’s stimulus by presentation of a rapidly changing series of Mondrian-like patterns to the other eye. Called continuous flash suppression, this potent form of interocular suppression has gained popularity as a tool for studying perception outside of awareness.
Complementing those psychophysical procedures are “no report” strategies that derive indirect measures of BR dynamics (e.g., eye movements or EEG recordings) without requiring people to decide and report what they are seeing. There are developments in data analysis as well. Investigators are beginning to use reverse correlation strategies that look backward in time within BR phases to identify possible stimulus events (e.g., fluctuations in stimulus salience or intensity) or observer-related events (e.g., pupillary dilation) that might precede BR state changes. In general, being able to channel the dynamics of BR and dissect state durations in these ways afford opportunities to isolate neural events inducing BR, as well as those prompting state changes during BR.
Conclusion
Despite being caricatured as a laboratory artifact by one famous, mid-20th century perception psychologist, BR has continued to fascinate and, thus, draw scientific interest. Moreover, the variety of journals in which work on BR appears attests to the broad interest BR enjoys among diverse disciplines. Among the reasons for this widespread, enduring interest are the following:
BR has been popularly touted as a means for identifying neural concomitants of consciousness (Maier et al., 2012), a challenge seen by some researchers as tractable in the near future (Melloni et al., 2021).
BR, in conjunction with computational modeling, may offer a fruitful means for tracing the evolution of activity within hierarchically organized neural circuits involved in autonomous decision making (Oizumi et al., 2014) and for studying dynamic behavior of neural systems working within and near the margins of instability (Pastukhov et al., 2013).
BR dynamics, as indexed by alternation rate, provide a reliable biological marker associated with psychiatric conditions, including bipolar disorder (Ngo et al., 2011) and schizophrenia (Ye et al., 2019).
BR may be a crucial component of the binocular matching process whereby the brain registers and resolves ambiguities associated with binocular viewing (e.g., Goncalves & Welchman, 2017; Grossberg et al., 2008; Wilson, 2017). It is worth noting that dissimilar monocular stimulation, which triggers BR, arises also during natural binocular viewing. For instance, the two eyes can receive dissimilar monocular inputs when one is viewing an object that partially occludes another (Nakayama & Shimojo, 1990) or when one is viewing an object with a smooth, shiny surface that gives rise to differential specular reflections imaged on the two retinas (Muryy et al., 2014).
BR potentially furnishes insights about impaired binocular vision associated with conditions such as amblyopia (Bao et al., 2018) and about limitations to the procedure called monovision, which involves differential refractive correction of the two eyes to compensate for presbyopia or imbalance in refractive power following cataract surgery (Zheleznyak et al., 2015).
In the final analysis, however, the fundamental allure of BR stems from the beguiling trick it plays on us: Normally visible, potentially interesting visual figures take turns emerging and disappearing from visual awareness when, in fact, what we are looking at remains unchanged. BR, in other words, is indeed perceptual magic (Kim & Blake, 2005).
Recommended Reading
Alais, D. & Blake, R. (2005). Binocular rivalry. MIT Press. An edited volume comprising 18 chapters presenting data and theory bearing on existing controversies in the field.
Blake, R. (1989). A neural theory of binocular rivalry. Psychological Review, 96(1), 145–167. https://doi.org/10.1037/0033-295X.96.1.145. An article laying out a conceptualization of binocular rivalry as the culmination of a dynamic interaction between excitatory and inhibitory processes transpiring within early stages of visual processing.
Blake, R. (2014) Binocular rivalry updated. In J. S. Werner & L. M. Chalupa (Eds.), The new visual neurosciences (pp. 827–846). MIT Press. A chapter reviewing recent empirical and theoretical developments in the study of binocular rivalry.
Brascamp, J. W., Klink, P. C., & Levelt, W. J. M. (2015). The ‘laws’ of binocular rivalry: 50 years of Levelt’s propositions. Vision Research, 109(A), 20–37. https://doi.org/10.1016/j.visres.2015.02.019. An article updating Levelt’s doctoral dissertation, a landmark publication that laid out propositions quantifying how stimulus strength affects rivalry dynamics and that set the benchmark for theories of binocular rivalry.
Sterzer, P., Stein, T., Ludwig, K., Rothkirch, M., & Hesselmann, G. (2014). Neural processing of visual information under interocular suppression: A critical review. Frontiers in Psychology, 5, Article 453. https://doi.org/10.3389/fpsyg.2014.00453. A comprehensive introduction to binocular rivalry’s close relative continuous flash suppression, a popular tool for studying visual processing outside of awareness.
Supplemental Material
sj-docx-1-cdp-10.1177_09637214211057564 – Supplemental material for The Perceptual Magic of Binocular Rivalry
Supplemental material, sj-docx-1-cdp-10.1177_09637214211057564 for The Perceptual Magic of Binocular Rivalry by Randolph Blake in Current Directions in Psychological Science
Footnotes
Acknowledgements
I am grateful to Jan Brascamp, Tomas Knapen, George Sperling, Nicholas Wade, David Heeger, and David Alais for helpful discussion during the writing phase of this project. I also thank Kris Klink, David Leopold, Jude Mitchell, Alexander Pastukhov, and Frank Tong for artwork revised and included in portions of some of the figures.
Transparency
Action Editor: Robert L. Goldstone
Editor: Robert L. Goldstone
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.