
Abstract
Background
Acute heart rate variability (HRV) is used for assessment of functional autonomic profile in awake resting state in humans subjects. Evaluation of 24-h variations in the sympatho-vagal balance across sleep stages through HRV are valuable to understand the basis of various lifestyles diseases, however, such studies are uncommon.
Purpose
The present study modelled the sleep-wake associated cardiac autonomic changes in freely moving female Wistar rats across light and dark phase using HRV as a tool.
Methods
Sleep states (non-rapid eye movement [NREM] and rapid eye movement [REM] sleep) were estimated through electroencephalogram (EEG) and electromyogram (EMG), while the ECG signal was acquired simultaneously for HRV analysis. To understand circadian dynamics, HRV was analysed for noise-free distinct and transitional state segments of sleep–wakefulness.
Results
In time domain parameters, RR intervals during NREM sleep were longer in comparison to wake and REM sleep. In the frequency domain, reductions in LF and an increase in HF power during NREM sleep were evident compared to other states. RR interval during wakefulness in the dark phase was significantly decreased compared to the light phase. The RR interval was the longest during the early phase of the light period (6–10
Conclusion
Parasympathetic predominance during NREM sleep was similar to a human having a monophasic pattern, while a higher sympathetic tone during the majority of transitional states in rats, which are polyphasic in nature, indicated risk of dysregulated sympatho-vagal balance with fragmented sleep. Further, the highest parasympathetic activity during the early light phase and the highest sympathetic drive during the dark phase may serve as an adaptive mechanism for survival in nocturnal life.
Keywords
Introduction
Sleep is an integral component of life wherein wide variations in its duration, pattern and adaptive dynamics during the life span are reported in various species, diurnal or nocturnal, as an adaptation to survive and sustain in nature.1, 2 Rodents, which are one of the commonly used experimental animals, display two distinct sleep states similar to humans, that is, non-rapid eye movement (NREM) and rapid eye movement (REM) sleep, each having its own characteristic features, regulatory mechanisms and functions.1, 3, 4 Distinct polyphasic sleep in rats with pronounced nocturnal variations, unlike humans, which are diurnal by nature but are largely monophasic sleep except during neonatal life, provides an interesting perspective to understand the interactions of the neural mechanism of sleep with the autonomic nervous system (ANS) and other systems of the body.1, 5–10 The gender specific variations in sleep, especially in females, during the menstrual cycle (Oestrous cycle in rats), pregnancy and menopause, provide another dimension of the effects of sleep loss on heart-brain interaction during ageing to understand physical, emotional and mental health issues that are on the rise in the current lifestyle.11–16 Sleep is gaining more importance now after the COVID-19 epidemic for attaining good health and well-being in all domains of clinical and traditional practices, like Indian Ayurveda, etc.14, 17–20
Simultaneous evaluation of sleep and autonomic function tests is not commonly carried out due to practical difficulty in signal acquisition and overnight stay in a sleep lab. 16 In a clinical setup, heart rate variability (HRV) assessment that captures beat-to-beat variation in heart rate during a quiet resting state is a non-invasive and widely used diagnostic tool to evaluate the sympathetic-parasympathetic (autonomic) balance.14, 21–24 Variations in HRV are measured using time and frequency domains as each parameter provide an indication of autonomic health (Table 1). Even though this is generally carried out for a couple of minutes, long-term HR recording (24 h) can provide extensive comparison of the HRV parameters during different states like rest, wake, sleep, etc. 25
HRV Parameters and Their Interpretation as per Task Force of the European Society of Cardiology and the North American Society of Pacing Electrophysiology (1996). 25

The variability in cardiac rhythms during different stages of sleep offers a unique perspective into the interaction between the sympathetic and parasympathetic parts of the ANS, but such detailed studies are less common.26–33
With a growing need to study the developmental changes as a result of insomnia on cardiovascular diseases, effects of several surging environmental factors, including global warming, pollution, etc., a suitable animal model is required to assess the state-dependent changes in the HRV, especially in females, as such studies are less common. The aim of this study was to develop a rat model to assess dynamic changes in the cardiac ANS activity during sleep–wakefulness (S–W) by using HRV as a tool, and also to address changes in circadian pattern during light and dark phases in the key HRV parameters in female Wistar rats.
Methods
Animals
Experiments were conducted on female adult Wistar rats housed in a polystyrene cage with a 12:12 h light-dark phase (light on at 6
Procedure of Implantation of Electrodes for Recording S–W and ECG Signals
To assess S–W in rats, the EEG and EMG electrodes were implanted under anaesthesia (Ketamine 50 mg/kg and Xylazine 5 mg/kg body weight, im) as described previously.1, 34 In brief, two miniature screw electrodes (1 mm shaft diameter) and another one as a reference electrode were implanted bilaterally on the skull above the parietal cortex for recording EEG, along with loop electrodes in the nuchal muscle for acquiring EMG. For ECG, insulated micro wires (0.1 mm diameter) were implanted subcutaneously under the forearms of the animal, and the reference electrode was placed on the dorsal side of the rat.35, 36 The electrodes were connected to an IC socket, and the whole assembly was fixed on the skull using dental cement.1, 17 Analgesics and antibiotics were given to the animal during the post-surgical recovery period.
Recording and Analysis of S–W
The S–W and ECG signals of a conscious, freely moving rat were acquired simultaneously using the data acquisition system Biopac MP150 (Biopac System, CA, USA) with a sampling rate of 1,000 Hz. The signals were amplified (×5,000), filtered (EEG: 0.1–35 Hz; EMG: 1–500 Hz), and digitised at 1 kHz. The S–W recordings of 24 h were scored manually (offline), taking epochs of 30 sec durations. The S–W stages were classified into W, NREM, and REM sleep.1, 37
Analysis of ECG Signals for S–W State-dependent HRV Analysis
HRV analysis was carried out using the software Kubios Scientific version 4.1.1. The standardisation of frequency filters for HRV analysis was performed based on the Nyquist-Shannon Sampling Theorem and previous reports38, 39 (Table 2). In this study, frequency filters chosen were LF: 0.25–0.80 Hz, and the HF: 0.80–2.50 Hz. As the HR in rats is higher than in humans, the optimisation of epoch lengths of W, NREM or REM sleep (30–120 s) was used due to shorter cycle duration, particularly the NREM and REM sleep. In adult rodents, the LF and HF bands are adjusted to account for their faster heart rates and respiratory cycles, with standard cut-off frequencies for LF between 0.20 Hz and 0.80 Hz and for HF between 3 Hz and 6 Hz.40, 41
Cut-off Frequency Range Used in Various HRV Studies in the Adult Rodent Model.
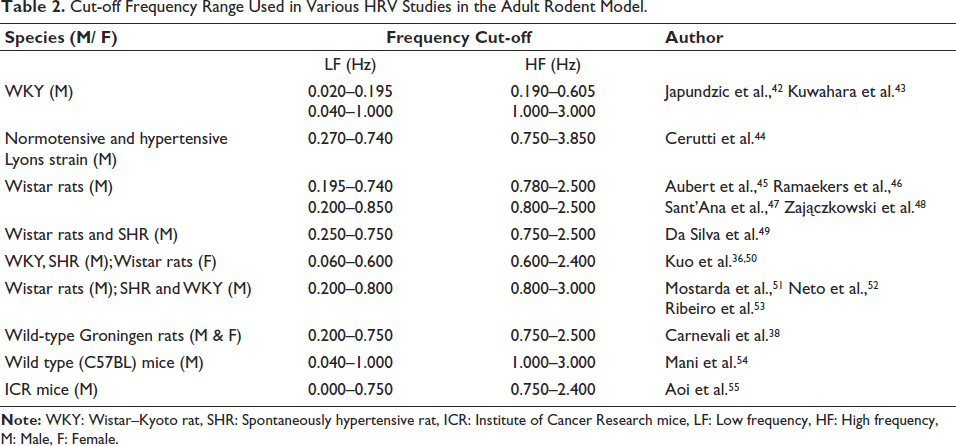
To standardise the epoch length for HRV analysis, 120-second epochs in NREM sleep were analysed first and then fragmented into 60- and 30-s epochs and the variations observed in the results. Since there were no significant changes observed in these three duration epochs, 30 s epoch length was taken for further comparison in this study (Table 2). For HRV analysis across all sleep-wake states, the time domain parameters included HR, RR interval, SDNN (standard deviation of normal-to-normal beat interval), and rMSSD (root mean square of standard deviation). In the frequency domain, LF power, LF nu, HF power, HF nu, and the LF/HF ratio were analysed.
To investigate HRV pattern across S–W stages, 72-h polysomnographic data were segmented into 30-second epochs and classified into W, NREM and REM sleep based on standard scoring criteria 9 and HR traces were labelled for each state of S–W. Considering short sleep S–W cycles, an illustration of changes occurring during the transitional states, that is, NREM stage occurring immediately preceding or following another state, was carried out separately. S1 and S2 fragments were subdivided based on visual inspection of the amplitude of the wave during NREM sleep. Thus, for transitional states for S1, four conditions were taken: (a) W-S1, where W preceded the S1 state, (b) S1-S2, S1 to S2, that is, where S1 preceded the S2 state, (c) REM-S1, where REM sleep preceded the S1 state and (d) S1-REM, where S1 preceded REM sleep.
To assess the overall day and night pattern, the averaging of HRV parameters was taken for W, NREM and REM sleep epochs. The 24-h sleep–wakefulness and ECG recordings were segmented into six 4-hour time bins: 6
Statistical Analysis
Statistical analysis was carried out using IBM SPSS Statistics 30.0.0. Comparisons of time spent in W, NREM and REM sleep (% time in each state) during day versus night were done by using a t-test.
For comparison of the statistical differences between 30s, 60s, and 120s epoch length of NREM sleep, the normality of the data was checked by the Shapiro–Wilk test. The NREM sleep stage was selected for epoch-length comparison to have a more stable stage with the least artefact. Since the data was normally distributed, the difference of the results obtained from three different epoch durations (30s (n = 40), 60s (n = 20), and 120s (n = 10)) was examined with Welch’s ANOVA. Eta squared (η 2 ) was calculated to determine the effect size to assess the magnitude of an observed effect. For threshold, η 2 = 0.01 (small effect), η 2 = 0.06 (medium effect) and η 2 = 0.14 (large effect) were considered.
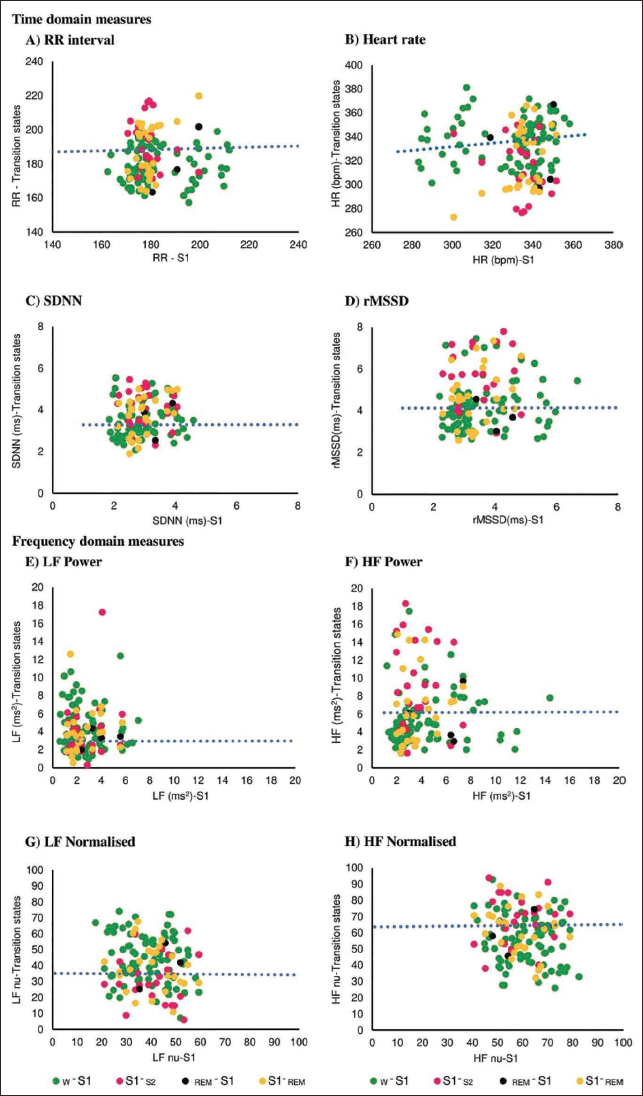
For HRV parameters during transitional states number of epochs taken for analysis are given in brackets, that is, W-S1 (W to S1, n = 84), S1-S2 (S1 to S2 sleep, n = 23), REM-S1 (REM to S1, n = 4), and S1-REM (S1 to REM, n = 27), and stable S1 stage (n = 357). The difference between the HRV during stable S1 sleep stage (n = 357) and these four transitional states was illustrated in a scatter plot with a linear regression trend line fitted using the least-squares method. The Pearson correlation coefficient was used to assess the correlation between variables.
Subsequently, the day and night changes in HRV parameters for clear noise-free epochs of W (day n = 18; night n = 17), NREM (S1) (day n = 357; night n = 200) and REM sleep (day n = 156; night n = 100) were performed by one-way ANOVA. Differences were considered statistically significant at p < .05 in all analyses.
Finally, for the comparison of circadian pattern in the state-dependent changes in HRV parameters, the 24 h data were divided into three bins of 4 h in the light phase, that is, 6
Results
Profile of Sleep–Wakefulness States for Three Consecutive Days
The sleep staging was done for three days’ data using EEG and EMG signals. The percentage of different sleep stages was analysed and tabulated in 24 h, and the difference in day and night S–W was also incorporated (Table 3).
Percentage of Time in Stages of S–W in Normal Rat: Twenty-four Hour and Day-night Profile.
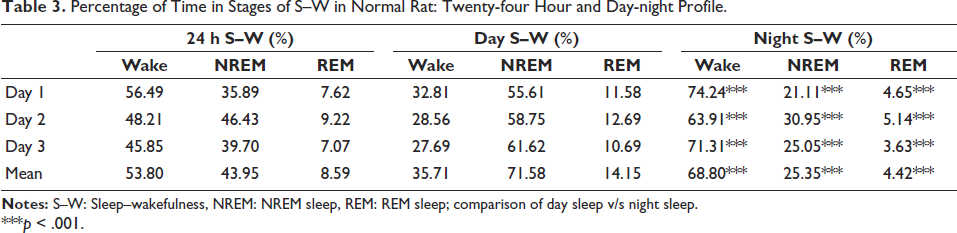
***p < .001.
Standardisation of Epoch Length (S–W) for HRV Analysis
The results of the comparison between 30s (n = 40), 60s (n = 20) and 120s (n = 10) data are given in Table 4. The Welch’s-ANOVA test results showed that no significant differences were found in three data sets (p < .05), as shown in Table 4. The effect size was considered small (η 2 = 0.01–0.06) for all HRV parameters in 60s and 120s. The 95% confidence interval (CI) for the difference between means showed that the difference is not statistically significant (95% CI includes 0). Generally, REM sleep epochs are in the range of a few seconds to 2–3 minutes. 9
Comparison Between HRV Parameters (Time and Frequency Domain) Obtained from Three Different Epoch Lengths of NREM Sleep.
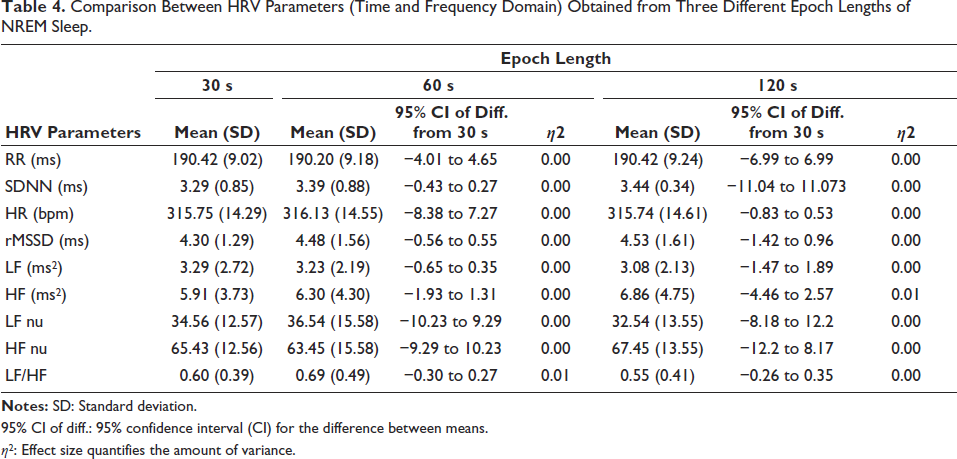
95% CI of diff.: 95% confidence interval (CI) for the difference between means.
η 2 : Effect size quantifies the amount of variance.
Comparison of HRV Parameters in Time & Frequency Domains for the Transitional States
Comparison of HRV parameters of stable S1 state (with classical sympatho-vagal profile) and the HRV profile of the transitional stages is shown in Figure 1. The scatter plot showed a wide distribution of data points in all four S1 states (a) W-S1, (b) S1-S2, (c) REM-S1 and (d) S1-REM for HRV parameters with a low overall correlation score.
Comparison of HRV Parameters Across W, NREM and REM Sleep During Day and Night (Averaged)
The RR interval was higher during NREM-S1 sleep compared to both W and REM sleep during the day and night. Furthermore, RR intervals were greater during daytime compared to nighttime W and REM sleep than nighttime. Among all stages, NREM sleep showed the lowest HR compared to W and REM sleep during both night and day. SDNN values for W and REM sleep were higher than the NREM-S1 state during the day and nighttime period. State-dependent changes in rMSSD were only found during nighttime.
During NREM sleep, LF power and LF nu were lower compared to W and REM sleep during the day. The differences between NREM and REM sleep were observed during the night phase only. HF nu was higher during NREM sleep compared to W and REM sleep at daytime. Both day and nighttime, the LF/HF ratios were lower during NREM sleep compared to REM sleep.
LF power and HF power showed differences only in REM sleep for day and night comparisons. However, LF nu, HF nu and LF/HF ratio showed significant changes between the daytime and nighttime W (Table 5).
Comparison of HRV Parameters Across W, NREM and REM Sleep During Day and Night.
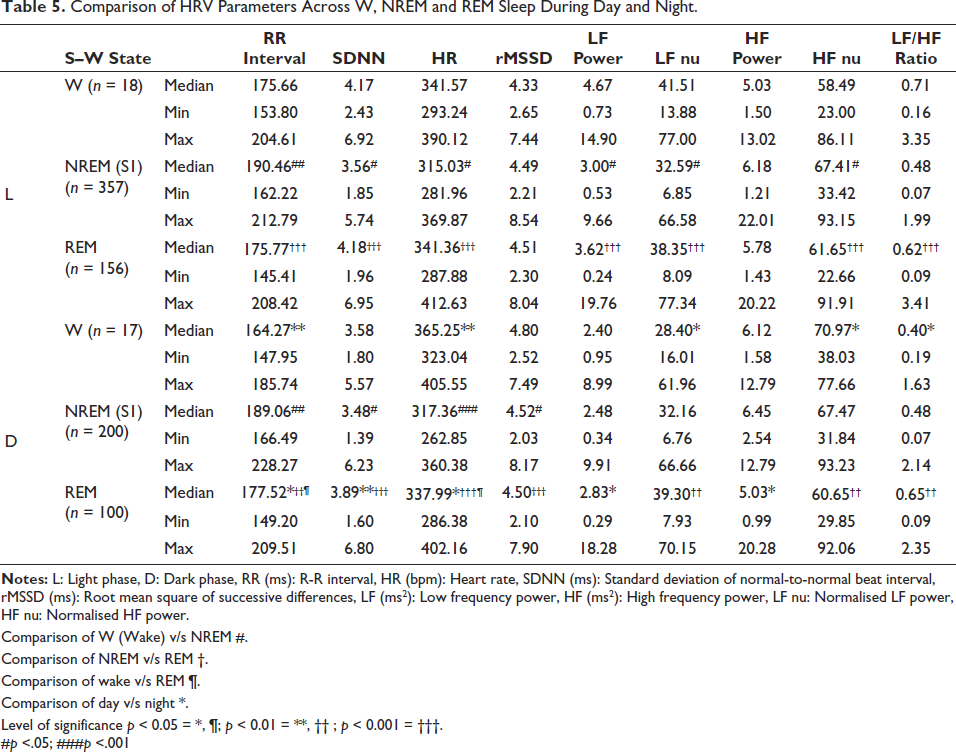
Comparison of W (Wake) v/s NREM #.
Comparison of NREM v/s REM †.
Comparison of wake v/s REM ¶.
Comparison of day v/s night *.
Level of significance p < 0.05 = *, ¶; p < 0.01 = **, †† ; p < 0.001 = †††.
#p <.05; ###p <.001
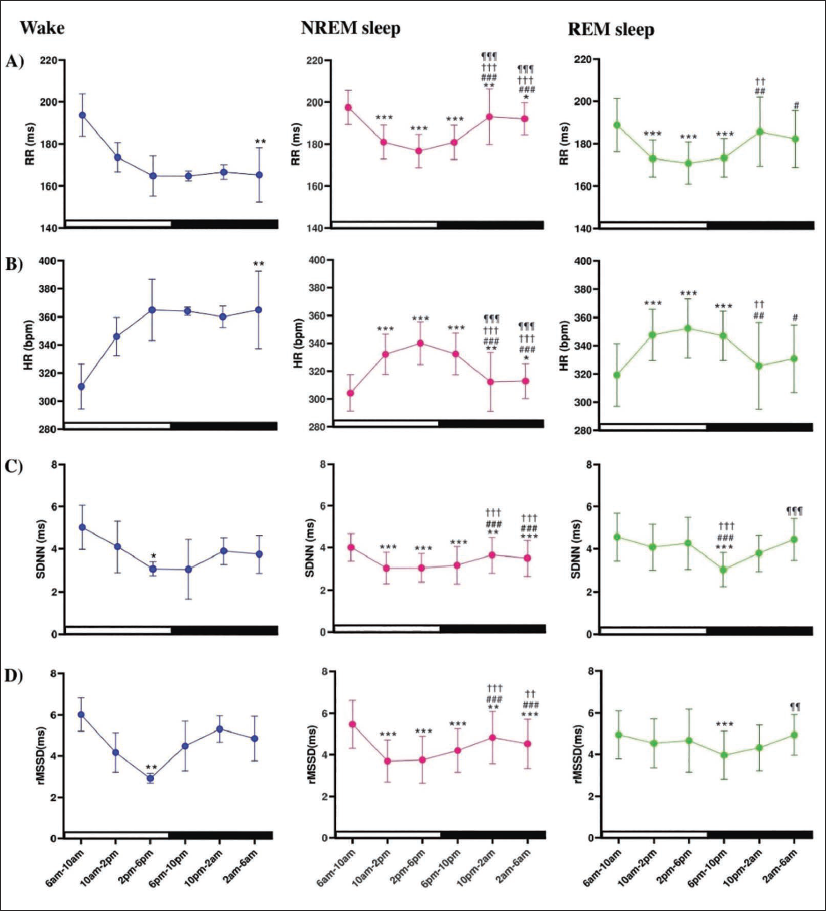
Circadian Pattern in HRV Parameters Across Light and Dark Phases in 24-h S–W
For W, RR intervals gradually decreased from 6
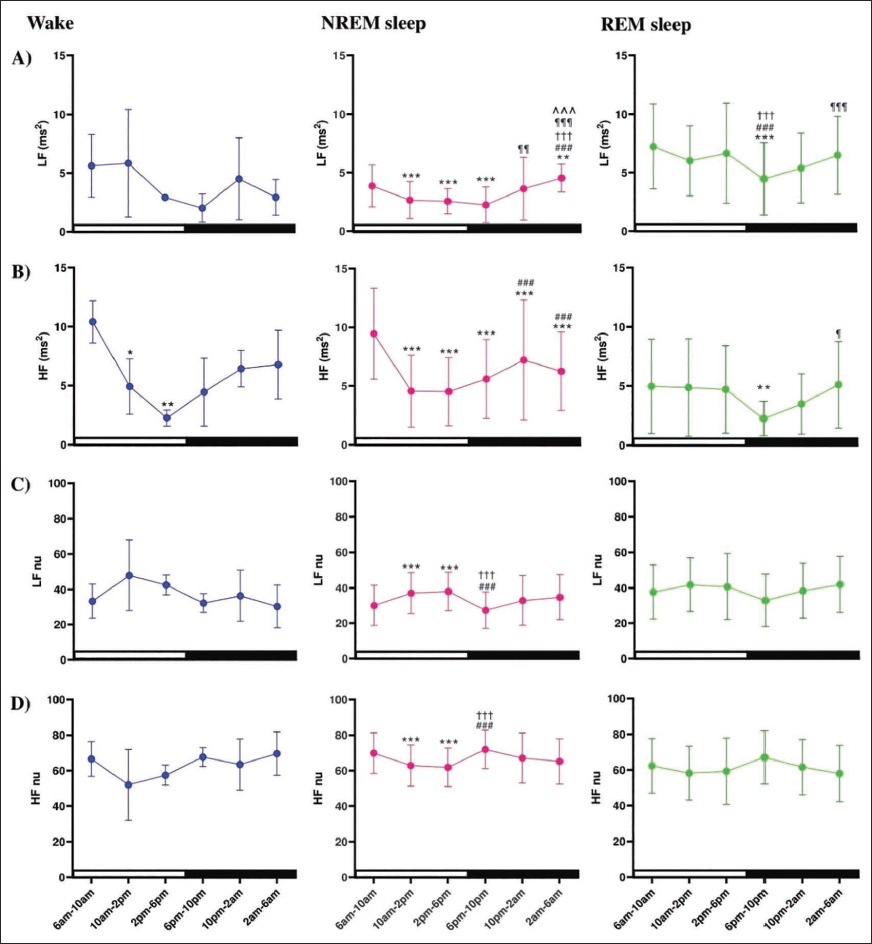
In NREM sleep, LF power decreased during the daytime from 6
Discussion
The present study described the dynamic changes in the sympatho-vagal balance during different stages of S–W for day and nighttime in a control female rat. The percentages of time spent in each stage of S–W of the animal confirmed that it had normal sleep, indicating its normal health state. 9
HRV results obtained from the data of transitional states indicated wide variation in the time and frequency domain parameters, reflecting a dynamic sympatho-vagal fluctuation with change in state. The lower RR interval and higher HR in the epochs in the transition states, especially for W-S1, REM-S1 and S1-REM, compared to stable S1, may indicate the higher sympathetic activity at those times. It may also indicate a delay in activation of the parasympathetic branch during the transition between different stages. This result is in line with another study where the high RR interval was reported during an NREM state epoch that transitioned from W state in Wistar Kyoto rats. 56 This delayed processing may be attributed to the time required for the neural mechanisms regulating sympatho-vagal balance to fully adapt to the transition into a specific sleep stage.
Studies indicate that ageing, sleep patterns, and sympatho-vagal balance are interrelated, particularly in females. Age-related declines in sleep quality, along with acute or chronic sleep disruption, have been associated with heightened sympathetic activity in women. 57 The observed sympathetic predominance during transitional states in our study suggests that fragmented sleep or reduced sleep bout duration may disrupt ANS balance.
The longer RR interval during daytime W state compared to night (high HR during night W) suggested a deeper parasympathetic influence of the daytime sleep in this species. The increase in RR interval during NREM sleep than during W 51 and the reduced HR during NREM sleep than during the W state is consistent with the previous reports. 59 During REM sleep, HR was higher than during NREM sleep in our study. However, higher variability in HR is reported during REM sleep in both humans and rats.50, 58 At certain points, the HR can be higher in NREM sleep than in REM sleep, but the average HR did not change significantly. 59 The circadian influence on RR interval and HR appears most prominent during W. Sleep stages (NREM and REM sleep) showed a more stable autonomic profile, regardless of time of day. In a previous report, the HR displayed a circadian variation in SHR and WKY rats, with the lowest values during the daytime and the highest during the night, without differentiating the sleep stages. 60
In humans, observation of lowered HR during NREM sleep as compared with W suggests an increase in vagal activity during NREM sleep.61–63 HR during REM sleep is highly variable due to changes in cardiovascular responses during this state.64, 65 Sleep-related changes in HR appear to be mediated predominantly by changes in ANS, wherein the lowest HR during deep sleep (N3–NREM sleep) in humans indicate predominant parasympathetic tone. 66 Studies investigating the effect of sleep stages on autonomic control have reported circadian regulation of the HR and vagal activity.26, 63 A similar trend of reduction in HR during sleep compared to W is also reported in domestic dogs. 67
SDNN reflects overall HRV, both sympathetic and parasympathetic influences and is especially sensitive to longer recordings. 25 A decrease in SDNN during REM sleep at night might suggest more stable or elevated sympathetic tone. During nighttime, a slight reduction (even though non-significant) in SDNN for the wake state aligns with increased HR, suggesting a sympathetic dominance. Stable rMSSD across stages and time (Day vs. Night) may suggest that short-term HRV linked to parasympathetic activity remains consistent regardless of sleep stage and circadian timing (Day vs. Night). rMSSD primarily reflects parasympathetic activity and is less influenced by sympathetic tone or long-term trends. The increase in rMSSD during NREM sleep at night suggests a stronger parasympathetic activation during nighttime NREM sleep. The lack of difference during daytime might be due to a higher baseline sympathetic tone during the day, masking subtle parasympathetic shifts. The lack of variability in rMSSD might indicate limited sensitivity of rMSSD to subtle circadian or sleep-stage-specific autonomic fluctuations in rodent species.
Decreased LF and HF power on REM sleep during night but without any change in wake or NREM stages during day and night, indicates that REM sleep is a complex state with fluctuating autonomic tone. The reduced LF power in REM compared to W (only during daytime) corroborates another study in WKY and SD rats, where the LF significantly decreased during NREM sleep compared to W, with partial reversals during REM sleep. 50
The drop in LF nu during NREM sleep compared to W/REM sleep at daytime reflects the sympathetic withdrawal or parasympathetic predominance. No reduction in LF nu from W to NREM sleep at night and increased HF nu during the NREM sleep at daytime confirms the predominance of parasympathetic tone at daytime sleep. The low LF nu during nighttime wake than day is conflicting. This may suggest that circadian influences may dampen sympathetic drive in rats even during active wake states in the dark phase. This may be due to (a) the comparatively longer wake period, probably gearing for a more relaxed wake during night than day, or (b) the higher homeostatic drive for sleep during the night and (c) that may cause an increased parasympathetic drive during the quiet wakefulness.
No significant differences in LF nu during NREM or REM sleep between light and dark phases, which is in contrast to time domain measures (high sympathetic activity in dark phase), may indicate that the frequency domain measures are less sensitive to a broad time frame (12 h) in an animal with polyphasic sleep, like a rat. A comprehensive understanding of the circadian pattern of HRV may be helpful in understanding the changes occurring in fragmented sleep, especially obstructive apnoea in humans.
In humans, during NREM sleep, the HF component increases while the LF component decreases, indicating enhanced parasympathetic activity and reduced sympathetic activity during this sleep stage. 68 In contrast, compared to NREM sleep, REM sleep is characterised by a higher LF, but a lower HF component. During REM sleep, LF values approximate W levels while remaining distinctly high, indicating a higher sympathetic activity in the REM state.68, 69 Autonomic regulation during sleep highlights a parasympathetic dominance in NREM and sympathetic resurgence in REM sleep, with circadian inputs also contributing to cardiac regulation, as evidenced by changes in HR over the sleep period. The HF pattern in dogs during sleep also confirms the predominance of parasympathetic tone during NREM sleep. 70
The sleep pattern of rats is influenced by both circadian and ultradian rhythms. In a 12-h light-dark phase, rats exhibited clear 24-h circadian rhythms in W, NREM, and REM sleep. Unlike humans, rats exhibit a polyphasic sleep pattern, meaning they sleep multiple times throughout a 24-h period rather than having one long night’s sleep like humans.9, 71 Rats are less active and sleep more during the day. 9 The result obtained in our study showed the lowest HR in all states during the initial bin of the light phase, which gradually increased and peaked during the last bin of the light phase, which is similar to previous reports.50, 72 Similar changes in HR during day and night are reported without monitoring sleep states. 60 The RR interval and HR patterns suggest the strongest parasympathetic activity during early morning W and NREM sleep, particularly during 1st bin of the light phase.
Continuous 24-h ECG recordings from healthy human adults have demonstrated increased RR intervals at night, which is known as nocturnal bradycardia. In summary, HR slows down at night, and HRV shows circadian variations influenced by changes in HR and sympathetic tone. 73 In a previous study, higher sympathetic tone was reported during the dark phase than the light phase in male Wistar rats, but this study did not specify the sleep or wake-dependent changes in the HRV parameters, as the sleep was not recorded electrophysiologically. 74
Decreased LF power in the NREM sleep during the light phase suggested a decline in sympathetic modulation, but increased power during the dark phase, particularly between 2
In young healthy adults (diurnal), higher HRV during the day and lower at night indicate a robust circadian rhythm with increased parasympathetic activity at night. 76 Since rats are a nocturnal animal, one would expect an increased sympathetic drive during the night and predominance of parasympathetic drive during the daytime. However, due to the polyphasic nature of S–W in rats, probable adaptive variations in the sympathetic and parasympathetic influences are depicted in our study.
Conclusion and Limitations
This study described distinct changes in HRV parameters between different states of S–W and also showed the changes observed during dark-light phases. Predominant parasympathetic influence during the NREM sleep compared to W and REM sleep in rats was similar to humans. Further, a predominant parasympathetic drive during the late dark phase and early light phase in rats corroborated adaptive changes due to their nocturnal life.
This is the first study to describe S–W state-dependent changes in HRV, that is, in time-domain and frequency-domain parameters during light and dark phases to understand dynamic changes in the autonomic balance in female rats. In this study, we also standardised the epoch length and frequency cut-off in adult rats. Though the overall pattern of sympatho-vagal balance is similar in humans and rats, subtle changes are observed in rats due to differences in their sleep patterns. In the normal female rats, changes in sleep across three consecutive days may be attributed to hormonal fluctuations associated with the oestrous cycle. 9
The observed sympathetic predominance during transitional sleep states highlights a potential biomarker for autonomic dysregulation associated with sleep fragmentation, particularly in females. This finding may aid in the early identification of individuals at risk for cardiovascular or stress-related disorders linked to poor sleep quality. Importantly, the use of rats as an experimental model is well justified, as their natural sleep architecture is characterised by short sleep bouts, which are analogous to the fragmented sleep patterns commonly seen in humans with sleep apnoea, sleep fragmentation during pregnancy, PCOS, hypertension, age-related sleep disorders in females, and many other sleep disorders and comorbid conditions. The Insights from our study will contribute to understanding sleep and cardiac autonomic physiology, the mechanisms underlying sleep-related cardiac regulation and its disruption in disease states.
A study could be done on a larger number of rats so that oestrous cycle-associated changes could be deciphered and a better understanding of hormonal influences on cardiac autonomic regulation could be obtained.
Footnotes
Acknowledgements
The authors acknowledge SCTIMST, Trivandrum, an Institute of National Importance under the Department of Science and Technology, Government of India, for providing the infrastructure and facilities for conducting this research study.
Authors’ Contribution
AVA: Data analysis, methodology, writing and reviewing the draft, approved the final manuscript.
NN: Conceptualisation, interpretation of results, writing and reviewing the draft, supervision, approved the final manuscript.
KKG (corresponding author): Conceptualisation, data curation, interpretation of results, writing and reviewing the draft, supervision, approved the final manuscript
Data Availability Statement
The data for the current study can be made available from the corresponding author upon a reasonable request.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: The author (KKG) utilised the Learning Resource Allowance from her Institute for the publication of this article. Contingency grant and Junior and Senior Research Fellowship to AVA from the University Grant Commotion (UGC), India (UGC Ref No: 908/CSIR-UGC NET Dec 2017) are acknowledged.
Patient Consent
Not applicable.
Statement of Ethics
This study was approved by the Institutional Animal Ethics Committee of the Sree Chitra Tirunal Institute for Medical Sciences and Technology, Trivandrum, Kerala (SCT/IAEC-306/January 2019/100).