
Abstract
Background and Purpose
Plumbagin (PLB), a natural compound derived from Plumbago zeylanica L., exhibits potent antioxidant, anti-inflammatory, antibacterial, and anticancer properties. However, its role in modulating periodontitis via the phosphatidylinositol 3-kinase/protein kinase B (PI3K/AKT) pathway and autophagy remains unclear. This study explores PLB’s therapeutic potential in periodontitis, focusing on these mechanisms.
Materials and Methods
In vitro, Cell Counting Kit-8 was used to examine cell viability, and LysoTracker Red was used to assess the degree of autophagy activation. The effects of PLB on periodontitis and the PI3K/AKT signaling pathway in vitro were detected by real-time quantitative polymerase chain reaction and Western blot. In vivo, we established rat models of periodontitis to validate the efficacy of PLB.
Results
PLB exhibited a dose-related suppressive influence on periodontitis progression. Notably, PLB significantly downregulated the components of the PI3K/AKT pathway and modulated autophagy. In rat models, PLB treatment effectively alleviated periodontal inflammation and alveolar bone resorption.
Conclusion
PLB exhibits anti-inflammatory properties in periodontitis by suppressing the PI3K/AKT pathway and enhancing autophagy. Our results offer a scientific basis for using PLB in the treatment of periodontitis, emphasizing its promise as an innovative remedy for this dental condition.
Introduction
Periodontitis is a persistent inflammatory disease that impacts the gingiva and surrounding periodontal structures (Kinane et al., 2017; Kumar, 2021). Its pathological process involves the intricate interactions between host immune responses and oral microbes, primarily initiated by the accumulation of dental plaque (Sedghi et al., 2021). Dental plaque, a biofilm composed of diverse microbial populations embedded in an exopolysaccharide-rich matrix, adheres to tooth surfaces and plays a critical role in disease progression (Belstrom et al., 2021; Karygianni et al., 2020). Among the pathogens involved, Porphyromonas gingivalis (P. gingivalis) is the principal pathogenic microorganism, which plays a pivotal role in the development of the disease. It contributes to the formation of plaque biofilm by secreting virulence factors, such as lipopolysaccharides (LPS), which trigger host inflammatory responses and damage periodontal supporting structures (Reyes, 2021).
Autophagy, an intracellular degradation system, is crucial for preserving the stability of cells by eliminating damaged cytoplasmic components such as organelles, proteins, and pathogens in lysosomes (Boya et al., 2013; Filomeni et al., 2015). Several studies have revealed that macrophages can eliminate P. gingivalis via autophagy, thereby mitigating excessive inflammatory responses (Belanger et al., 2006; Park et al., 2017). Furthermore, phosphatidylinositol 3-kinase/protein kinase B (PI3K/AKT) is a canonical inflammatory signaling pathway that is frequently activated during the progression of periodontitis. It plays a pivotal role in osteoblast proliferation and apoptosis, and influences the activity of downstream effectors through cytokine secretion (Liu et al., 2019; Wu et al., 2013).
Plumbagin (PLB), a quinone compound derived from Plumbago zeylanica L., possesses beneficial biological effects, such as its antioxidant, anti-inflammatory, antibacterial, and anticancer properties (Padhye et al., 2012). PLB can maintain cartilage homeostasis and inhibit inflammatory responses through the downregulation of pro-inflammatory cytokines (Shu et al., 2022). Despite these promising findings, the potential of PLB to modulate periodontitis through the PI3K/AKT pathway and autophagy remains largely unexplored. Therefore, the objective of this research is to explore the impact of PLB on periodontitis, particularly focusing on how it impacts the PI3K/AKT pathway and autophagy.
Materials and Methods
Ethics
All experimental procedures involving animals were approved by the Ethical Review Committee of Jiangsu Kebiao Medical Technology Group Co., Ltd., and conducted in accordance with institutional and national guidelines for the care and use of laboratory animals. Efforts were made to minimize animal suffering, including the use of anesthesia and humane euthanasia protocols.
For the in vitro studies, the RAW264.7 cell line was obtained from the American Type Culture Collection, and no human or primary animal tissues were used, thus requiring no additional ethical approval. However, all cell culture experiments followed standard biosafety and ethical research practices.
Study Design
The present study utilized an experimental preclinical design involving both in vitro and in vivo investigations to evaluate the effects of PLB on periodontitis. For the in vitro component, mouse monocyte-macrophage RAW264.7 cells were cultured under standard conditions and stimulated with LPS to establish an inflammatory model of periodontitis. Cells were then treated with varying concentrations of PLB (0–20 µg/mL) to assess cytotoxicity, anti-inflammatory effects, and modulation of the PI3K/AKT pathway and autophagy processes. Mechanistic studies were performed using the PI3K/AKT agonist 740Y-P to validate pathway involvement. The in vivo experiments employed a ligature-induced periodontitis model in Sprague-Dawley rats (n = 4 per group), with treatment groups consisting of control, periodontitis, and PLB-treated animals. Outcome measures included micro-CT analysis of alveolar bone loss, histological evaluation of gingival inflammation, and molecular analysis of pathway markers in gingival tissues.
Cell Culture
Mouse monocyte macrophage RAW264.7 cells were obtained from the American Type Culture Collection (USA). The cells were grown in Dulbecco’s Modified Eagle Medium supplemented with 10% fetal bovine serum and placed at 37°C and 5% CO2. 100 ng/mL LPS (Beyotime, China) was introduced to RAW264.7 cells to simulate an inflammatory response in vitro. Additionally, RAW264.7 cells were exposed to a 25 µg/mL PI3K agonist 740Y-P solution (MedChemExpress, China) for 1 h, aimed at exploring the mechanism of the PI3K/AKT pathway. This concentration of 740Y-P was determined based on preliminary experiments in our laboratory, which confirmed that 25 µg/mL effectively activates the PI3K/AKT pathway in RAW264.7 cells without inducing significant cytotoxicity, and is consistent with the manufacturer’s recommended working concentration range for this cell type.
Cell Counting Kit-8 (CCK-8) Assay
RAW264.7 cells were plated in 96-well plates at 8 × 103 cells per well and then pretreated with 0, 1.25, 2.5, 5, 10, and 20 µg/mL PLB for 24 h. Subsequently, each well was introduced with 10 µL of CCK-8 solution (Beyotime). They were placed at 37°C for 3 h and then assessed at 450 nm using a microplate reader (Thermo Fisher Scientific, USA).
Western Blot
RAW264.7 cells underwent lysis with a radioimmunoprecipitation assay (RIPA) buffer (Solarbio Biotech, Beijing, China) with phenylmethylsulfonyl fluoride (NIC Biotech, Shanghai, China). Equal concentrations of proteins were then segregated using sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and shifted to polyvinylidene fluoride (PVDF) membranes (NIC Biotech, Shanghai, China). Blocking of the membranes was executed with 5% nonfat milk for 60 min at ambient temperature, followed by incubation overnight at 4°C with various primary antibodies against tumor necrosis factor-α (TNF-α) (ab6671; 1:1,000), interleukin-6 (IL-6) (ab233706; 1:1,000), interleukin-1β (IL-1β) (ab283818; 1:1,000), interleukin-10 (IL-10) (ab310329; 1:1,000), sequestosome 1 (p62) (ab109012; 1:10,000), microtubule-associated protein 1A/1B-light chain 3-II (LC3-II) (ab192890; 1:2,000), Beclin-1 (ab302669; 1:1,000), AKT (ab38449; 1:1,000), phosphorylated protein kinase b (P-AKT) (ab81283; 1:5,000), PI3K (ab302958; 1:1,000), phosphorylated phosphatidylinositol 3-kinase (P-PI3K) (ab278545; 1:1,000), and β-actin (ab6276; 1:5,000). The samples were then exposed to anti-mouse secondary antibodies for an hour at ambient temperature after washing. These antibodies were sourced from Abcam (Cambridge, UK). Visualization of the protein bands relied on an enhanced chemiluminescence system (Servicebio, Beijing, China), and the resulting data were analyzed using a ChemiDoc™ XRS system (Bio-Rad, Hercules, CA, USA).
Quantitative Real-time Polymerase Chain Reaction (qRT-PCR) Analysis
A ribonucleic acid (RNA) isolation kit from Thermo Fisher Scientific (Waltham, MA, USA) was utilized to extract total RNA; subsequently, a complementary deoxyribonucleic acid (cDNA) synthesis kit from the same manufacturer was employed to carry out reverse transcription. The PCR process involved an initial denaturation phase at 95°C for 5 min, followed by 50 cycles. Each cycle involved heating to 95°C for 20 s, cooling to 65°C for 30 s, and heating to 72°C for 30 s. The samples were analyzed on a CFX96 Touch Real-Time PCR Detection System (Bio-Rad, Hercules, CA, USA). Table 1 lists the sequences of the primers used in this study.
Quantitative Real-time Polymerase Chain Reaction (qRT-PCR) Primer Sequences.
Enzyme-linked Immunosorbent Assay (ELISA)
After treatment, the cell culture medium was gathered and subjected to centrifugation at 1,000 g for 20 min at 4°C. Cytokine levels (IL-6, IL-1β, and IL-10) were determined from the supernatant in accordance with the manufacturer’s instructions (RandD Systems, Minneapolis, MN, USA) and then assessed under a microplate reader at 450 nm, with 540 nm as the correction wavelength.
LysoTracker Red Staining
For autophagy analysis, RAW264.7 cells were plated in six-well plates at 1 × 105 cells/well. After treatment, LysoTracker Red (Thermo Fisher Scientific, Waltham, MA, USA) was introduced into the culture medium at a 1:17,000 dilution in a dark environment. After incubation for 45 min at 37°C, the samples were visualized using a fluorescence microscope (Olympus, Tokyo, Japan).
Animals Model
Female Sprague-Dawley rats (200–280 g, aged 4–6 weeks) were obtained from Zhejiang Weitonglihua Laboratory Animal Technology Co., Ltd. The rats were raised under specific pathogen-free conditions and acclimated for 7 days before the experiments. Periodontitis was induced by securing a wire around the base of the left maxillary first molar using ligation. The rats were assigned into three groups: control group (n = 4), periodontitis (negative control (NC)) group (n = 4), and PLB group (n = 4). For the PLB group, PLB was first dissolved in dimethyl sulfoxide (DMSO) to prepare a stock solution, then diluted to the final concentration with normal saline (0.9% NaCl) (the final DMSO concentration was ≤0.1%). Rats in this group received intraperitoneal injections of 4 mg/kg PLB (prepared in the aforementioned solvent) five times weekly over a 4-week period (Zheng et al., 2017). The NC group and the control group were injected with the same volume of solvent (0.1% DMSO in normal saline) to exclude solvent-related effects. DMSO at a concentration of ≤0.1% is widely recognized as nontoxic and noninflammatory in animal studies, and preliminary experiments confirmed that this solvent did not affect periodontal inflammation, alveolar bone resorption, or the PI3K/AKT pathway/autophagy-related protein expression in rats. Twice weekly, the ligation was inspected, and any ligatures that had become dislodged or loose were promptly replaced.
Micro-computed Tomography (Micro-CT) Scanning
The left maxillary alveolar bone samples from each group were scanned using a Micro-CT scanner (Bruker, USA). Mimics software was used to process the images of samples into three-dimensional (3D) reconstruction, and the distance of cemental-enamel junction–alveolar bone crest (CEJ–ABC) was determined using CTAn software (Bruker, USA).
Hematoxylin-Eosin (H&E) Staining
The rats were euthanized for subsequent histological analysis. 4 µm tissue slices were stained with H&E dyes (Beyotime, Shanghai, China) according to the manufacturer’s instructions and then observed under a light microscope (Olympus, Tokyo, Japan).
Statistical Analysis
Data were analyzed using Statistical Package for the Social Sciences (SPSS) 20.0 (SPSS Inc., Armonk, NY, USA), expressed as the mean ± standard deviation. To compare multiple datasets, one-way analysis of variance (ANOVA) was utilized, whereas for pairwise comparisons, independent sample t-tests were conducted. A p < .05 was considered statistically significant.
Results
In Vitro Cytotoxicity of PLB
Concentrations maintaining RAW264.7 cell viability above 80% were suitable for further experimentation. Our findings revealed that concentrations of PLB below 10 µg/mL exhibited no toxic effects (Figure S1 in the supplemental material).
PLB Inhibits Inflammatory Responses In Vitro
Through qRT-PCR analysis, it was observed that PLB dose-dependently inhibited the expression of TNF-α, IL-6, and IL-1β (p < .05). Simultaneously, it dose-dependently stimulated the expression of IL-10, an anti-inflammatory cytokine (p < .05, Figure 1A). ELISA assays further confirmed these findings by demonstrating corresponding changes in the secretion of these cytokines (p < .05, Figure 1B).
Plumbagin (PLB) Inhibits Inflammatory Responses In Vitro. (A) Quantitative Real-time Polymerase Chain Reaction (qRT-PCR) Analysis Revealed That PLB Dose-dependently Suppressed Tumor Necrosis Factor-α (TNF-α), Interleukin-6 (IL-6), and Interleukin-1β (IL-1β), Accompanied by Enhancing Interleukin-10 (IL-10) Expression. (B) Enzyme-linked Immunosorbent Assay (ELISA) Assays Further Demonstrated Corresponding Changes. ***p < .001 versus Control.

PLB Inhibits the PI3K/AKT Signaling Pathway In Vitro
Through Western blot analysis, upon LPS stimulation, a notable elevation in the expression of P-PI3K and P-AKT was detected (Figure 2A). Treatment with PLB, however, resulted in a pronounced and concentration-dependent decrease in the expression of these proteins, indicating suppression of the PI3K/AKT pathway by PLB (p < .05, Figure 2B).

PLB Promotes Autophagy In Vitro
In comparison to the group treated with LPS, the PLB-treated groups (5 and 10 µg/mL) displayed increased autophagic activity, as evidenced by the enhanced fluorescence intensity (p < .05, Figure 2C and 2D). The findings further supported the above observations via Western blot, demonstrating elevated levels of LC3-II and Beclin-1, which are proteins associated with autophagy. Additionally, it revealed a reduction in p62 levels in the groups treated with PLB (p < .05, Figure 2E and 2F).
Effect of PI3K/AKT Agonist 740Y-P on PLB-induced Autophagy
RAW264.7 cells were treated with the PI3K/AKT agonist 740Y-P. LysoTracker Red staining revealed that while PLB alone enhanced autophagic activity, co-treatment with 740Y-P significantly reduced autophagic fluorescence (p < .05, Figure 3A and 3B). It was noted that 740Y-P mitigated the increase in LC3-II and Beclin-1 expression and reversed the reduced p62 levels induced by PLB (p < .05, Figure 3C and 3D). These findings suggest that PLB enhances autophagy through its suppressive effects on the PI3K/AKT pathway.
Effect of Phosphatidylinositol 3-Kinase/Protein Kinase B (PI3K/AKT) Agonist 740Y-P on Plumbagin (PLB)-induced Autophagy. (A, B) LysoTracker Red Staining Revealed That While PLB Alone Enhanced Autophagic Activity, Co-treatment with 740Y-P Significantly Reduced Autophagic Fluorescence. (C, D) Through Western Blot Analysis, We Observed That 740Y-P Mitigated the Increase in Microtubule-Associated Protein 1A/1B-Light Chain 3-II (LC3-II) and Beclin-1 and Reversed the Decreased Sequestosome 1 (p62) Levels Induced by PLB. β-actin Served as the Reference Standard. **p < .05 versus PLB. ***p < .001 versus PLB.

PLB Inhibits Inflammation by Suppressing the PI3K/AKT Signaling Pathway
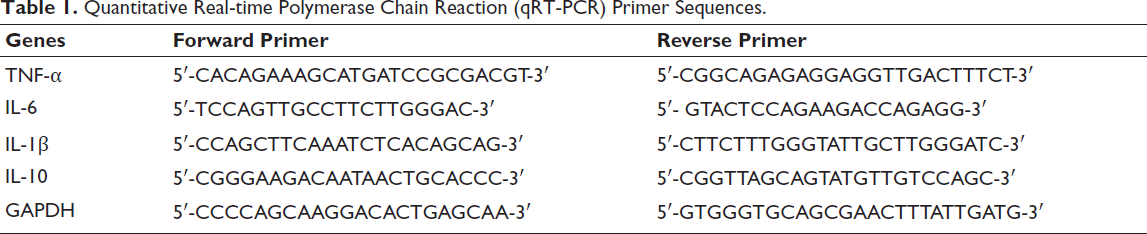
PLB suppressed the production of TNF-α, IL-6, and IL-1β, while enhancing IL-10 levels. However, 740Y-P partially counteracted these effects (p < .05, Figure 4A). ELISA assays also yielded consistent results, confirming that PLB achieves its inflammatory-reducing properties via the PI3K/AKT pathway inhibition (p < .05, Figure 4B).
Effect of Plumbagin (PLB) and 740Y-P on Inflammation Responses. (A) Quantitative Real-time Polymerase Chain Reaction (qRT-PCR) Analysis Showed That PLB Suppressed the Production of Tumor Necrosis Factor-α (TNF-α), Interleukin-6 (IL-6), and Interleukin-1β (IL-1β) and Enhanced Interleukin-10 (IL-10) Levels. However, 740Y-P Partially Counteracted These Effects. (B) Enzyme-linked Immunosorbent Assay (ELISA) Assays Yielded Consistent Results. **p < .05 versus Negative Control (NC). ***p < .001 versus NC.
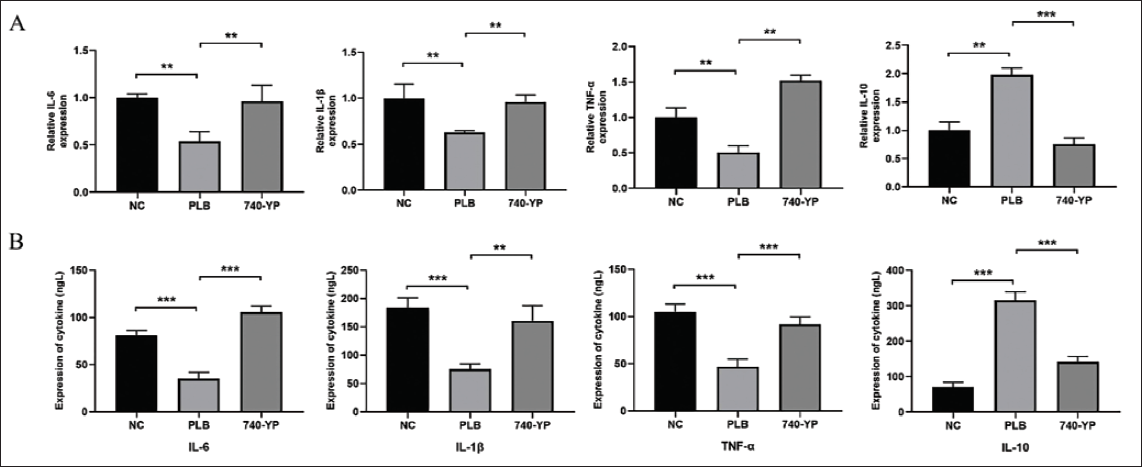
PLB Inhibits Periodontitis in Rat Models of Periodontitis
The therapeutic potential of PLB in periodontitis was evaluated using a rat model. Upon H&E staining, we found a notable decrease in inflammatory cell infiltration in the gingival tissue of the PLB-treated group when contrasted with the NC group (Figure 5A). Micro-CT scanning showed that PLB treatment mitigated alveolar bone resorption, as evidenced by reduced bone loss and improved bone height. Furthermore, in comparison to the NC group, the PLB-treated group demonstrated notably shorter CEJ–ABC distances of the second molars (p < .05, Figure 5B). Moreover, H&E staining did not reveal notable toxic effects of PLB on major organs (Figure S2 in the supplemental material).
Plumbagin (PLB) Inhibits Periodontitis in Rat Models of Periodontitis. (A) Upon Hematoxylin-Eosin (H&E) Staining, We Found a Notable Decrease in Inflammatory Cell Infiltration in the Gingival Tissue of PLB-treated Group When Contrasted with the Negative Control (NC) Group. (B) Micro-computed Tomography (Micro-CT) Scanning and Statistical Analysis of Cemental-Enamel Junction–Alveolar Bone Crest (CEJ-ABC) Distance in Each Group of Rats After Three-dimensional (3D) Reconstruction Showed That PLB Treatment Mitigated Alveolar Bone Resorption, as Evidenced by Reduced Bone Loss and Improved Bone Height. ***p < .001 versus NC.

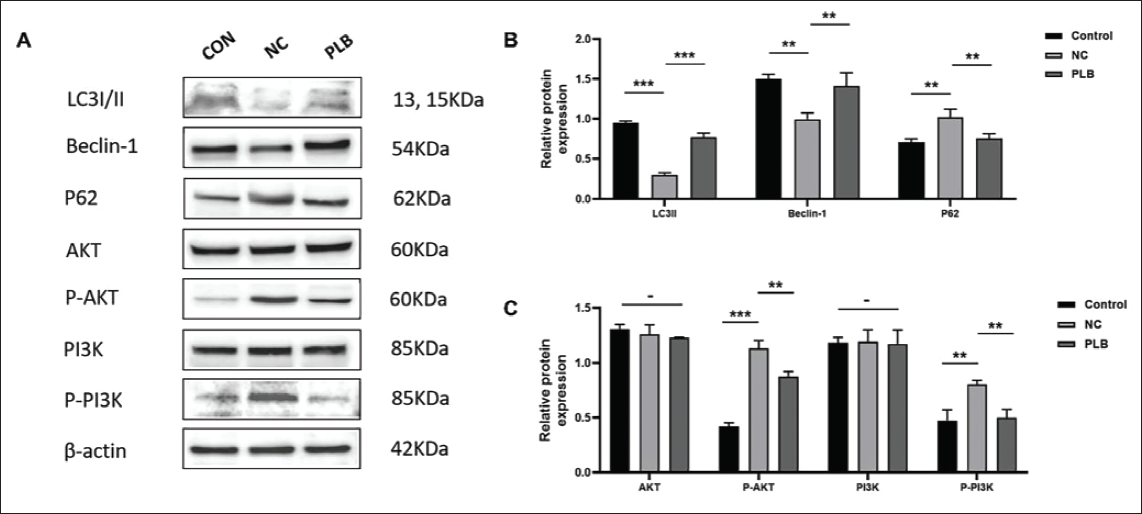
PLB Modulates PI3K/AKT Signaling Pathway and Autophagy In Vivo
Western blot analysis of rat gingival tissue confirmed that compared to the NC group, PLB led to a downregulation of P-PI3K, P-AKT, and p62 proteins and an upregulation of LC3-II and Beclin-1 proteins (p < .05, Figure 6A–6C). These results indicate that PLB modulates autophagy via the PI3K/AKT pathway inhibition.
Effect of Plumbagin (PLB) on Autophagy In Vivo. (A–C) Western Blot Analysis of Rat Gingival Tissue Confirmed That PLB Led to a Downregulation of Phosphorylated Phosphatidylinositol 3-Kinase (P-PI3K), Phosphorylated Protein Kinase B (P-AKT), and Sequestosome 1 (p62) Proteins and an Upregulation of Microtubule-Associated Protein 1A/1B-Light Chain 3-II (LC3-II) and Beclin-1 Proteins Compared to the Negative Control (NC) Group. β-actin Served as the Reference Standard. -p > .05 versus Control. **p < .05 versus NC. ***p < .001 versus NC.
Discussion
Periodontitis, a chronic inflammatory disease primarily affecting the gingiva and periodontal tissues, poses significant risks to oral health (Arora & Arora, 2021). Current pharmacological treatments often have limitations due to side effects, highlighting the need for safer and more effective therapeutic strategies (Kashyap et al., 2022; Pop et al., 2022). Recent advancements in the field of Chinese herbal medicine have shed light on the potential efficacy of PLB in treating a wide array of conditions, such as acne, fractures, and bacterial infections, and having beneficial biological effects (Inbaraj & Chignell, 2004; Padhye et al., 2012; Yan et al., 2015). While PLB’s anti-inflammatory effects have been documented in various diseases, the precise underlying mechanisms of its action in periodontitis remain elusive.
Pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α play critical roles in the pathogenesis of periodontitis by promoting inflammation and tissue destruction (Beltrami-Moreira et al., 2016; Jang et al., 2021; Kaur et al., 2020; Mai & Liao, 2020; Malaviya et al., 2017). In contrast, IL-10 acts as an anti-inflammatory cytokine, mitigating excessive immune responses (Saraiva et al., 2020). Our results showed that PLB treatment in RAW264.7 cells significantly reduced the expression of IL-1β, IL-6, and TNF-α while increasing IL-10 levels, suggesting its potent anti-inflammatory properties. These findings align with previous studies demonstrating PLB’s ability to modulate cytokine production in other inflammatory conditions (Inbaraj & Chignell, 2004; Padhye et al., 2012; Yan et al., 2015).
The PI3K/AKT signaling pathway is a key regulator of inflammatory responses and has been implicated in the progression of periodontitis (Xia et al., 2023). Our study revealed that PLB effectively downregulated the phosphorylation of PI3K and AKT, indicating its inhibitory effect on this pathway. The reversal of PLB’s anti-inflammatory effects by the PI3K agonist 740Y-P further confirmed that PLB mitigates inflammation through PI3K/AKT pathway inhibition. This mechanism is consistent with the role of PI3K/AKT in regulating macrophage activation and cytokine production during inflammatory processes.
Autophagy is a cellular self-digestion mechanism involving the isolation of cytoplasmic contents within autophagosomes and their subsequent fusion with lysosomes for degradation (Yamamoto et al., 2023). Several studies have demonstrated intricate connections involving inflammation and the process of autophagy (Liu et al., 2017; Pei et al., 2015). Our observation using LysoTracker Red staining revealed that inflammatory reactions triggered by LPS caused a decrease in autophagic activity in RAW264.7 cells. Consistent with these findings, Western blot analysis demonstrated that PLB enhanced autophagic activity through the upregulation of LC3-II and Beclin-1, which are proteins associated with autophagy, while simultaneously lowering the levels of p62. Furthermore, the introduction of the PI3K agonist 740Y-P was found to mitigate the PLB-induced enhancement in autophagic activity, as evidenced through both LysoTracker Red staining and Western blot analysis. These findings suggest that PLB achieves its anti-inflammatory effects by inhibiting the PI3K/AKT pathway and thereby modulating autophagy.
In vivo experiments, using a rat model of periodontitis induced by ligating the necks of molars, demonstrated that PLB alleviated periodontal inflammation and alveolar bone resorption without causing significant toxic side effects. Western blot analysis of tissue samples from these rats further demonstrated that PLB downregulates the production of proteins associated with the PI3K/AKT pathway while enhancing the synthesis of proteins related to autophagy.
Conclusion
This study provides compelling evidence that PLB exerts anti-inflammatory effects in periodontitis by inhibiting the PI3K/AKT pathway and enhancing autophagy. These findings not only elucidate the mechanisms underlying PLB’s therapeutic potential but also support its development as a novel treatment for periodontitis.
Limitations
Despite the promising findings, this study has several limitations. First, the research relied on an in vitro macrophage model and a rat periodontitis model, which may not fully replicate the complex immune–microbial interactions seen in human periodontitis. Additionally, the study focused primarily on the PI3K/AKT pathway and autophagy, while other inflammatory signaling pathways (e.g., nuclear factor-κB (NF-κB), mitogen-activated protein kinase (MAPK)) that contribute to periodontitis were not explored. The in vitro experiments used LPS rather than live P. gingivalis or other periodontal pathogens, potentially overlooking microbial-specific effects on autophagy and inflammation. Furthermore, the PLB dosage was limited to nontoxic concentrations, and long-term toxicity was not assessed. The small sample size in the animal study may also limit the statistical robustness of the findings. While this study included solvent control (0.1% DMSO in normal saline) in the NC and control groups to rule out solvent confounding, the solvent’s potential long-term effects (beyond the 4-week experimental period) on periodontal tissues or systemic metabolism were not evaluated—future studies could extend the observation window to address this. Most importantly, the lack of clinical validation means that the therapeutic potential of PLB in human periodontitis remains speculative. Future studies should incorporate human trials, broader mechanistic investigations, and long-term safety evaluations to confirm these findings.
Summary
This study investigates the therapeutic potential of PLB, a natural quinone derived from P. zeylanica L., in modulating periodontitis through the PI3K/AKT pathway and autophagy. Periodontitis, a chronic inflammatory disease initiated by microbial dysbiosis, leads to progressive destruction of periodontal tissues. Existing treatments remain limited by side effects, highlighting the need for novel strategies. In vitro, mouse RAW264.7 macrophages were stimulated with LPS to induce inflammatory conditions and treated with varying concentrations of PLB. PLB significantly reduced pro-inflammatory cytokines (TNF-α, IL-6, IL-1β) while enhancing anti-inflammatory IL-10 expression. Western blot and LysoTracker Red assays confirmed that PLB suppressed PI3K/AKT phosphorylation and promoted autophagy, as evidenced by increased LC3-II and Beclin-1 with decreased p62 levels. Mechanistic validation with the PI3K/AKT agonist 740Y-P revealed that PLB’s anti-inflammatory and pro-autophagic effects were mediated through inhibition of this pathway. In vivo, a ligature-induced rat model of periodontitis demonstrated that PLB alleviated gingival inflammation and reduced alveolar bone resorption. Micro-CT and histological analyses confirmed improved periodontal outcomes, while Western blot of gingival tissues showed modulation of autophagy-related proteins consistent with in vitro findings. Importantly, PLB displayed no observable systemic toxicity in major organs. Collectively, the findings highlight PLB as a promising therapeutic agent against periodontitis, acting through dual mechanisms: suppression of PI3K/AKT signaling and enhancement of autophagy. While results provide strong preclinical evidence, limitations include small sample size, reliance on animal models, and lack of clinical validation. Future studies should focus on human trials, long-term safety, and exploration of additional inflammatory pathways.
Footnotes
Abbreviations
3D: Three-dimensional; AKT: Protein kinase B; ANOVA: Analysis of variance; CCK-8: Cell Counting Kit-8; cDNA: Complementary deoxyribonucleic acid; CEJ–ABC: Cemental-enamel junction–alveolar bone crest; DMSO: Dimethyl sulfoxide; ELISA: Enzyme-linked immunosorbent assay; GAPDH: Glyceraldehyde-3-phosphate dehydrogenase; H&E: Hematoxylin-Eosin; IL-1β: Interleukin-1β; IL-10: Interleukin-10; IL-6: Interleukin-6; LC3-II: Microtubule-associated protein 1A/1B-light chain 3-II; LPS: Lipopolysaccharides; MAPK: Mitogen-activated protein kinase; Micro-CT: Micro-computed tomography; NC: Negative control; NF-κB: Nuclear factor-κB; P. gingivalis: Porphyromonas gingivalis; PI3K/AKT: Phosphatidylinositol 3-kinase/protein kinase B; PLB: Plumbagin; p62: Sequestosome 1; P-AKT: Phosphorylated protein kinase B; P-PI3K: Phosphorylated phosphatidylinositol 3-kinase; PCR: Polymerase chain reaction; PVDF: Polyvinylidene fluoride; qRT-PCR: Quantitative real-time polymerase chain reaction; RIPA: Radioimmunoprecipitation assay; RNA: Ribonucleic acid; SDS-PAGE: Sodium dodecyl sulfate-polyacrylamide gel electrophoresis; SPSS: Statistical Package for the Social Sciences; TNF-α: Tumor necrosis factor-α.
Authors’ Contributions
LW conceived and designed the experiments. SL, YZ, and CT performed most of the experiments. HW and FQ performed the data analysis. SL provided administrative support. All authors wrote and reviewed this manuscript.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval and Informed Consent
All experimental procedures involving animals were approved by the Ethical Review Committee of Jiangsu Kebiao Medical Technology Group Co., Ltd., and conducted in accordance with institutional and national guidelines for the care and use of laboratory animals. Efforts were made to minimize animal suffering, including the use of anesthesia and humane euthanasia protocols.
For the in vitro studies, the RAW264.7 cell line was obtained from the American Type Culture Collection, and no human or primary animal tissues were used, thus requiring no additional ethical approval. However, all cell culture experiments followed standard biosafety and ethical research practices.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by Application Basic Research of Changzhou Science and Technology Bureau (CJ20220060); Science and Technology Project of Changzhou Municipal Health Commission (Major) (ZD202321); and Science and Technology Project of Changzhou Municipal Health Commission (Youth) (QN202235).
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.