
Abstract
Background
When the endometrium matures in step with the required morphological changes, blastocyst implantation occurs more effectively.
Purpose
The present study aimed to examine the reproductive toxicity of apricot kernel oil (AK oil) by using an animal model of early pregnancy in adult BALB/c mice. Materials and Methods: Fifty adult BALB/c mice were first acclimated and then mated. Thirty-two confirmed pregnant mice were allocated into four groups. Monitoring involved recording weight gain and abortion rates, and on gestation day 15 the animals were euthanized for collection of tissue and blood samples. Hematological analysis was performed to evaluate multiple blood indices, while levels of sex hormones and serum inflammatory markers (CRP, IL-6, IL-1β, TNF-α) were measured. Antioxidant capacity, lipid peroxidation, and thiol content in uterine tissue were determined, and gene expression associated with the p53/Cas-3/Bax/Bcl-2 apoptotic pathway was assessed by RT-qPCR.
Results
Exposure to AK oil resulted in significant (p <0.05) decreases in maternal weight gain and placental weight, together with a significant (p <0.05) increase in uterine weight. Hematological findings demonstrated raised WBC and lymphocyte counts, along with significantly reduced (p <0.05) progesterone and estrogen concentrations. Elevated (p <0.05) cytokine levels were observed at the higher AK oil doses. Gene expression analysis showed upregulation (p <0.05) of pro-apoptotic genes and downregulation (p <0.05) of Bcl-2, collectively indicating adverse effects of AK oil on early pregnancy.
Conclusion
Accumulation of AK oil may stimulate the HPG axis and consequently promote reactive oxygen species generation, oxidative stress, mitochondrial apoptosis, and reproductive toxicity.
Introduction
The uterus is a key organ in the genital system. It has two major components: the inner endometrium, which is crucial for nidation and fetal growth, and the outer myometrium, which offers vital support and produces the contractile forces required for childbirth. Estrogen (E) and progesterone (P) are principal hormones responsible for regulating endometrial growth (Poole et al., 2023). Variations in hormonal levels can induce multiple endometrial responses, such as increased thickness, enhanced glandular secretion, stromal cell decidualization, and augmented blood perfusion, all of which are essential for effective embryo implantation (Nascimento et al., 2023). Progesterone can diminish the excitability of uterine muscle fibers, reduce the uterus’s sensitivity to oxytocin, and consequently lower uterine contractions. In addition, it may attenuate the maternal immune response, thereby supporting the development of embryos within the uterine environment. When the endometrium matures in step with the required morphological changes, blastocyst implantation occurs more effectively. In contrast, if endometrial maturation falls behind these morphological alterations, endometrial receptivity to the blastocyst decreases, which in turn lowers the likelihood of pregnancy (Dias Da Silva et al., 2024). The estrogen receptor (ER) and progesterone receptor (PR) are protein receptors located in the glandular epithelial and stromal cells of the endometrium. These receptors bind specifically to their respective hormones, forming hormone–receptor complexes that enable the hormones to exert their biological actions and to drive endometrial proliferation. Aberrant expression levels of ER and PR in the endometrium can disturb the regulation and secretion of estrogen (E) and progesterone (P), thereby contributing to decreased pregnancy rates (Bosolasco et al., 2021).
Isoflavonoids are a class of phytoestrogens primarily found in soy and other legumes, garnering significant attention for their potential impacts on human health, particularly regarding hormonal functions and reproductive health. Studies indicate that high intake of isoflavones during pregnancy may be associated with adverse outcomes, such as low birth weight or increased risk of abortion, as maternal soy consumption can influence fetal hormone levels and alter developmental pathways (Akbaribazm et al., 2021). In rodent models, excessive consumption of isoflavonoids during pregnancy has been linked to developmental anomalies and increased rates of miscarriage. Isoflavonoids can also modulate the secretion of follicle-stimulating hormone (FSH) and luteinizing hormone (LH), potentially altering ovarian function and ovulation (Caceres et al., 2023). Research shows that isoflavones exhibit both estrogenic and anti-estrogenic effects, affecting levels of estrogen and progesterone during the menstrual cycle and pregnancy. Additionally, isoflavonoids can influence relaxin and oxytocin levels, potentially impacting labor and delivery (Sleiman et al., 2021). Isoflavonoids have been found to influence pathways like PI3K/AKT, crucial for cell survival and apoptosis, with studies indicating that certain isoflavones can upregulate pro-apoptotic proteins (like Bax) and downregulate anti-apoptotic proteins (like Bcl-2), leading to increased apoptosis in reproductive tissues (Zughaibi et al., 2021). They also possess antioxidant properties, reducing oxidative stress in tissues; however, at high concentrations, they may contribute to oxidative damage. Isoflavonoids can modulate the release of pro-inflammatory cytokines, such as interleukin (IL)-6, IL-1β, and tumor necrosis factor-alpha (TNF-α), which are associated with infertility and adverse pregnancy outcomes. Some studies suggest that isoflavones may reduce inflammation in reproductive tissues, promoting better outcomes (Chaturvedi et al., 2021). Isoflavonoids have been shown to affect contraction patterns and smooth muscle responsiveness in the myometrium, impacting labor, and can alter the endometrial environment, crucial for implantation and supporting a growing embryo. The modulation of ovarian hormones by isoflavonoids may lead to changes in follicular development and ovulation, potentially affecting fertility. While isoflavonoids possess potential health benefits, their effects on pregnancy and reproductive health can vary widely based on dosage, timing, and individual health conditions (Blázovics et al., 2022). Therefore, it is essential for pregnant women or those trying to conceive to consult with a healthcare provider about their dietary choices. For further insights, consider reviewing clinical trials investigating the effects of soy isoflavonoids during pregnancy, animal studies focusing on rodent models assessing isoflavonoid effects on reproductive health, and meta-analyses summarizing data from multiple studies on the impacts of isoflavonoids.
Apricot kernel oil (AK oil), obtained from the seeds of apricots (Prunus armeniaca L.), contains a variety of bioactive constituents, including fatty acids, vitamins, and amygdalin, the latter of which can release cyanide following metabolism (Tahir et al., 2024). Its toxicity is largely attributed to this cyanogenic component, which can cause symptoms such as nausea and dizziness and, in severe cases, may lead to respiratory failure and death, raising regulatory concerns regarding its ingestion. Animal investigations indicate that high doses may induce reproductive toxicity and elevate abortion rates through mechanisms such as hormonal imbalance and oxidative stress, although limited data are available concerning its effects in humans (Hong et al., 2024; Tahir et al., 2024). Despite its traditional use for various health-related purposes, the safety of AK oil during pregnancy has not been clearly established, and the presence of cyanogenic compounds warrants caution and medical consultation for pregnant individuals. Considering the fundamental importance of normal uterine structure and function in maintaining a healthy pregnancy, it is necessary to determine how AK oil exposure may influence the uterus. This includes identifying any potential toxic effects and elucidating possible mechanisms of reproductive toxicity, if present. In this study, we examined the adverse effects of AK oil on early pregnancy by analyzing hematological parameters, serum concentrations of sex hormones, AK oil-related outcomes, and morphological alterations in the uterus. Additionally, we analyzed the expression levels of genes associated with oxidative stress, apoptosis, and sex hormone receptors.
Materials and Methods
Preparation of AK Oil
A total of 2,000 g of dried apricot kernels were first ground into a fine powder using a soil grinder (Catalog No. H-4199.5F; Humboldt Mfg. Co., USA). This powder was then mixed with a hexane–methanol solution in a 30:70 v/v proportion and incubated at 40°C for 72 h in the absence of light. After incubation, the mixture was passed through a No. 42 paper filter (Catalog No. 1442-125; Bedfont Lakes Business Park, Feltham, UK). The solvent phase was then removed using a rotary evaporator (Model No. 9230; Buchi Rotavapor, Buchi Instruments, Basel, Switzerland). The recovered oil was then weighed, showing an extraction yield of 20%, and stored at –4°C (Hong et al., 2024; Wu et al., 2024).
Animal Care and Experimental Groups
Fifty adult BALB/c mice, between 5 and 8 weeks of age and weighing approximately 32 ± 5 g, were housed in propylene cages under controlled environmental conditions. The room temperature was set at 25 ± 4°C, with relative humidity maintained at 35 ± 5%, and a 12-h light/dark cycle was used. The mice were allowed a 72-h acclimatization period in the laboratory environment before the study commenced. The animals were provided free access to tap water and regular laboratory rat pellets, and all experimental procedures adhered to international guidelines, protocols, and ethical standards approved by the Ethics Committee of the Second Affiliated Hospital of Xi’an Medical College.
After an adaptation period of 1 week, the female mice were paired with healthy male mice overnight. Detection of a vaginal plug was recorded as day 0.5 of gestation (GD). Once pregnancy was confirmed, 32 pregnant mice were randomly assigned to four groups (n = 8). One group served as the control and received 100 µL of normal saline by gavage for 14 days, whereas the remaining three groups were given AK oil by gavage at doses of 200, 400, and 800 mg/kg for the same duration. Maternal body weight and abortion incidence were monitored daily, and any abnormal behaviors were noted and documented. On GD15, the mice were euthanized, and their blood, organs, and tissues were collected for subsequent analyses (Chen et al., 2021; Hong et al., 2024; Wu et al., 2024).
Evaluation of Pregnant Outcomes
The daily conditions and behaviors of each pregnant mouse were carefully monitored. When a mouse showed a marked, rapid loss of body weight, its uterus was treated with 2% ammonium sulfide to visualize possible implantation sites, which were regarded as markers of abortion in this study. On GD15, the mice were euthanized as previously described, and the gestational sac was removed, thoroughly inspected, and photographed. The components of the gestational sac—the uterus, fetuses, and placentas—were then carefully dissected and separated. The total number of fetuses was determined, and the weights of both the fetuses and placentas were measured and documented (Ladyman et al., 2021).
Hematological Analysis and Sex Hormone Level Analysis
On GD15, euthanasia was performed using a pre-anesthetic regimen consisting of 100 mg/kg xylazine 2% (Catalog No. 7361-61-7, Sigma, St. Louis, MO, USA), followed by anesthesia with 15 mg/kg ketamine 10% (Catalog No. K1884, Sigma, St. Louis, MO, USA). After euthanasia, blood samples were obtained from the heart. Serum was obtained by centrifuging the collected blood at 10,000 g for 15 min. The hematological evaluation encompassed measurements of white blood cells (WBC), lymphocytes (LY), monocytes (MONE), neutrophils (NEUT), red blood cells (RBC), hemoglobin (Hb), and platelets (PLT). Serum concentrations of the sex hormones estradiol (E) and progesterone (P) were quantified using the ADVIA 2120i Hematology System (model number 2120i, Siemens Healthineers, Hamburg, Germany). All experimental steps followed the manufacturer’s instructions and protocols (Adiguzel & Celik-Ozenci, 2021; Chen et al., 2021).
Assessment of Serum C-reactive Protein (CRP), IL-6, IL-1β, and TNF-α
Serum samples were obtained as previously described. The concentrations of these serum biomarkers were measured using commercial enzyme-linked immunosorbent assay (ELISA) kits from R&D Systems, Inc. (Centennial, CO, USA) for CRP (Catalog No. DY999B), IL-6 (Catalog No. R6000B), IL-1β (Catalog No. RLB00), and TNF-α (Catalog No. RTA00). All measurements were performed strictly in accordance with the manufacturer’s instructions and protocols (Rauf et al., 2023).
Total Antioxidant Capacity (TAC), Lipid Peroxidation (TBARS), and Thiol Levels in Uterine Tissue
TAC in uterine tissue was assessed using ferric reducing ability of plasma (FRAP) assays. Uterine tissue was carefully dissected free from adjacent adipose tissue, and tissue homogenates were then prepared. Specifically, 100 µL of the tissue homogenate was mixed with 200 µL of cold phosphate-buffered saline and then placed into a 2 mL polyethylene tube. Subsequently, 10 µL of FRAP solution (Catalog No. EIAFECL2; Thermo Fisher Scientific Inc., Lonsee, Germany) was added, and the mixture was incubated at 25°C for 15 min. Following incubation, the sample was centrifuged at 12,000 g for 10 min, and absorbance was determined with a UV–visible spectrophotometer, in accordance with the previously described procedure (Cecchini Gualandi et al., 2023).
Thiobarbituric acid reactive substances (TBARS) assays were performed to assess uterine lipid peroxidation. In this procedure, 100 µL of uterine homogenate was transferred into a 2 mL polyethylene tube and combined with 100 µL of TBARS solution (Catalog No. EEA021; Invitrogen Inc., Lonsee, Germany). As previously described, the mixture was then incubated at 37°C for 30 min, after which absorbance was determined using a UV–visible spectrophotometer (Cecchini Gualandi et al., 2023).
For quantification of total thiol content in uterine tissue, 250 µL of the tissue homogenate was incubated with 12 µL of Tris-ethylenediaminetetraacetic acid (EDTA) for 10 min at 25°C. The absorbance of this mixture was first measured at 412 nm using an ELISA reader (Awareness Technology, Stat Fax ELISA reader, model 303 microwell readers, USA) and denoted as A1. Next, 20 µL of DTNB (Ellman’s Reagent, 5,5-dithio-bis-(2-nitrobenzoic acid)) was added to the sample, which was then incubated for a further 15 min at 25°C. The absorbance of the mixture was again read at 412 nm using the same ELISA reader and recorded as A2. A DTNB solution alone was used as the blank, and its absorbance at 412 nm was designated as B. The thiol concentration was calculated using the following formula:
Total thiol concentration (µM) = (A2 – A1 – B) × 1.07/(0.05 × 13.6) (Cecchini Gualandi et al., 2023).
Real-time Quantitative Polymerase Chain Reaction (RT-qPCR) Analysis
Total RNA was isolated from uterine tissue using the AxyPrep Multisource Total RNA Miniprep Kit (Axygen Scientific, Union City, CA, USA) in accordance with the manufacturer’s instructions. Complementary DNA (cDNA) was then generated with the Takara PrimeScript RT reagent kit (Catalog No. RTK0104; Takara Bio, Tokyo, Japan). The qPCR primers were designed by Qingke Biotechnology (Hunan, China), and RT-qPCR was carried out using TB Green Premix Ex Taq II (Catalog No. RR820A) on an Applied Biosystems thermocycler (QuantStudio 1, Chicago, IL, USA). Sequences of the qPCR primers are presented in Table 1, with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) chosen as the reference gene. Relative gene expression was computed from critical threshold (Ct) values using the 2–∆∆Ct method.
∆∆CT = [(CT Sample – Ct GAPDH) – (CT Sample – CT Control)], Fold change of genes = 2–∆∆CT (Shi et al., 2022).
Primer Sequences.

Stereological Analysis
The basic volumes of uterine tissues were estimated using immersion methods. After determining tissue shrinkage, isotropic uniform random (IUR) sections were generated with the orientator technique. Processed tissues were embedded in paraffin, and 5-µm-thick sections were prepared from the blocks (Leica Microsystems). These sections were stained with hematoxylin and eosin (H&E). Tissue shrinkage was evaluated by determining the final diameter of a circular section after processing and applying the following formula:
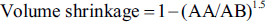
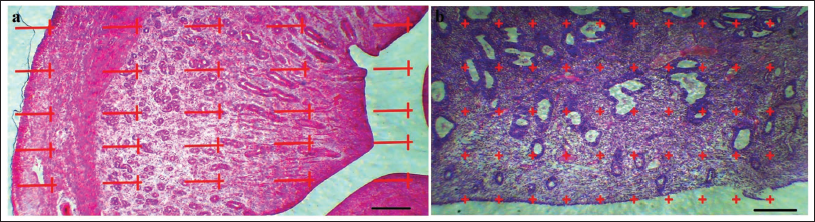
For the 5-µm sections, the volume density of the target structures—including uterine volume (UV), lumen volume (LV), mucosa volume (MV), lamina muscularis volume (LMV), and adventitia volume (ADV)—was determined using a point-counting method in combination with Delesse’s formula (Figure 1).
(a) Line Probe (25 Lines) to Estimate the Length Density of the Endometrial Structures. The Number of Lines and Points Located on Each Structure (∑p), the Length of Each of the Lines in the Probe is Given by Linear Magnification (l/p), and the Number of Lines Located on the Internal Part of Structures (Σl). Finally, Count the Numbers in the Following to Calculate the Density of the Level: L = 2× Σl/Σp × l/p (Light Photomicrograph of the Uterus; Hematoxylin and Eosin [H&E] Staining, ×100). (b) Point Probe (50 Points) to Estimate Volume Density. The Primary Volumes of Uterine Tissues were Evaluated Using Immersion Techniques, with Tissue Shrinkage Rates Estimated. The Volume Density of Various Uterine Structures, Including Uterine Volume (UV), Lumen Volume (LV), Mucosa Volume (MV), Lamina Muscularis Volume (LMV), and Adventitia Volume (ADV), was Assessed Using the Point-counting Method Alongside Delesse’s Formula:
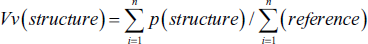
Here, 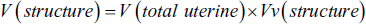
The coefficient of error (CE) for the volume estimation obtained through the Cavalieri method was determined using the following formula:
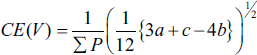
where,
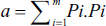
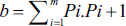
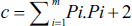
Subsequently, the coefficient of error for fractional volume was determined as follows:
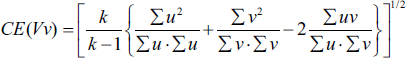
where k is the number of sections, ∑u is the total number of points on the reference space, and ∑v is the total number of points on the desired structure (Bojarzadeh et al., 2024).
Histopathological Examination
On GD15, uterine tissues from the pregnant sacs were immediately fixed in 10% formalin. The samples were then dehydrated through a graded series of ethanol and embedded in paraffin. From these paraffin blocks, 5 µm-thick sections were prepared and stained with H&E. Histological images were obtained using a BX61TRF light microscope (Olympus, Tokyo, Japan) (Russo et al., 2022).
Statistical Analyses
Statistical analyses were performed using SPSS 22.0 software (IBM Inc., Endicott, NY, USA). Data normality was evaluated with the Kolmogorov–Smirnov test, with p > .05 indicating a normal and homogeneous distribution. **Comparisons across the different groups were performed using one-way analysis of variance (ANOVA). All data are reported as mean ± standard deviation (SD). Statistical significance was defined as p < .05 (marked with *) and p < .01 (marked with **).
Results
Body Weight Changes in Pregnant Mice
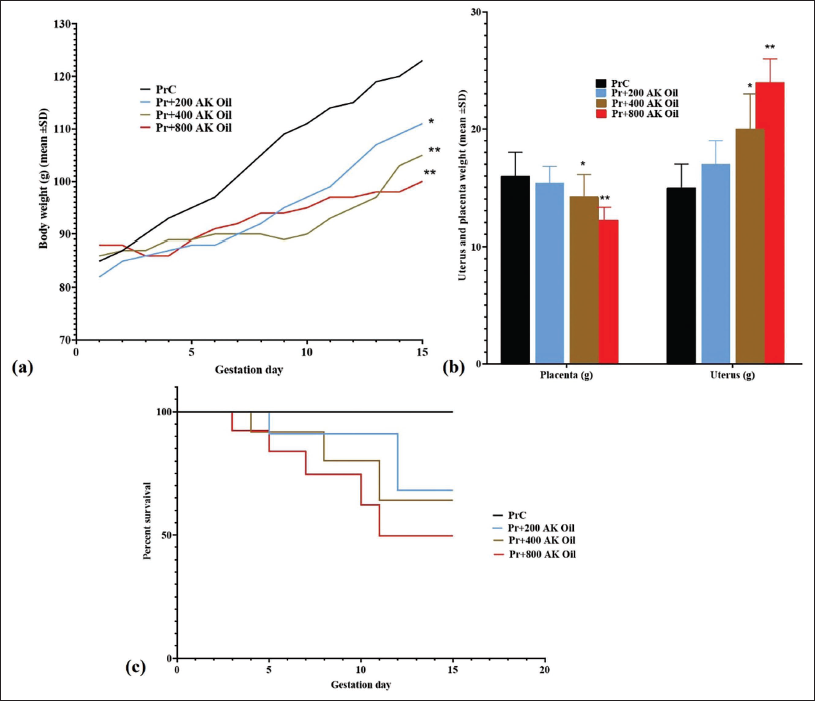
Under normal conditions, an increase in body weight was anticipated as gestation progressed. In contrast, our findings showed a marked attenuation of weight gain on GD15 in mice treated with 200 mg/kg (p < .05), 400 mg/kg (p < .01), and 800 mg/kg (p < .01) of AK oil, relative to the control group (PrC) (Figure 2a).
(a) The Body Weight Growth (g), (b) Placenta and Uterus Weight (g), and (c) Abortion Rate (%) in Pregnant Mice After Oral Exposure to Apricot Kernel Oil (AK Oil). *p < .05 and **p < .01 Versus the Control Group (PrC), n = 6, Values were Shown as Mean ± Standard Deviation (SD).
The Weight of the Fetus and Placenta
The mean placental weight showed a significant decrease in the groups treated with 400 mg/kg (p < .05) and 800 mg/kg (p < .05) of AK oil. In contrast, uterine weight exhibited a significant increase in these same groups (400 mg/kg, p < .05; 800 mg/kg, p < .01) on GD15 relative to the control group (PrC) (Figure 2b).
Abortion Rate in Pregnant Mice
In this investigation of AK oil toxicity, the experimental groups displayed varying fetal abortion rates when compared with the control group, which exhibited no abortion events. Specifically, abortion rates of 30%, 40%, and 50% were observed in the groups receiving 200 mg/kg, 400 mg/kg, and 800 mg/kg of AK oil, respectively (Figure 2c).
Hematology and Serum Biochemistry
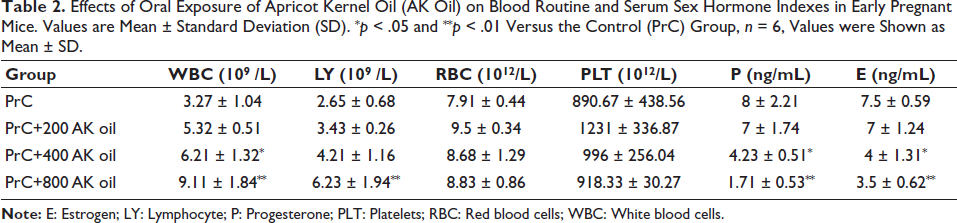
In the PrC group, WBC, LY, RBC, and PLT values were 3.27 ± 1.04, 2.65 ± 0.68, 7.91 ± 0.44, and 890.67 ± 438.56, respectively. After exposure to 200 mg/kg AK oil, WBC, LY, and RBC counts increased, and PLT levels rose markedly. At a dose of 400 mg/kg, WBC and LY continued to increase, whereas progesterone and estrogen levels decreased significantly (p < .05). In the 800 mg/kg group, WBC and LY showed further increases, accompanied by more pronounced declines in progesterone and estrogen (p < .01). Overall, AK oil exposure significantly affects hematological indices and sex hormone levels in early pregnant mice, with more intense changes at higher doses (Table 2).
Effects of Oral Exposure of Apricot Kernel Oil (AK Oil) on Blood Routine and Serum Sex Hormone Indexes in Early Pregnant Mice. Values are Mean ± Standard Deviation (SD). *p < .05 and **p < .01 Versus the Control (PrC) Group, n = 6, Values were Shown as Mean ± SD.
Serum Cytokines
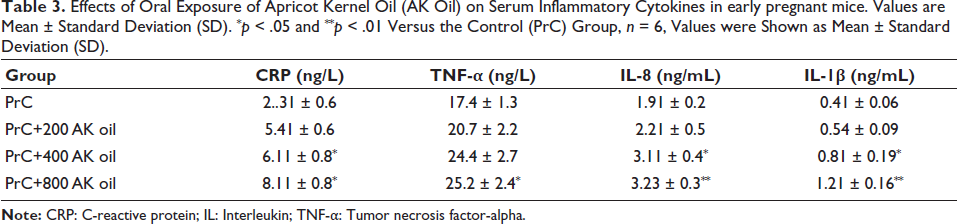
The findings demonstrate a significant rise in the levels of CRP, TNF-α, IL-8, and IL-1β in mice treated with higher doses of AK oil [400 (p < .05) and 800 (p < .01) mg/kg] compared with the control group, indicating a dose-dependent inflammatory response associated with AK oil exposure during early pregnancy (Table 3).
Effects of Oral Exposure of Apricot Kernel Oil (AK Oil) on Serum Inflammatory Cytokines in early pregnant mice. Values are Mean ± Standard Deviation (SD). *p < .05 and **p < .01 Versus the Control (PrC) Group, n = 6, Values were Shown as Mean ± Standard Deviation (SD).
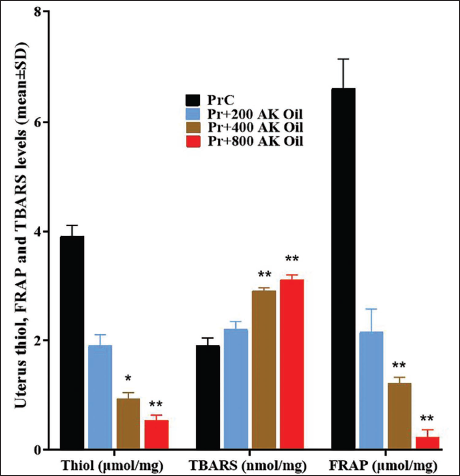
Uterine Tissue FRAP, TBARS, and Thiol
The control group (PrC) had significantly higher thiol concentrations (3.91 µmol/mg) than the AK oil-treated groups. The 200 mg/kg group had a mean thiol level of 1.91 µmol/mg (p < .05), the 400 mg/kg group had 0.94 µmol/mg (p < .01), and the 800 mg/kg group had 0.54 µmol/mg (p < .01). TBARS levels increased substantially in the AK oil-treated mice: the PrC group exhibited a TBARS value of 1.91 nmol/mg, the 200 mg/kg group reached 2.21 nmol/mg (p < .05), and the 400 mg/kg and 800 mg/kg groups showed even higher values of 2.91 nmol/mg (p < .01) and 3.12 nmol/mg (p < .01), respectively. FRAP results demonstrated a marked decline in antioxidant capacity in AK oil-treated groups, with the PrC group showing a FRAP value of 6.61 µmol/mg, compared with 2.16 µmol/mg (p < .05) in the 200 mg/kg group, 1.22 µmol/mg (p < .01) in the 400 mg/kg group, and a significantly lower value of 0.23 µmol/mg (p < .01) in the 800 mg/kg group. Collectively, these data indicate that oral AK oil exposure markedly alters antioxidant defenses and lipid peroxidation in uterine tissues during early pregnancy (Figure 3).
The Mean Uterus Tissue Levels of Ferric Reducing Ability of Plasma (FRAP) and Thiobarbituric Acid Reactive Substances (TBARS) in Pregnant Mice After Oral Exposure to Apricot Kernel Oil (AK Oil). *p < .05 and **p < .01 Versus the Control Group (PrC), n = 6, Values were Shown as Mean ± Standard Deviation (SD).
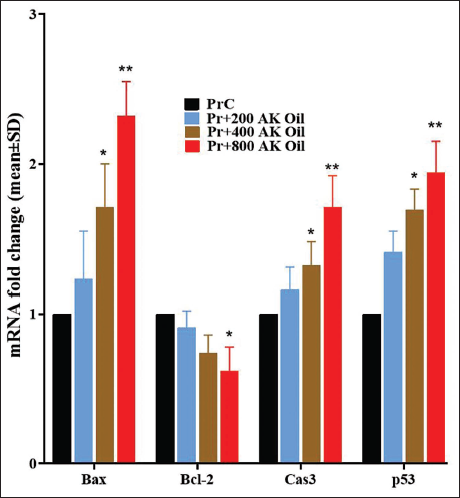
Expression of Uterine Bax, Bcl-2, p53, and Caspase-3 Genes
The present study assessed the influence of oral AK oil exposure on uterine expression of the genes Bax, Bcl-2, p53, and caspase-3 in early pregnant mice. Bax expression rose significantly from a control value of 1.00–1.23 (200 mg/kg, p < .05), 1.71 (400 mg/kg, p < .01), and 2.32 (800 mg/kg, p < .01). In contrast, Bcl-2 expression decreased from 1.00 in the control group to 0.91 (200 mg/kg), 0.74 (400 mg/kg), and 0.62 (800 mg/kg), with all values significantly lower than the control (p < .05). Caspase-3 levels increased from 1.00 in controls to 1.16 (200 mg/kg), 1.32 (400 mg/kg), and 1.71 (800 mg/kg), with significant elevations in the latter two groups (p < .05 and p < .01, respectively). Finally, p53 expression increased from 1.00 in controls to 1.41 (200 mg/kg), 1.69 (400 mg/kg), and 1.94 (800 mg/kg), with all treated groups showing significant increases (p < .05 and p < .01). Overall, these results suggest that AK oil exposure markedly modulates the expression of pro-apoptotic and anti-apoptotic genes in uterine tissue during early pregnancy (Figure 4).
The Mean Uterus Ratio of Bax, Bcl-2, Caspase-3, and p53 Gene Expression in Pregnant Mice After Oral Exposure to Apricot Kernel Oil (AK Oil). *p < .05 and **p < .01 Versus the Control Group (PrC), n = 6, Values were Shown as Mean ± Standard Deviation (SD).
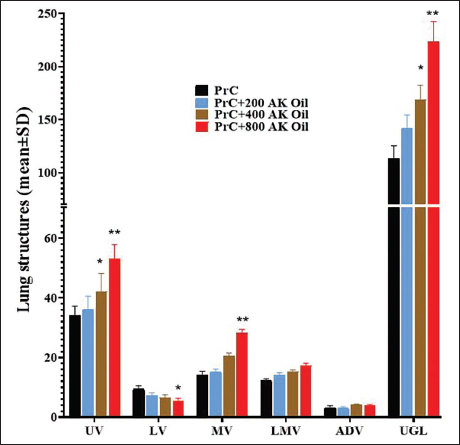
Uterine Stereological Parameters
The study investigated the effects of AK oil on uterine stereological indexes in early pregnant mice. UV significantly increased with higher doses of AK oil, with the PrC group showing a UV of 34 ± 3.2, rising to 53.21 ± 4.6 in the PrC+800 group (p < .05). Conversely, LV decreased from 9.4 ± 1.2 in the control to 5.42 ± 0.91 in the highest dose group (p < .01). MV and LMV also increased significantly, with MV rising from 14.2 ± 1.2 to 28.23 ± 1.2 and LMV from 12.21 ± 0.67 to 17.2 ± 0.91. ADV remained stable across groups, while uterine glands length (UGL) increased markedly, from 113.21 ± 12.23 in the control to 223.3 ± 19.2 in the highest dose group (p < .01) (Figure 5).
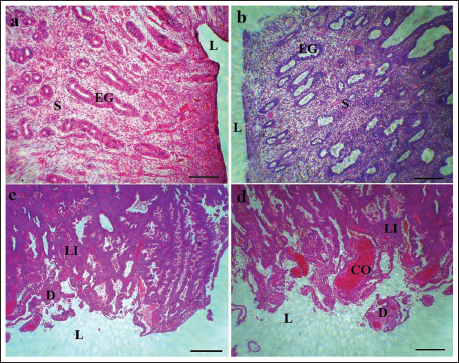
Uterine Histopathology Changes
In the AK oil-treated groups, changes in the uterine endometrium were observed in a dose-dependent manner. These changes ranged from mild at a dose of 200 mg/kg to severe lesions at 800 mg/kg. Notable findings included increased lymphocytic infiltration, degeneration of the endometrial gland stromal tissue, occlusion of uterine spiral vessels, generalized mucosal hemorrhage, and endometrial gland edema, all of which also exhibited dose-dependent effects (Figure 6).
The Histopathological Changes in Uterine Tissue in (a) PrC, (b) PrC+200 Apricot Kernel Oil (AK Oil), (c) PrC+400 AK Oil, and (d) PrC+800 AK Oil. Lymphatic Infiltration (LI), Uterus Lumen (L), Congestion (CO), Degeneration/Necrosis (D), Uterine Stroma (S), and Endometrial Gland (EG) (Hematoxylin and Eosin [H&E] Staining × 100, Scale Bar = 200 µm).
Discussion
This study examines AK oil’s effects on pregnant mice, showing reduced weight gain, decreased placental weights, and increased abortion rates at higher doses. Hematological analyses revealed elevated WBC and decreased hormone levels, alongside increased inflammatory markers and altered gene expression, indicating significant impacts on uterine health and fetal development.
Endometrial toxicity refers to harmful effects on the endometrial lining of the uterus, impacting fertility and pregnancy, often through mechanisms like oxidative stress, inflammation, and disruption of hormonal signaling. Abortion can be induced by various factors, including hormonal imbalances and toxic substances, with research indicating that certain plant extracts may lead to abortifacient effects in animal models (How et al., 2021). Key hormones involved include estrogen, which regulates reproductive functions and whose disturbances can cause menstrual irregularities; progesterone, essential for maintaining pregnancy, where imbalances may lead to miscarriage; and oxytocin, involved in labor and lactation, with dysregulation affecting reproductive health. While there are limited direct studies on plant extracts and endometrial toxicity in humans, most evidence being anecdotal or derived from traditional medicine practices, animal studies have provided some insights (Singh et al., 2024). Isoflavonoids, a class of phytoestrogens primarily found in soybeans and other legumes, are known for their estrogen-like activity and potential health benefits or toxicity. They can affect the endometrial lining through mechanisms such as oxidative stress and hormonal disturbance, with research indicating that certain isoflavonoids may induce changes in endometrial tissue, leading to toxicity (Tang & Zhang, 2022). In the present study, AK oil disrupted the HPA axis by mimicking estrogen and suppressed the synthesis of estrogen and progesterone in a dose-dependent manner. Additionally, it not only disrupted hormonal pathways but also induced inflammatory pathways, enhancing pro-inflammatory cytokines in early pregnant mice through the secretion of CRP, TNF-α, IL-8, and IL-1β. This response led to LY infiltration in the endometrium of the AK oil-exposed groups.
Isoflavonoids such as formononetin, daidzein, and biochanin A are phytoestrogens that can mimic estrogen in the body, potentially influencing the HPG axis and affecting the secretion of hormones like LH and FSH. Studies suggest these compounds can modulate estrogen levels, impacting ovarian function and follicular development (Singh et al., 2024). Flavonoids, including apigenin, quercetin, and kaempferol, are known for their antioxidant properties and may influence reproductive toxicity by affecting menstrual cycle phases (follicular and secretory) through increasing estrogen and progesterone levels. Research indicates their potential roles in endometrial gland formation and overall reproductive toxicity. Isoflavonoids and flavonoids can enhance or inhibit estrogenic activity, influencing estradiol levels and reproductive processes, while also potentially affecting progesterone secretion and impacting the secretory phase of the menstrual cycle and endometrial toxicity (Sleiman et al., 2021). Some studies suggest that high doses of isoflavonoids may have abortifacient properties, impacting pregnancy outcomes in animal models; for instance, studies in mice and rats have shown varying effects on reproductive health, with some isoflavonoids linked to altered hormonal levels and potential abortion, while limited studies in rabbits and sheep suggest impacts on reproductive performance and hormonal balance (Bernstein et al., 2021). Isoflavonoids can mimic estrogen, potentially leading to hormonal imbalances, and may alter progesterone levels, affecting pregnancy maintenance, as well as influencing oxytocin secretion, which impacts labor and lactation (Hong et al., 2022). In terms of apoptosis pathways, isoflavonoids may influence the expression of pro-apoptotic (Bax) and anti-apoptotic (Bcl-2) proteins, affecting cell survival in the endometrium. They can activate the p53 pathway, promoting apoptosis in endometrial cells. They may also suppress the Akt/PI3K signaling pathway, crucial for cell survival and growth, influence PTEN activity, and enhance the activity of caspase-3, a key step in the apoptotic pathway (Sleiman et al., 2021; Zughaibi et al., 2021). Overall, research indicates that isoflavonoids significantly impact endometrial toxicity, abortion, hormonal disturbances, inflammatory cytokine secretion, and apoptosis pathways in both human and animal models.
Assessing blood cell parameters, RBC, LY, and PLT, provides valuable insights into the maternal physiological state and immune response in abortion models. Changes in WBC counts reflect immune reactions to stressors, while RBC evaluations indicate the blood’s capacity for oxygen and nutrient transport; anemia can pose significant risks for fetal distress (Dias Da Silva et al., 2024). LY counts reveal the status of the adaptive immune system, and PLT counts assess hemostatic balance, where abnormalities may lead to complications. These hematological parameters also serve as biomarkers for physiological stress and toxicity, which can influence abortion rates. Overall, analyzing these blood parameters enhances our understanding of the mechanisms underlying abortion and their correlation with adverse fetal outcomes (Bosolasco et al., 2021). In the PrC group, the initial counts of WBC, LY, RBC, and PLT were recorded at 3.27 ± 1.04, 2.65 ± 0.68, 7.91 ± 0.44, and 890.67 ± 438.56, respectively. Following exposure to 200 mg/kg of AK oil, there was a noticeable increase in WBC, LY, and RBC levels, along with a significant rise in PLT counts. At the 400 mg/kg dosage, both WBC and LY counts continued to increase, while the 800 mg/kg group demonstrated a further increase in WBC and LY counts. These results indicate that AK oil exposure significantly alters blood parameters in early pregnant mice, with effects becoming more pronounced at higher doses. While some studies offer promising insights into their protective effects, further research is needed to fully elucidate the mechanisms and implications of isoflavonoid exposure on reproductive health. In the present study, AK oil increased the apoptosis rate of endometrial stromal cells in early pregnant mice by enhancing cytochrome c-dependent mitochondrial apoptotic pathways, as evidenced by the expression of Bax, Bcl-2, and caspase-3 genes. The results also significantly indicated that AK oil enhances oxidative stress in the uterine tissue of early pregnant mice, leading to an unexpected decrease in total antioxidant capacity and an increase in lipid peroxidation (Tareen et al., 2021). This finding contrasts with the typical effects of polyphenolic compounds and may be linked to the induction of abortion and uterine toxicity associated with isoflavone compounds. Furthermore, various studies have reported the presence of cyanide compounds in AK oil, suggesting that the increased abortion rates observed in animal models treated with AK oil could also be attributed to these compounds.
Conclusion
The findings of this study indicate that AK oil exposure significantly affects the reproductive health of pregnant mice, as evidenced by reduced weight gain, decreased placental weights, and increased abortion rates at higher doses. Hematological and biochemical analyses revealed notable alterations in blood parameters and hormone levels, alongside elevated inflammatory markers and changes in gene expression related to apoptosis. The observed endometrial toxicity, characterized by oxidative stress and inflammatory responses, suggests that AK oil disrupts hormonal signaling and may adversely affect fetal development. These results underscore the importance of exercising caution regarding the use of AK oil during pregnancy due to its potential abortifacient properties and toxic effects. Clinically, this raises concerns about the safety of AK oil in pregnant populations, necessitating further investigation to elucidate the underlying mechanisms and their implications for reproductive health. However, limitations of this study include the reliance on animal models, which may not fully replicate human responses, and the need for additional research to confirm these findings in clinical settings.
Footnotes
Abbreviations
ADV: Adventitia volume; AK oil: Apricot kernel oil; BALB/c: Bayliss, a laboratory strain of mice; CE: Coefficient of error; CRP: C-reactive protein; E: Estradiol; ELISA: Enzyme-linked immunosorbent assay; FSH: Follicle-stimulating hormone; FRAP: Ferric reducing ability of plasma; GD: Gestation day; Hb: Hemoglobin; IL: Interleukin; IUR: Isotropic uniform random; LH: Luteinizing hormone; LMV: Lamina muscularis volume; LV: Lumen volume; LY: Lymphocytes; MONE: Monocytes; MV: Mucosa volume; NEUT: Neutrophils; P: Progesterone; PLT: Platelets; RBC: Red blood cells; RT-qPCR: Real-time quantitative polymerase chain reaction; TAC: Total antioxidant capacity; TBARS: Thiobarbituric acid reactive substances; TNF-α: Tumor necrosis factor-alpha; UGL: Uterine glands length; UV: Uterine volume; WBC: White blood cells.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
The experimental protocols of this study were approved by the Second Affiliated Hospital of Xi’an Medical College ethics committee.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Informed Consent
Not applicable.