
Abstract
Background
Childhood exposure to a high-fat diet (HFD) has long-term implications on metabolism and behaviour. Exercise and switching to a normal low-fat diet during adulthood can sometimes reverse a few biochemical consequences of HFD exposure during childhood.
Purpose
This study proposes to address the effect of aerobic exercise on HFD-induced metabolic and behavioural changes by observing the biometric, biochemical and anxiety-like behaviour assessed by elevated plus maze in adolescent male and female Wistar rats.
Methods
Fourteen female and 18 male wistar rat pups aged 35 days were divided into 3 groups [control, HFD+exercise and HFD]. The HFD and HFD+exercise alone received HFD for 10 weeks. In addition HFD +exercise were subjected to forced swimming for 30 minutes 5 days a week for 4 last weeks. At the end of 10th week, elevated plus maze recording was done to assess anxiety.
Results
We observed that both male and female Wistar rats’ body weight increased within 2 weeks of HFD exposure and continued to increase till 5th week. In male rats, after 4 weeks of exercise, the weight was comparable to that of the control group, while weight decreased in female rats. In both males and females, plasma glucose and lipid profiles were increased after HFD. However, exercise seems to lower these in the HFD + exercise group. Female HFD + exercise group rats showed a significantly higher number of exits from closed arm to open arm and climbing than their sex-matched control on Day 3.
Conclusion
This shows that the exercise ameliorated the anxiogenic behaviour in HFD-fed adolescent female rats but not in male rats to the same extent. When combined, these findings highlight that HFD and exercise throughout adolescence have sex-specific differential effects on biometric, biochemical, and anxiety-like behaviour.
Introduction
Pre-puberty and adolescence are crucial developmental periods when environmental influences, such as diet, affect physiology and metabolism. Consuming a high-fat diet (HFD) and a sedentary lifestyle contributes to metabolic syndrome and obesity in people of all ages, especially in adolescents.1–3 Interestingly, animals treated with HFD during adolescence and adulthood exhibit distinct behavioural and biochemical responses.1, 4–7 HFD impairs cognition and anxiogenic behaviour.1, 4, 5, 8, 9 An elevated plus maze (EPM) test of adult male Sprague-Dawley rats’ anxiety in response to a short-term diet high in fat, carbs or protein revealed that the diet high in fat alone decreased the rats’ anxiety. 10
In juvenile rats, neuronal plasticity arises from continuous synaptogenesis and neuronal network maturation,8, 11 making the pre-pubertal and adolescent stages susceptible to chronic disruption of hippocampal activity by HFD.12, 13 Moreover, this period corresponds to the childhood and pre-pubertal period in humans, where consuming fat-enriched palatable meals is a general practice in modern society. A study by Ferreira et al 14 demonstrates that diets high in sugar and saturated fats are more detrimental to juvenile rats than diets high in sugar alone when it comes to their effects on anxiety-related behaviours, spatial learning and memory and neurogenesis.
Exercise is an established approach to combat HFD-induced obesity, metabolic disorders and reverse behavioural consequences in humans and rodents. 15 The positive effects of exercise on the metabolic derangement caused by HFD are contentious.9, 16, 17 Typically, an acute bout of exercise can result in behavioural changes. 18 Because we wanted to assess the interventional effects during adolescence, we chose a 4-week exercise protocol, as the pups became adults after this period. Furthermore, the impact of exercise on HFD-induced anxiogenic behaviour is sparsely studied, particularly in adolescence, with one study by Maniam and Morris 19 demonstrating that providing rats with palatable food and/or exercise improved their behavioural reactions after experiencing stress (separation from dams) in their early lives. They revealed that voluntary exercise, either alone or in combination with HFD, improved the behaviour and metabolic outcomes of rats that had experienced early life stress. Though studies have assessed the consequences of exercise and HFD, the gender difference and the effect of HFD during adolescence have not been sufficiently studied. Hence, in this study, we wanted to assess the impact of 10-week supplementation of HFD coupled with 4 weeks of aerobic exercise on the biometric, biochemical and anxiety-like behaviour in age-matched adolescent male and female rats.
Methods
Animals
After obtaining approval from the Institutional Animal Ethics Committee, all the experiments were performed following the guidelines of the Committee for the Purpose of Control and Supervision of Experiments on Animals. For this study, we have used 14 female and 18 male Wistar pups belonging to different litters, aged 35 days (weaning on the 25th day + 10 days of habituation) and weighing between 30 and 50 g. The animals were in-house bred and maintained in the animal house with standard room temperature and humidity. They were kept in groups of the same sex in plastic cages with wired lids. On the 35th day, the pups were separated from the dams and randomly divided into three groups (control (standard laboratory diet), HFD + exercise and HFD alone).
Experimental Design
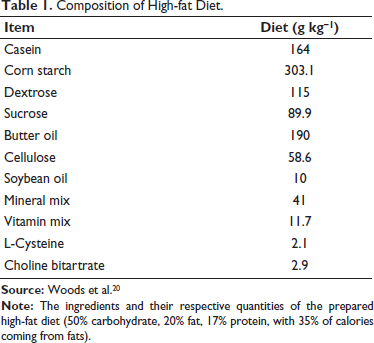
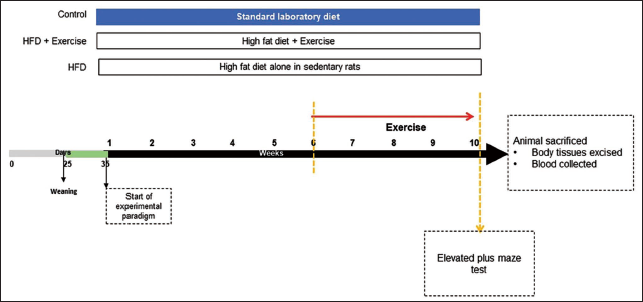
The HFD was freshly prepared in the laboratory. The composition of HFD is adapted from the previous report listed in Table 1. 20 The control adolescent rats (male [n = 6] and female [n = 5]) were provided with a standard laboratory diet and water ad libitum for 10 weeks. The HFD adolescent pups (male [n = 5] and female [n = 4]) were provided HFD and water ad libitum for 10 weeks. Both these groups of rats were not subjected to any exercise regimen but were exposed to tank water deep enough to stand in while keeping their heads out of the water. They continued to feed on standard laboratory and HFD diet for 10 weeks. The HFD + exercise adolescent pups (male [n = 7] and female [n = 5]) were provided HFD and water ad libitum for 10 weeks. From the 6th week onwards till the end of the 10th week, the HFD + exercise rats were placed in a large water tank (1.5 m diameter) with enough water not allowing them to get limb support from the tank base. The temperature of the water was maintained between 25 ± 2°C. The HFD-fed rats were exposed to forced swimming exercises for 30 min daily with a metal ring weighing 5% of their body weight (BW) attached to their torso. This amounts to moderate-intensity aerobic exercise. 21 This exercise protocol was followed 5 days a week for 4 weeks following a previous study. 22 The detailed experimental protocol is depicted in Figure 1.
Composition of High-fat Diet.
Experimental Design Representing 10 Days of Habituation in the Laboratory.
Behavioural Assessment
After the treatment protocol was over, the anxiety-like behaviour was assessed in the EPM. EPM assesses anxiety-like behaviour based on the natural tendency of rodents to prefer dark, closed places and avoid height and open spaces.
23
The time required to enter any closed arms (CA) was noted as escape latency (EL). The 3-day test paradigm to assess EL also hints at the learning ability of the animal to escape to the dark, CA. The EPM, made of wood, was elevated at 30 cm from the base height and consisted of two CA and two open arms (OA) measuring 50 cm in length. In all the cohorts of rats, the EPM was done in the light cycle at the same time (between 10
Biochemical Evaluation
After the completion of the behavioural evaluation, the overnight fasted animals were euthanised through cervical dislocation, and the blood samples were collected after a cardiac puncture. Blood was collected in ethylenediaminetetraacetic acid tubes, and the plasma obtained was used to determine fasting lipid profile and blood glucose. Plasma glucose estimation was done by the glucose oxidase–peroxidase method (Beckman Coulter analyser, CA, USA). 25 Blood samples for total cholesterol (TC) were analysed by spectrophotometric analysis (Beckman Coulter analyser, CA, USA) by the cholesterol oxidase–peroxidase method (Agappe Diagnostic Ltd, New Delhi). The same enzymatic analysis, along with an immune-inhibition assay, was employed to estimate the high-density lipoprotein (HDL; Agappe Diagnostic Ltd, New Delhi) and triglyceride (TG; Agappe Diagnostic Ltd, New Delhi). All the protocols were followed as per the manufacturer’s instructions. Plasma low-density lipoprotein (LDL) and very low-density lipoprotein (VLDL) levels were calculated, as Friedewald et al 26 mentioned earlier. After weighing the whole liver, approximately 100 g of liver tissue was cut, minced and homogenised in phosphate buffer saline (pH 7.4) using the Teflon homogeniser. The total lipid profiling from the liver sample was done using a spectrophotometric kit following the manufacturer’s protocol. 27
Weight of Organs
All adolescent rats’ weekly BW was recorded throughout the experiment (Figure 1). After the blood collection, the abdominal cavity of the animals was cut, and the liver and visceral fat pads were excised. The liver and visceral adipose tissue (VAT) were immediately weighed using an analytical weighing scale to calculate the absolute weight (g) and the organosomatic indices of the organs (g/100 g of final BW). The following five main VAT depots were dissected: retroperitoneal, perirenal, perigonadal, mesenteric and inguinal.
Statistical Analysis
The data from the experiments were run through the Shapiro–Wilk normality test. The relative BW comparison between control (standard laboratory diet), HFD + exercise and HFD sex-matched pups across every week was made using the Kruskal–Wallis test and Dunn’s post hoc. The weight of VAT, liver and biochemical estimation was compared between-diet sex-wise and also between-sex diet-wise. Comparisons of the EPM variables between three groups based on sex and diet type were done using mixed-model analysis of variance (ANOVA) and Tukey’s post hoc analysis. Diet versus sex factors influencing the biochemical and structural weight were also compared using mixed-model ANOVA and Tukey’s post hoc analysis. Statistical test was done using GraphPad Prism v.9 software, and significance was considered at p < .05.
Results
Biometrics
A significant rise in BW was observed in HFD + exercise males (F1,3 = 16.9, p = .02) and females (F1,3 = 14.9, p = .02) from 2nd to 5th week of HFD supplementation. We observed a significant increase in BW from 4th to 9th week in HFD-fed male pups, and in female rats, the increase spanned from 6th to 10th week of exposure. As the exercise was started from the 6th week, the change in BW was comparable between the control and HFD + exercise males, but a significant decline (F1,3 = 11.0, p = .01) was seen in HFD + exercise female rats (Figure 2a and 2b). Absolute VAT weight was increased in both HFD + exercise and HFD male (p = .01) and female (p = .0005), while in female pups, organosomatic VAT weight was also more in HFD + exercise (p = .01) and HFD (p < .0001; Figure 2c). HFD female pups (p = .02) and HFD + exercise (p < .0001) group had higher absolute VAT weight compared to their diet-matched males. Lower absolute liver weight (F1,10 = 14.8, p = .003) was observed in control females (p = .0006) compared to males. Both absolute and organosomatic liver weights were increased only in female pups of HFD + exercise (p = .05) and HFD groups (p = .0003; Figure 2d).

Biochemical Evaluation
Plasma glucose in HFD + exercise (F1,19 = 6.3, p = .02) and in HFD (F1,19 = 8.9, p = .003) female rats was significantly higher compared to control, while the males showed no inter-group difference (Figure 3a). Inter-diet (F1,11 = 1,060, p < .0001) and inter-sex (F5,37 = 10.5, p = .007) influence on total plasma cholesterol was significant; the inter-sex difference was more prominent in female pups (Figure 3c). Sex (F5,37 = 11.2, p < .0001) and diet (F1,11 = 947.4, p < .0001) influenced plasma HDL levels, which were higher in HFD + exercise (p = .0006) and HFD female (p < .0001) adolescent rats compared to their control (Figure 3d). Compared to the control, the diet type (F3,28 = 159.2, p < .0001) significantly increased plasma LDL in both males and females (Figure 3e). Plasma VLDL showed no inter-diet or inter-sex statistical significance compared to controls (Figure 3f). The biochemical parameters obtained from the liver homogenate did not show any inter-sex and/or inter-diet differences (Figure 3b–3f).

Behavioural Evaluation
Control female rats showed significantly higher EL (F3,39 = 12.3, p < .0001) compared to the male rats on Day 1 (D1; Figure 4a). EL significantly decreased in HFD + exercise female rats (F2,18 = 6.6) on Day 2 (D2; p = .04) and Day 3 (D3; p = .002) compared to D1. On D1, HFD-fed male rats had longer EL than the control and HFD + exercise group, while HFD-fed female rats had shorter EL than the control diet group. On D2 and D3, the HFD + exercise female rats had shorter EL (p = .006) than control females. On D3, HFD-fed male rats showed decreased NOEX compared to the sex-matched control (p = .02), while HFD female rats had less NOEX and increased NOEN compared to HFD + exercise group (p = .05; Figure 4b and 4c). OA time in HFD-fed female rats was significantly decreased (F5,25 = 5.6) and increased in HFD-fed females on all three EPM days compared to control (p = .04) and HFD + exercise (p = 0.05) group female rats (Figure 4d).
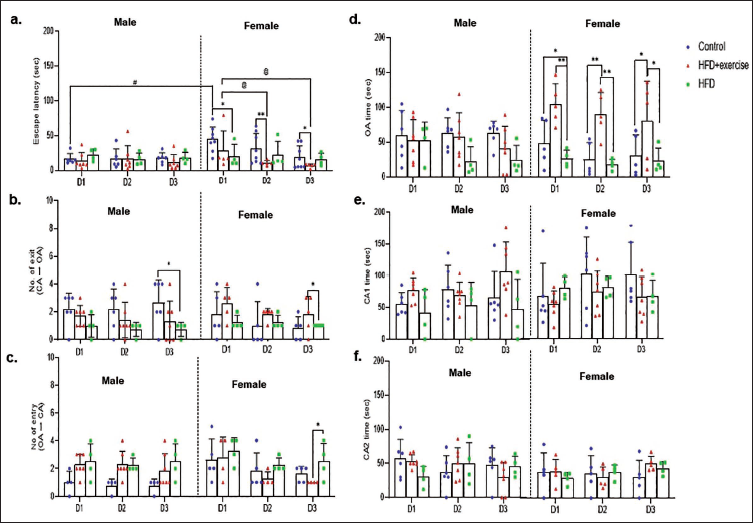
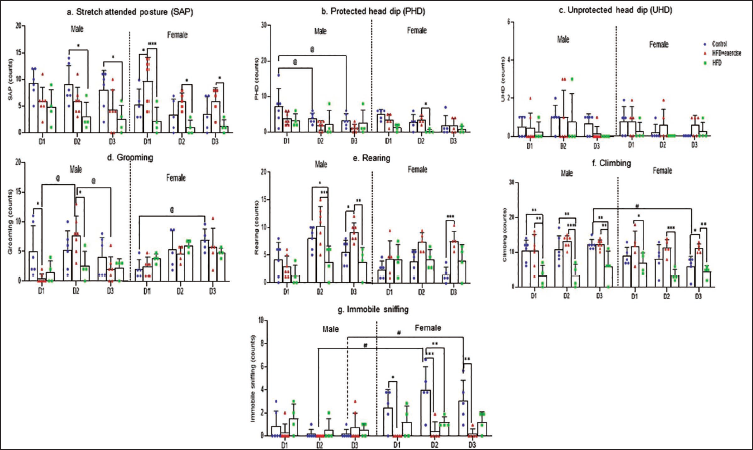
Inter-diet (F1.9, 47.4 = 5.4, p = .009) and intra-sex influence on SAP was observed (F5,25 = 6.7, p = .0004). On D2 and D3, HFD-fed male rats showed decreased SAP compared to sex-matched control, while in female rats on all 3 days, decreased SAP (p = .01) was observed in the HFD group compared to HFD + exercise (Figure 5a). pHD progressively decreased day-wise in control male rats (F2,57 = 9.3, p = .04), while no inter-sex between-diet variation was observed between male and female rats (Figure 5b). In HFD-fed female rats, pHD decreased compared to HFD + exercise on D2 (p = .01). Grooming showed a significant difference across days of recording (F2,57 = 8.4, p < .005) and also between diet and sex (F6,57 = 3.7, p = .004). Control female rats showed increased grooming on the D3 compared to D1; a similar trend was observed in HFD + exercise females, though not significant. HFD + exercise males showed decreased grooming on D1 compared to control (p = .0205). Grooming decreased significantly on D1 in HFD + exercise males compared to control and increased on D2 compared to D1, further decreasing on D3 (p = .0007; Figure 5d). In HFD-fed female rats, increased grooming was observed compared to the control group on D1 (p = .03). HFD-fed male pups showed lowered rearing compared to control and HFD + exercise on D2 (p = .04), while on D3, HFD + exercise females showed increased rearing (p < .001) compared to control (Figure 5e). Both HFD-fed female and male rats showed decreased climbing (Figure 5f) compared to control and HFD + exercise rats (F3,57 = 6.1, p < .005) on all EPM days (p = .0061). Control females had increased IS (Figure 5g) compared to males (p = .008), while inter-diet significantly lowered IS was seen in HFD + exercise females (F3,57 = 6.1, p < .005).
Discussion
Diet is an important environmental factor that affects the developing brain and behaviour and causes biochemical alteration. This study evaluated the effects of a post-weaning HFD and the beneficial effect of aerobic exercise on the morphometric, biochemical and anxiety-like behaviour in adolescent male and female rats.
Biometric Evaluation
Diet-induced BW gain in adolescent male and female rats was observed, where the gain was significantly ‘exercised away’ only in female rats. Boukouvalas et al 1 observed no significant difference in initial and final BW in adolescent male and female rats, and Wang et al 5 reported diet-induced weight gain in the 5-week male rats fed with soybean oil-rich HFD for 10 weeks. Switching to a low-fat diet for 8 weeks and treadmill exercise for 12 weeks reversed adiposity and weight gain in male rats, where exercise showed more promising results.5, 28 Another study showed that 8 weeks of swimming and resistance training were able to improve lipid profile and adiposity in HFD-fed male rats. 17 Taking a clue from these previous reports, we have exposed our rats to a 4-week swimming protocol17, 22 and found that exercise could reduce BW in HFD-treated female adolescent rats only. We have restricted ourselves to a 4-week swimming protocol to investigate the results during the adolescent period of the rats.29–32
In our study, there was no significant increase in both absolute and normalised liver weight in the HFD + exercise males, while in females, both weights increased in the HFD + exercise group, and it further increased in the HFD group. Similar to our study, absolute and normalised liver weight was increased in fish-oil-based HFD-fed rats compared to the controls. 4 Increased intra-abdominal fat pads were observed in lard-based HFD compared to soybean oil-based fat diet. 5 Similar results are observed in our study, too, where VAT increased in the HFD + exercise and HFD group in females and males, but the organosomatic index in males did not show a significant difference. Hence, this signifies that though 4 weeks of aerobic exercise does produce a decline in weight in female rats, this was not associated with the VAT depots. Our findings are consistent with a previous study that did not observe a significant VAT weight or adipocyte area decrease with continuous 8-week exercise in rats provided with HFD. 17
Effect of HFD and Exercise on Biochemical Estimation During Adolescence
We have found increased TC, HDL and LDL in male and female adolescent rats in both the HFD and exercise groups. The LDL values decreased, and HDL increased in the HFD + exercise rats compared to the HFD group; this may be due to the effect of exercise. Our results were similar to some previous reports, with some incongruency, which could be due to the type of fat used, duration and type of exercise protocol, age, and sex-specific.3–5,17,28,33,34 An increased plasma TC and HDL in rats fed with HFD for 12 weeks, followed by another 12 weeks of treadmill exercise, was seen, while no change in plasma LDL, TG and plasma glucose was observed. 28 Plasma TG was significantly higher in sedentary HFD-fed rats compared to sedentary control rats, while swimming for 8 weeks lowered TC and elevated HDL compared to sedentary control male rats, with no effect on plasma TG. 17 In another study, swimming for 8 weeks in male rats did not affect HDL, TC and fasting glucose in HFD-fed rats. 16 It is worth mentioning here that in all these studies, by the end of the exercise protocol, the HFD-fed rats were beyond adolescence. In our study, we have chosen an HFD and exercise protocol within such a time frame that the observation was purely confined to adolescents.30, 32
It has been observed that HFD-fed animals and humans develop dyslipidemia and metabolic syndrome.35, 36 Elevated plasma TG, TC, LDL and reduced HDL compared to control was observed after 4 weeks of HFD feeding in young male rats. 3 In 6-week-old male rats, plasma TG was significantly higher in coconut-oil-based HFD, and it was not significantly increased in lard-based, olive oil-based and fish-oil-based HFD supplementation. In this study, fasting glucose was significantly increased only in olive oil-based HFD-fed rats. 4 Adult female Lewis rats showed increased plasma LDL and decreased HDL compared to control diet-fed rats, 34 while no change in lipid profile and plasma glucose was observed in adult female Sprague-Dawley rats fed with HFD. 33 Fasting glucose and TC showed no significant difference in adolescent male rats supplemented with lard-based and soybean oil-based HFD compared to when these HFD-fed rats were switched to low-fat diets. 5 Another report quotes that HFD during adolescence produces glucose intolerance, increased fat deposition and reproductive changes in male rats, which were partially reversed by 30 days of exercise. 2 Thus, the differences in our observations from these previous reports could either be due to the type of fat used or the effect of exercise or because the biochemistry of adult and adolescent rats may vary in a sex-specific manner.3, 33, 37, 38 However, 4 weeks of aerobic exercise was not enough to bring about favourable changes in all the biochemical parameters except for some parameters in a sex-specific manner. We have observed no significant change in liver lipid profile based on inter-diet comparison. Some studies showed elevated liver TG based on the dietary fat content,4, 39 while others showed no change or reduced liver lipid profile in HFD-fed animals compared to control.3, 40
Effect of HFD and Exercise on Behavioural Assessment During Adolescence
An HFD, along with exercise, influences anxiety-like behaviour in a sexually dimorphic way. Emotionality, as evident from the EPM D1, shows reduced EL in HFD-fed female rats, while habituated learning reduced EL in HFD + exercise female rats only and not male rats compared to control on D2 and D3. In HFD-fed female pups, reduced OA was seen compared to the HFD + exercise group on the third recording day, indicating that even with practice, HFD feeding could not reduce anxiety in both sexes. Similarly, OA time was significantly increased in HFD + exercise females, while reduced OA time was seen in the fed group. HFD feeding in female rats showed reduced time spent in CA2, indicating less exploratory behaviour. We observed reduced sniffing, increased climbing and rearing on D3 in HFD + exercise female rats compared to males and their HFD counterparts, indicating that HFD coupled with exercise made females more efficient in decision-making and less anxious. Changes in SAP and head dip in HFD + exercise females also reflect that the female adolescents were less anxious than male adolescents. Remarkably, HFD feeding in both sexes caused less SAP and head dip, indicating reduced exploration. Previously, a 20-week HFD exposure to adolescent male mice enhanced anxious behaviour. 6 Moreover, it has been shown previously that EPM evaluation of anxiety is age and sex dependant, 41 whereas, generally, females were less fearful. 42 Similar to our findings, adult female rats exposed to 10 weeks of HFD showed heightened anxiety. 43
Though many studies have shown that exercise can ease the detrimental metabolic effects of HFD, the behavioural outcome from rodent studies where interaction between HFD and exercise is contentious.9, 44, 45 We have found that aerobic exercise coupled with HFD could not make the adolescent male rats any less anxious, unlike the females. Voluntary wheel-running exercises attenuated the anxiety-like behaviour in HFD-fed male mice,44, 45 while similar to our study, the HFD-induced adverse behavioural effects in adult male mice could not be ‘exercised away’ in another study. 9 Rats, being natural swimmers, can swim without producing strong resistance,46, 47 and swimming has been used as an effective exercise protocol to alleviate many diseases in rodents. 47 Moreover, it has been previously reported that moderate-intensity swimming effectively increased the expression of interleukin 10 in the VAT, controlling BW and reducing the epididymal and retroperitoneal adipocyte areas, producing positive effects on lipid profile. 48
Long-term (8–12 weeks) exposure to HFD disrupts the hippocampal and amygdala-dependent memory in juvenile male rats, which is glucocorticoid-mediated. However, HFD exposure in adult male rats enhanced long-term potentiation and object location memory. 11 Moreover, neurocognitive derangement was observed in adolescent male rats, which were restored with control diet supplementation during adulthood. 7 Adolescence is a vulnerable time for long-lasting dysfunction of cortical and hippocampal activity due to HFD.12, 13
Conclusion
In rats, 10 weeks of HFD combined with 4 weeks of exercise cause hyperglycaemia and dyslipidaemia during their early childhood and adolescence. Despite a reduction in BW, exercising for 4 weeks did not reduce VAT mass. Adolescent rats exhibit HFD-induced anxiety-like behaviour, which may have been ‘exercised away’ in females but not males. As a result, HFD has sex-specific behavioural effects, with females responding positively.49, 50 As a result, additional research into the oestrous cycle would be beneficial in corroborating our preliminary findings.
Footnotes
Acknowledgement
The authors want to acknowledge the rats for sacrificing their lives.
Author Contribution
Archana Gaur conceptualised the study, conducted the investigation and formal analysis, developed the methodology, managed project administration, provided supervision. Trina Sengupta curated the data, conducted formal analysis, investigated, conducted software analysis, and performed validation. Sutirtha Ghosh conducted the investigation and carried out a formal analysis. Prasunpriya Nayak curated the data, conducted the investigation, managed project administration, arranged the resources, conducted software analysis and provided supervision. Nachimuthu Maithilikarpagaselvi conducted the investigation and arranged the resources.
Authors’ Notes
The work was conducted at the All India Institute of Medical Sciences, Jodhpur, Rajasthan. The first, second and third authors were affiliated to this institute but shifted later.
Statement of Ethics
All experimental conditions and procedures were performed in compliance with the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA) guidelines, applicable at the time of conducting the experiments. The study was conducted at the All India Institute of Medical Sciences, Jodhpur, under Mou with Jai Narain Vyas University, Jodhpur with the ethical clearance number JNVU/IAEC/2020/09.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest concerning the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.
Patient Consent
The study does not involve patients.