
Abstract
Background
Diabetes mellitus represents a significant health challenge that affects the broader population. A small tree, Commiphora gileadensis (C. gileadensis), is native to the Middle East.
Purpose
This research investigated the impact of C. gileadensis on blood glucose levels, HbA1c, GLUT2, GLUT4, and lipid profile.
Materials and Methods
Sixty male mice were divided into six groups: negative control, untreated diabetic, and three groups of diabetic mice treated with C. gileadensis sap, methanolic extract, acetone extract, and the metformin-treated group. Random blood glucose levels, HbA1c, lipid profile, GLUT2, and GLUT4 were assessed for each mouse across the five groups.
Results
The random blood glucose levels, triglycerides, total cholesterol, and HbA1c were markedly decreased in mice administered with C. gileadensis extracts in comparison to the untreated diabetic group. Serum and hepatic GLUT2 were markedly increased in the metformin-treated group compared to untreated diabetic and diabetic treated with C. gileadensis. Serum and muscle GLUT4 levels were elevated in diabetic mice that were administered with the three C. gileadensis extracts relative to the untreated diabetic group.
Conclusion
This study concludes that C. gileadensis sap and its methanol and acetone extracts lower serum random blood glucose levels, HbA1c, and lipid profiles in mice with type 2 diabetes mellitus. Moreover, C. gileadensis extracts may reduce serum random blood glucose levels and HbA1c in diabetic mice via inducing muscle GLUT4.
Introduction
Diabetes mellitus is a systemic metabolic condition characterized by long-term hyperglycemia, which may be induced by abnormalities in insulin production and/or action (Kim, 2019; Ozougwu et al., 2013). Diabetes dramatically raises the risk of stroke, amputation, kidney failure, and blindness; these consequences lead to significant morbidity and early mortality (André, 2021; Joshua et al., 2022). Despite the abundance of different oral hypoglycemic agents and insulin, effective treatment for diabetes remains out of reach (Shidlovskaya & Navalkivska, 2020). Diabetes mellitus has become a major healthcare issue in the twenty-first century (Chaudhury et al., 2017). Glucose is essential for all cells in the body, necessitating the presence of glucose transporters (GLUTs). As a result, all cells exhibit these essential proteins on their surface. The type and physiology of different GLUTs have been clarified by genetic investigations. These transporters can be broadly divided into two types: facilitated diffusion GLUTs and sodium-glucose linked transporters (SGLTs), which are further differentiated into a large number of isoforms. Each isoform of the GLUT has a distinct function in glucose metabolism, dictated by its tissue expression pattern, substrate selectivity, transport kinetics, and controlled expression under various physiological situations. GLUTs have garnered significant interest as potential therapeutic targets for a range of diseases. Insulin resistance describes a condition of diminished responsiveness of peripheral tissues to increasing insulin levels in the bloodstream, leading to persistently increased blood glucose concentrations. This hyperglycemic state is recognized as a defining feature of type 2 diabetes mellitus (T2DM), a major health problem marked by a rise in peripheral insulin resistance, followed by the death of β-cells and low insulin levels. Insulin resistance in T2DM is characterized by the decreased ability of GLUT4 in skeletal muscle to utilize extracellular glucose. The binding of insulin to its receptors accelerates the passage of glucose into cardiac muscle, skeletal muscle, and adipose tissue. This is done by quickly moving GLUT4 transporter vesicles to the plasma membrane and stopping the production of glucose in the liver. The integration of these regulatory pathways enhances the elimination of glucose from the bloodstream (Alexandra & Al-Hasani, 2020). The main GLUT family member, GLUT2, is present in hepatocytes and β-cells of the pancreas as well as in the kidney, intestines, and central nervous system. GLUT2 controls the predominant glucose uptake in hepatocytes according to the blood glucose level (Im et al., 2006). The main GLUT in skeletal muscle, GLUT4, is thought to be a rate-limiting factor for glucose uptake and metabolism, especially when the muscle is at rest (Deshmukh et al., 2015). Knockout GLUT4 mice, specifically in the muscle, developed a diabetic phenotype with systemic insulin resistance (Wang et al., 2017), while mice with normal and genetically diabetic db/db mice showed improved glucose tolerance and insulin sensitivity when GLUT4 was overexpressed (Lamb et al., 2010). Recent decades have seen significant advancements in therapeutic approaches for diabetes mellitus. Anti-diabetic medications may result in serious outcomes, such as hypoglycemic coma, and liver and kidney diseases (Alexandra et al., 2018; Eddouks et al., 2009). Herbal medicine is an expanding field within the medical sciences that warrants further examination. For millennia, plants have been essential in protecting humans and enhancing the quality of human life. Furthermore, they have been used as significant medicinal elements (Ali et al., 2015; Esther et al., 2020; Safaet et al., 2022). The primary therapeutic strategy involves the use of plant saps and extracts or their bioactive components (Prabu et al., 2019). Many plant species are used to treat diabetes mellitus worldwide (Ramesh et al., 2017; Wesam et al., 2016). These medicinal herbs offer oral hypoglycemic components for novel pharmaceutical formulations and dietary supplements (Alsherif, 2019; Hira et al., 2018). Commiphora gileadensis has a historical use in the treatment of various ailments (Bahare et al., 2019). C. gileadensis is broadly distributed across the Arab world, particularly in Saudi Arabia, and is found in proximity to the Sarawat Mountains in western Saudi Arabia (Hassan et al., 2023). Arabic designations for C. gileadensis include balm of Makkah, basin, and bisham. Hellenistic and Roman cultures employed C. gileadensis balsam as a fragrance (Al-Eisawi & Al-Ruzayza, 2015). Multiple studies have utilized an aqueous extract of this plant for its analgesic, diuretic, and anti-hypertensive properties (Khalid et al., 2023). In vivo and in vitro experiments have demonstrated that the C. gileadensis sap exhibits anti-bacterial properties (Al-Hazmi et al., 2020). A previous study indicated that the methanol extract of C. gileadensis improves wound healing (Al-Hazmi et al., 2022). However, the anti-diabetic activity of C. gileadensis, as a traditional Arabian herbal remedy, remains underexplored. Therefore, this study aims to elucidate the hypoglycemic efficacy and molecular mechanisms of C. gileadensis extracts in mice with T2DM by measuring blood glucose, HbA1c, and GLUT2/GLUT4 expression levels.
Materials and Methods
The present study was conducted from March to August 2024 at Taif University’s Faculty of Applied Medical Sciences.
Commiphora gileadensis Collection
In April 2024, fallen branches and leaves of C. gileadensis were gathered from Alaab Valley, a high mountain region close to the Al-Madinah region of western Saudi Arabia.
Commiphora gileadensis sap Production
Following the excision of C. gileadensis branch tips measuring 5 mm, sap was collected promptly. The sap underwent centrifugation for 10 min at 10,000 rpm, preceded by 15 min of agitation at room temperature after 50 g of the sap was diluted in ethanol. The supernatant was subsequently stored at −20°C until analysis (Eitan et al., 2015).
Commiphora gileadensis Methanol Extract Preparation
Following washing with tap water, the branches and leaves of C. gileadensis were dried in a hot-air oven at 40°C. Contaminants were removed through sieving, and the material was subsequently finely ground into powder. In a sterile funnel, 10 g of the powder was macerated for 24 h in 100 mL of 100% methanol. Before the extract was filtered through sterile filter paper, the funnel was vigorously shaken. The C. gileadensis extract was concentrated by drying in a water bath at 40°C and was then stored at 4°C for 2 weeks (Al-Hazmi et al., 2020).
Commiphora gileadensis Acetone Extract Formation
Following a 6-h drying period at 60°C in a vacuum oven, branches and leaves of C. gileadensis were pulverized with a razor blade. For 3 days, 10 g of raw C. gileadensis had been immersed in 200 mL of acetone at room temperature. The acetone was replaced every day using a magnetic stirrer. A rotary evaporator was utilized to eliminate the acetone residue from the extracted material. Then, the extract was stored at −20°C until analysis (Suliman, 2015).
Experiment Design
Sixty BALB/c male mice, with 20–25 g body weight and an age of 2 months, were housed in a well-ventilated standard rodent cage with woodchips, at a temperature of 25°C on a 12-h light/dark cycle. Throughout the trial, every mouse received tap water and a typical mouse meal. After 2 weeks of acclimation, these mice were divided into six groups at random.
The first group was designated as the negative control group. There was no intervention for the mice in this group.
T2DM was induced in mice in the second, third, fourth, fifth, and sixth groups by feeding them a high-fat (45%) and high-carbohydrate (35%) diet. Two months later, all mice showed fasting blood glucose ≥126 mg/dL (Sardi et al., 2020).
The second group, known as the positive control group, consists of untreated diabetic subjects. The third group was designated as the diabetic C. gileadensis methanolic extract-treated group. T2DM was induced in the mice, similar to the second group. Via intragastric gavage, after the establishment of T2DM, the mice received 200 mg/kg body weight per day of C. gileadensis methanolic extract (Asmaa et al., 2023). The fourth group comprised diabetic subjects that were administered with the acetone extract of C. gileadensis. T2DM was induced in the mice of this group in a manner analogous to that of the second group. Furthermore, the mice were treated with the acetone extract of C. gileadensis at the same dosage as the third group. The diabetic C. gileadensis sap-treated cohort was identified as the fifth group. All of the mice in this cohort received C. gileadensis sap at the same dosage as the third group after the start of type 2 diabetes. The sixth group was designated as the diabetic metformin-treated group. The diabetic mice in this group were treated with 200 mg/kg of body weight per day via intragastric gavage.
The mice in all six groups, after induction of T2DM, were fed a standard diet for rodents and had unrestricted access to tap water. Random blood glucose levels were estimated at 5-day intervals for all mice included in this study. At the end of the experiment, blood samples were collected to estimate HbA1c and lipid profiles.
Sample Characterization of Commiphora gileadensis Methanolic Extract Using Ultraperformance Liquid Chromatography Coupled with Mass Spectrometer
The ACQUITY UPLC BEH C18 (100 × 2.1 mm × 1.7 µm) column with Ultimate 3000 LC makes up the LC system. Solvents A (60% acetonitrile + 40% H2O + 10 mM ammonium formate) and B (10% acetonitrile + 90% isopropyl alcohol + 10 mM ammonium formate) make up the mobile phase, which has a gradient elution (0–10.5 min, 30%–100% B; 10.5–12.5 min, 100% B; 12.5–12.51 min, 100%–30% B; 12.51–16.0 min, 30% B). The mobile phase has a flow rate of 0.3 mL min−1. The sample manager temperature is set at 4°C, while the column temperature is kept at 40°C (Al-Hazmi et al., 2022).
Body Weight Measurement
This study used a digital balance (Model: Scout Pro SPU601, Ohaus, China) to measure each mouse’s body weight for 4 months. All mice in the five groups were weighed from the first day of the experiment and subsequently on day 1 and 1, 2, 3, and 4 months following treatment with various C. gileadensis extracts.
Blood Collection
Blood glucose levels were estimated at 3-day intervals after treatment with C. gileadensis extracts. At the end of the experiment, blood samples were drawn from each mouse to estimate HbA1c, lipid profile, GLUT2, and GLUT4 levels. Immediately, every blood sample was centrifuged for 15 min at 2,500 rpm. Before analysis, blood sera were stored at −80°C.
Preparation of Liver and Skeletal Muscle Homogenates
A kit with 2 mL beads was used to create homogenates of skeletal muscle and liver (OMNI International, Inc.). In a 2 mL microtube, a piece of the organ was combined with radioimmunoprecipitation assay buffer that contained a protease inhibitor.
For 2 min, the microtube was homogenized using a Read Roptor 12 machine (OMNI International, Inc.). A Sigma 1–14 k centrifuge was then used to centrifuge the microtube for 30 min at 8°C and 15,000 rpm. After the supernatant was aspirated carefully into a fresh microtube, it was centrifuged for 15 min at 15,000 rpm at 8°C. After being aspirated, the supernatant was stored at −20°C.
Estimation of Fasting Blood Glucose Levels, HbA1c, Serum Fasting Insulin Levels, and HOMA-IR Estimation
Fasting blood glucose levels were assessed using a colorimetric assay. All mice in this study underwent HbA1c testing at 4 months utilizing a glycohemoglobin kit from POINTE Scientific Inc., USA. Furthermore, serum insulin levels were measured for each mouse included in the study using a sandwich immunoassay (Invitrogen, Thermo Fisher Scientific). Homeostatic model assessment for insulin resistance (HOMA-IR) was conducted using the final fasting blood glucose level with the formula (fasting glucose (mg/dL) × fasting insulin (mU/L)/405 (for SI units: fasting glucose (mmol/L) × fasting insulin (mU/L)/22.5).
Estimation of Liver and Kidney Function Tests
Alanine aminotransferase (ALT) and aspartate aminotransferase were estimated for the mice in five groups. Serum creatinine and urea levels were estimated for all groups.
Estimation of Plasma GLUT2 and GLUT4 Levels
Competitive enzyme immunoassay was used to assess plasma GLUT2 and GLUT4 (Shanghai Ideal Medical Technology, China). The test has remarkable specificity for GLUT2 detection and has a sensitivity of 0.1 ng/mL. There was no observable cross-reactivity or interference between GLUT2 and its analogues. The intra-assay precision is defined as CV% < 5%, and the inter-assay precision is defined as CV% < 6%.
Estimation of Hepatic GLUT2
Using a competitive enzyme immunoassay, the amount of GLUT2 in the liver homogenate supernatant was determined (Shanghai Ideal Medical Technology, China). This test offers a high specificity for GLUT2 detection and a sensitivity of 0.1 ng/mL. There was no discernible cross-reactivity or interference between GLUT2 and its analogs.
Estimation of Skeletal Muscle GLUT4
GLUT4 in skeletal muscle was measured using a competitive enzyme-linked immunosorbent assay technique (Shanghai Ideal Medical Technology, China). The sensitivity and specificity of the test are equal to those of liver GLUT2 estimation.
Statistical Analysis
Prism software version 10 (GraphPad) was used for the statistical analysis. The mean ± standard deviation was used to express all data. Comparisons of weight, HbA1c, lipid profile, GLUT2, and GLUT4 between diabetic treated groups and the untreated diabetic group were made using a one-way analysis of variance (ANOVA) followed by a post hoc test (Tukey’s HSD test) to determine which groups have significant statistical differences. The comparison between random blood glucose levels in the same group throughout the experiment was done with the Wilcoxon signed-rank test. Furthermore, a two-way ANOVA was used to compare blood glucose levels over 21 days in the same group. The level of significance was set at p < .05 in comparison between the diabetic treated groups and the untreated diabetic group.
Results
Liquid Chromatography–Mass Spectrometry of C. gileadensis Methanolic Extracts
The liquid chromatography–mass spectrometry (LC–MS) chromatographic profile in negative mode indicated the presence of several lipid components. The predominant lipid components that were identified include ceramide at 69%, hexosylceramide at 18%, and phosphatidylethanolamine at 7.6%. Additionally, minor components were observed, such as dimethylphosphatidylethanolamine at 2% and phosphatidic acid at 0.97%, among others. Specifically, elevated quantities of monounsaturated fatty acids are present in ceramide, hexosylceramide, and phosphatidylethanolamine, with only hexosylceramide exhibiting a minimal quantity of polyunsaturated fatty acids.
Body Weight
In the 10th week, the diabetic groups treated with methanol extract, acetone extract, and sap exhibited significantly greater weights compared with the untreated diabetic group (p < .05).
Fasting Blood Glucose Levels
Fasting blood glucose levels 2 months after C. gileadensis extract treatment are presented in Figure 1. The diabetic mice treated with C. gileadensis sap had the lowest fasting blood glucose levels of all the diabetic groups. The diabetic mice treated with C. gileadensis methanol extract, acetone extract, or sap had marked reduced blood glucose levels compared with the untreated diabetic mice (p < .0001). No statistical differences in fasting blood glucose levels were found between the groups treated with metformin and C. gileadensis extracts.
Fasting Blood Glucose Levels of Mice in the Six Groups.

HbA1c Percentages
Figure 2 shows all groups’ HbA1c percentages. Diabetic mice treated with C. gileadensis methanol extract (5.40 ± 0.61), acetone extract (5.60 ± 0.48), or sap (5.10 ± 0.25) exhibited significantly lower HbA1c values than the untreated diabetic mice (13.4 ± 1.42) (p < .05). HbA1c value was non-significantly lower in the C. gileadensis sap-treated group than in the diabetic metformin-treated group. No statistical differences in HbA1c were found between the groups treated with metformin and C. gileadensis extracts.
HbA1c Percentage of Mice in the Six Groups.

Serum Insulin Levels
The serum insulin levels and HOMA-IR of all groups are shown in Table 1. The serum insulin levels and HOMA-IR did not show significant statistical differences between all groups. All groups had no insulin resistance except the untreated diabetic group. No statistical differences in insulin levels and HOMA-IR were observed between the groups treated with metformin and C. gileadensis extracts.
Serum Insulin Levels and Homeostatic Model Assessment for Insulin Resistance (HOMA-IR) in All Groups.

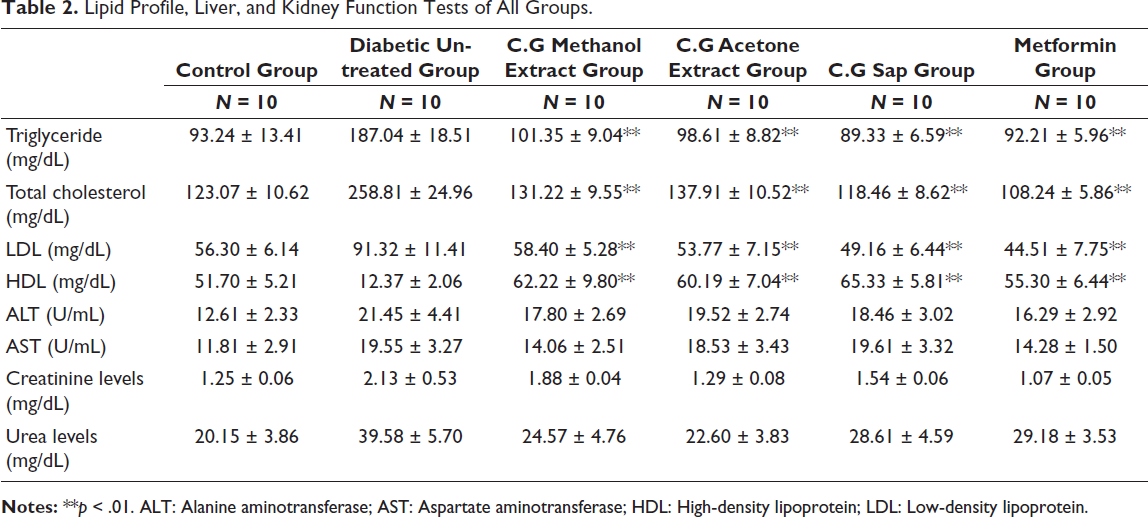
Lipid Profile, Liver, and Kidney Function Tests
The results of the lipid profile, liver, and kidney function tests are presented in Table 2. The diabetic group treated with C. gileadensis sap had the lowest triglycerides. The diabetic mice treated with C. gileadensis methanol extract (101.35 ± 9.04), acetone extract (98.61 ± 8.82), or sap (89.33 ± 6.59) had considerably lower triglycerides than the untreated group (187.04 ± 18.51) (p < .01). Diabetic mice treated with C. gileadensis sap had the lowest overall cholesterol values. Diabetic mice treated with C. gileadensis methanolic extract (131.22 ± 9.55), acetone extract (137.91 ± 10.52), and sap (118.46 ± 8.62) showed considerably reduced total cholesterol levels compared with the untreated diabetic mice (258.81 ± 24.96) (p < .01). Furthermore, the diabetic mice treated with C. gileadensis sap had significantly lower total cholesterol levels than those treated with metformin (p > .01). The low-density lipoprotein (LDL) diabetic mice treated with C. gileadensis methanol extract (58.40 ± 5.28), acetone extract (53.77 ± 7.15), or sap (49.16 ± 6.44) had significantly lower LDL levels compared with the untreated diabetic group (91.32 ± 11.41) (p < .01). The diabetic group treated with C. gileadensis methanol extract (62.22 ± 9.80), acetone extract (60.19 ± 7.04), or sap (65.33 ± 5.81) had significantly greater high-density lipoprotein (HDL) levels than the untreated diabetic group (12.37 ± 2.06) (p < .01). No statistical differences in lipid profile levels were found between metformin and C. gileadensis extracts. The liver and kidney function tests did not reveal any significant statistical differences across the groups. Furthermore, all tests are within the normal range.
Lipid Profile, Liver, and Kidney Function Tests of All Groups.
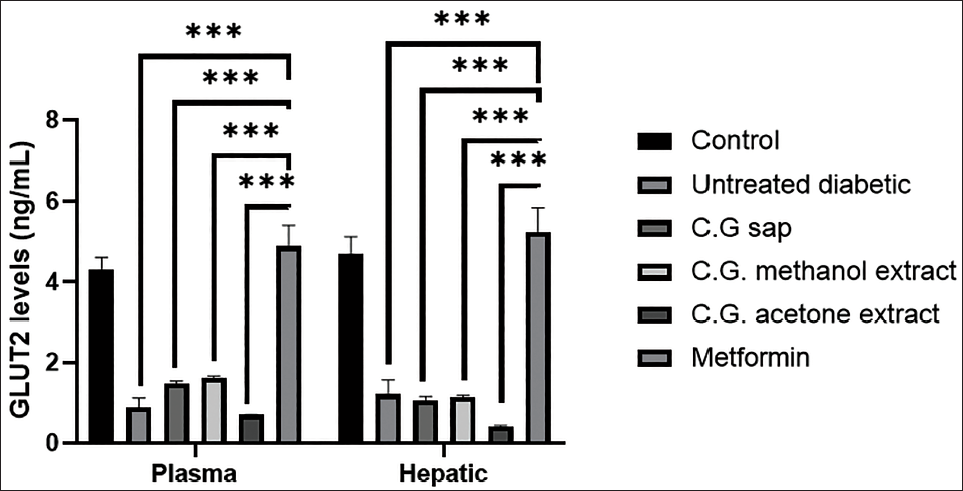
Serum and Hepatic GLUT2 Levels
Figure 3 presents the serum and hepatic GLUT2 levels of mice in all groups. The C. gileadensis extracts do not show statistically significant differences in plasma and hepatic GLUT2 in comparison to the diabetic untreated group. The plasma and hepatic GLUT2 levels were significantly increased in the metformin group compared with the untreated diabetic group (p < .001). Furthermore, the mice treated with metformin have significantly higher plasma and hepatic GLUT2 than the mice treated with C. gileadensis extracts (p < .001).
Plasma and Hepatic GLUT2 Levels of Mice in the Six Groups.
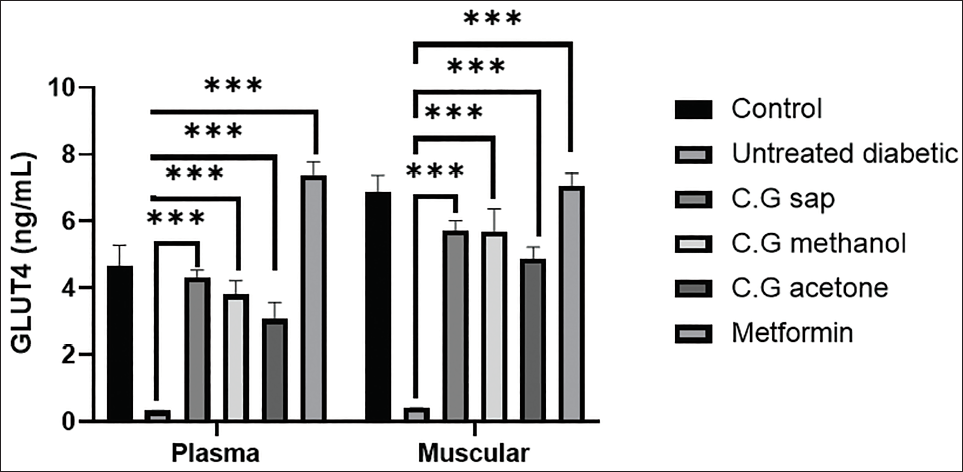
Serum and Muscular GLUT4 Levels
Compared with the untreated diabetic mice (0.41 ± 0.003), diabetic mice treated with the C. gileadensis methanolic extract (3.82 ± 0.50), acetone extract (2.97 ± 0.22), sap (4.4 ± 0.46), or metformin (7.62 ± 0.46) exhibited a significant increase in serum GLUT4 (p < .0001). Furthermore, the levels of muscular GLUT4 were significantly higher in diabetic mice that were treated with C. gileadensis methanolic extract (5.41 ± 0.32), acetone extract (4.73 ± 0.21), and sap (5.51 ± 0.32), or metformin (7.48 ± 0.31), than in the untreated diabetic mice (0.50 ± 0.002) (p < .0001) (Figure 4).
Plasma and Muscular GLUT4 Levels of Mice in Six Groups.
Discussion
About 90% of all cases of diabetes are T2DM. Insulin resistance, a disorder associated with T2DM, is a compromised body’s reaction to insulin. To sustain glucose homeostasis, insulin secretion is initially augmented to reduce its ineffectiveness, but insulin production diminishes over time, leading to T2DM. Although T2DM primarily affects people above 45 years old, rising rates of obesity, sedentary lifestyles, and energy-dense diet intake are making the disease more common in children, adolescents, and younger adults (Asmaa et al., 2023). If ideal glycemia cannot be achieved by diet and exercise, then metformin is the primary course of treatment. After metformin, numerous medications are recommended, including oral sulfonylureas and DPP-4 inhibitors. For patients with fatty liver disease, alpha-glucosidase inhibitors, pioglitazone, SGLT-2 inhibitors, and GLP-1 receptor agonists are possible medications. Recent research shows that liraglutide, a GLP-1 receptor agonist, and empagliflozin, an SGLT-2 inhibitor, prevent death and major cardiovascular events. Therefore, these drugs should be prioritized in cardiovascular disease patients. Priority should be given to treatments that do not induce hypoglycemia, including DPP-4 inhibitors, SGLT-2 inhibitors, GLP-1 receptor agonists, and pioglitazone in conjunction with metformin, as hypoglycemia is associated with elevated mortality rates. Albuminuria, blood pressure, and body weight reduction are further advantages of SGLT-2 inhibitors and GLP-1 receptor agonists (Asmaa et al., 2023). Medicinal plants or phytotherapy have been utilized economically worldwide for the prevention and treatment of different diseases, such as diabetes (Pinky et al., 2023). In the present study, liver and kidney function tests indicated that the different extracts of C. gileadensis are non-toxic. A prior study established the anti-oxidant activity of C. gileadensis, attributing it to the presence of different free radical scavengers, including saponins and flavonoids (Shah et al., 2019). In rats with type 1 diabetes mellitus, the aqueous extract of C. gileadensis lowers blood glucose levels (Al-Seeni, 2017). The current study showed that, in male mice with T2DM, the sap of C. gileadensis, along with its methanol and acetone extracts, decreased blood glucose levels. The C. gileadensis sap exhibited the most pronounced effect in lowering blood glucose levels in the shortest duration compared with the extracts derived from methanol and acetone. Furthermore, C. gileadensis sap, besides its methanol and acetone extracts, led to a reduction in HbA1c levels.
The 14 subunits that make up the GLUT2 protein are encoded by the SLC2A2 gene, located on chromosome 3 at q26.2 (Haddad et al., 2023). In hepatocytes, GLUT2 makes up over 97% of all GLUTs (Fagerberg et al., 2014). Hepatocytes’ GLUT2 is responsible for bidirectional transport in and out of the cell. A previous study revealed that GLUT2 inactivation inhibits hepatic glucose uptake, although glucose absorption in other organs persists at increased levels, hence maintaining systemic blood glucose equilibrium (He et al., 2017). Consequently, the impact on blood glucose homeostasis may be lessened if additional transporters are produced to compensate for the inactivation or decreased expression of the GLUT2 protein on the liver surface. A previous study examined the pancreatic islet-protective properties of split gill mushroom polysaccharides in rats with T2DM induced by a high-fat diet and low-dose streptozotocin. The high-dose polysaccharide group (240 mg/kg) significantly ameliorated islet pathological damage and enhanced insulin levels and GLUT2 expression, indicating its hypoglycemic effect via anti-oxidative stress mitigation and protection of pancreatic islet β-cell function (Seyer et al., 2013). The present study showed that the C. gileadnesis extracts do not affect plasma and hepatic GLUT2. The GLUT4 remains intracellular and is dynamically sorted to the plasma membrane. Insulin’s primary role is to activate GLUT4 in adipose and muscular tissues, which are essential for energy storage. Improved glucose absorption is the consequence of GLUT4 being redistributed from intracellular storage vesicles to the plasma membrane of adipose and muscle tissues. Muscle tissue glucose uptake is regulated by GLUT4, and T2DM results in a reduction in GLUT4 translocation (Matsathit et al., 2025). Patients with T2DM are unable to produce energy from glucose because of the poor expression or translocation of GLUT4 to the peripheral cell plasma membrane. In addition to drugs, a healthy diet and/or exercise can improve GLUT4 expression, concentrations, and cell surface translocation in individuals with T2DM. Additionally, a few possible artificial and natural substances that could activate the GLUT4 signaling pathways without insulin are emphasized for the efficient treatment of T2DM (Akash et al., 2018). A previous study by Assiene et al. (2025) found that the powdered leaf extracts of Corchorus olitorius L., which are rich in several families of bioactive compounds, notably polyphenols, flavonoids, and organic acids, have GLUT4 translocation activity. In the present study, C. gileadensis extracts induce muscular and plasma GLUT4, directly inducing GLUT4 because the insulin levels in the mice used in the study are normal. HOMA-IR values showed insulin resistance only in the untreated T2DM group.
Conclusion
This study concludes that C. gileadensis sap and its methanol and acetone extracts reduce random blood glucose, HbA1c, and the lipid profile in diabetic mice. Moreover, C. gileadensis extracts induce muscular GLUT4, and this may be the mechanism by which C. gileadensis reduces blood glucose levels and HbA1c in diabetic mice. C. gileadensis sap is more potent than other extracts in its anti-diabetogenic activity. A human-based study is recommended to evaluate the anti-diabetogenic effect of C. gileadensis, and an in vitro study of C. gileadensis’s effect on β-cell cultures should be conducted to determine the pathway by which this plant reduces blood glucose levels.
Footnotes
Acknowledgments
The author extends his appreciation to Taif University, Saudi Arabia, for supporting this work through project number (TU-DSPP-2024-270).
Declaration of Conflicting Interests
The author declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical Approval
The animal study protocol was accredited by the National Committee for Bioethics at Taif University (protocol code HAO-02-T-105), and the Committee considered that the proposal fulfills the requirements.
Funding
The author disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded by Taif University, Saudi Arabia, Project No. (TU-DSPP-2024-270).
Patient Consent
Not applicable.