
Abstract
Diabetes-related poor wound healing is a severe clinical challenge, causing prolonged morbidity, higher amputation risk, and high healthcare costs. Current treatment approaches are often inadequate, emphasizing the need for safe, cost-effective, multi-targeted therapies combating oxidative stress, inflammation, infection, and hyperglycemia. This study explored the diabetic wound healing potential of Solanum nigrum hydroalcoholic fruit extract (SNHE) and its molecular mechanisms employing experimental assays and network pharmacology analysis. The biological activities were evaluated using DPPH and ABTS radical scavenging tests, heat-induced red blood cell membrane stabilization for anti-inflammatory effects, α-amylase inhibition, and 2-NBDG glucose uptake in McCoy cells for antidiabetic activity. Antimicrobial efficacy was assessed using minimum inhibitory concentration (MIC) tests against Staphylococcus aureus, Bacillus subtilis, Escherichia coli, and Salmonella typhi, whereas fibroblast proliferation and migration were examined using MTT and wound-scratch assays. Zebrafish caudal-fin regeneration and streptozotocin-induced diabetic rat excision wound models were used to investigate in vivo activity. Network pharmacology was used to identify active chemicals, molecular targets, and pathways associated with diabetic foot ulcers, followed by molecular docking. SNHE demonstrated strong antioxidant properties (DPPH IC50 = 0.031 mg/mL; ABTS IC50 = 0.051 mg/mL), supressed α-amylase activity by 72.59%, enhanced glucose uptake by 245.2%, and inhibited hemolysis by 64.35%. The MIC values for SNHE ranged from 0.781–3.125 mg/mL, with the maximum sensitivity recorded for S. aureus. Effect of SNHE on McCoy cells migration resulted in 27.29% wound closure. In vivo, SNHE promoted 50.67% Zebrafish fin regeneration and 86.54% wound closure in diabetic rats. Network pharmacology revealed four active chemicals from S. nigrum—beta-ionone, ascorbic acid, citric acid, and nicotinamide—that interact with 117 DFU-related targets. Beta-ionone exhibited notable interactions with the key DFU genes AKT1, STAT3, PPARG, and PTGS2. Results suggest that S. nigrum has potential for enhancing diabetic wound healing, serving as both a useful systemic and topical therapy.
Introduction
Chronic wounds represent a substantial menace to global public health and economic stability. Individuals afflicted with diabetes mellitus (DM) are at the apex of susceptibility to the development of chronic non-healing wounds, owing to underlying patho-physiological factors encompassing hyperglycemia, neuropathy, compromised blood flow, and chronic inflammatory processes localized at the site of injury. The International Diabetes Federation (IDF) has proffered prognostic data indicating a prospective escalation in diabetic patient numbers, surging from 589 million in 2024 to 853 million by 2050, exacerbating the prevalence of chronic wounds. 1 It is noteworthy that the overall lifetime incidence of diabetic foot ulcers (DFUs) is estimated to range between 19% and 34%. Consequently, it is not unexpected that chronic wounds in the diabetes often culminate in lower extremity amputations, deleteriously affecting quality of life and necessitating judicious therapeutic interventions. 2
While a range of treatments exists for addressing impairments in wound healing, spanning from traditional wound care management to surgical interventions, recent therapeutic approaches have incorporated the utilization of antibiotics, hyperbaric oxygen therapy to optimize oxygen delivery to skin tissues, and topical dressings designed to create a moist environment optimum for stimulating the healing process. Further, topical approach of insulin, growth factors, stem cells, etc., has shown promising results. Unfortunately, the majority of these therapies have their own disadvantages, such as high cost, a risk of development of microbe resistance, and the propensity to cause allergic reactions. Therefore, it is necessary to discover and explore alternative treatments derived from natural sources which possess the potential to be considered safe, effective, and exhibit minimal adverse effects. 3 This has prompted the investigation of medicinal plants abundant in multifunctional phytoconstituents, as they can address the interconnected challenges of oxidative damage, chronic inflammation, infection control, and metabolic dysregulation that impede diabetic wound healing. 4
Plants are an excellent source of therapeutic treatments with negligible adverse side effects, low-cost, and convenient to use. 5 It has been demonstrated that administering medicinal plants to wounds is effective in preventing infections and accelerating the healing process. A significant part of the population in many developing nations practices traditional medicine for their regular medical requirements, often integrating it with modern medications. 6 Medicinal plants traditionally used for treating skin injuries and metabolic disorders may provide dual advantages in diabetic wound healing by simultaneously addressing infection, inflammation, and glycemic regulation.
Solanum nigrum, a dicotyledon belonging to the Solanaceae family, also referred to as “Black nightshade.” It is an annual plant with a maximum height of 1.25 m. S. nigrum has likely been used medicinally for over two thousand years. 7 S. nigrum's pharmacological potential is based on a wide range of phytochemicals including glycoalkaloids, glycoproteins, polysaccharides, and polyphenolic compounds, such as rutin, gallic acid, epicatechin, and caffeic acid. 8 These diverse bioactive compounds of S. nigrum collectively facilitate mechanisms crucial for wound healing in diabetes, including the mitigation of reactive oxygen species, modulation of inflammatory pathways, and inhibition of microbial growth.9,10
Traditionally, this herb has been used to treat renal irritation, herpes virus infections, cardiac conditions, ulcers and skin disorders. It also exhibits pharmacological properties that include antimicrobial, anti-dysenteric, anti-diabetic, anti-diuretic, and anti-malarial effects. 11 Additionally, the ripe fruits of S. nigrum have been used topically to promote wound healing due to their antibacterial and anti-inflammatory qualities, believed to contribute to the healing process. 12 Nonetheless, despite its extensive pharmacological range, no thorough studies have assessed the topical wound healing efficacy of S. nigrum in diabetic conditions. Moreover, there is a paucity of studies integrating experimental outcomes with network pharmacology and in silico analyses to comprehensively map its multi-target mechanisms of action, which could be essential for addressing the multifaceted pathology of diabetic wounds.
Thus, the purpose of this study was to assess the topical effectiveness of S. nigrum fruit hydro-alcoholic extract (SNHE) in managing wound healing and skin repair in diabetic condition, where the complex healing process is impaired.
Materials and methods
A flowchart illustrating the sequential methodologies used in the present study is shown in Figure 1.
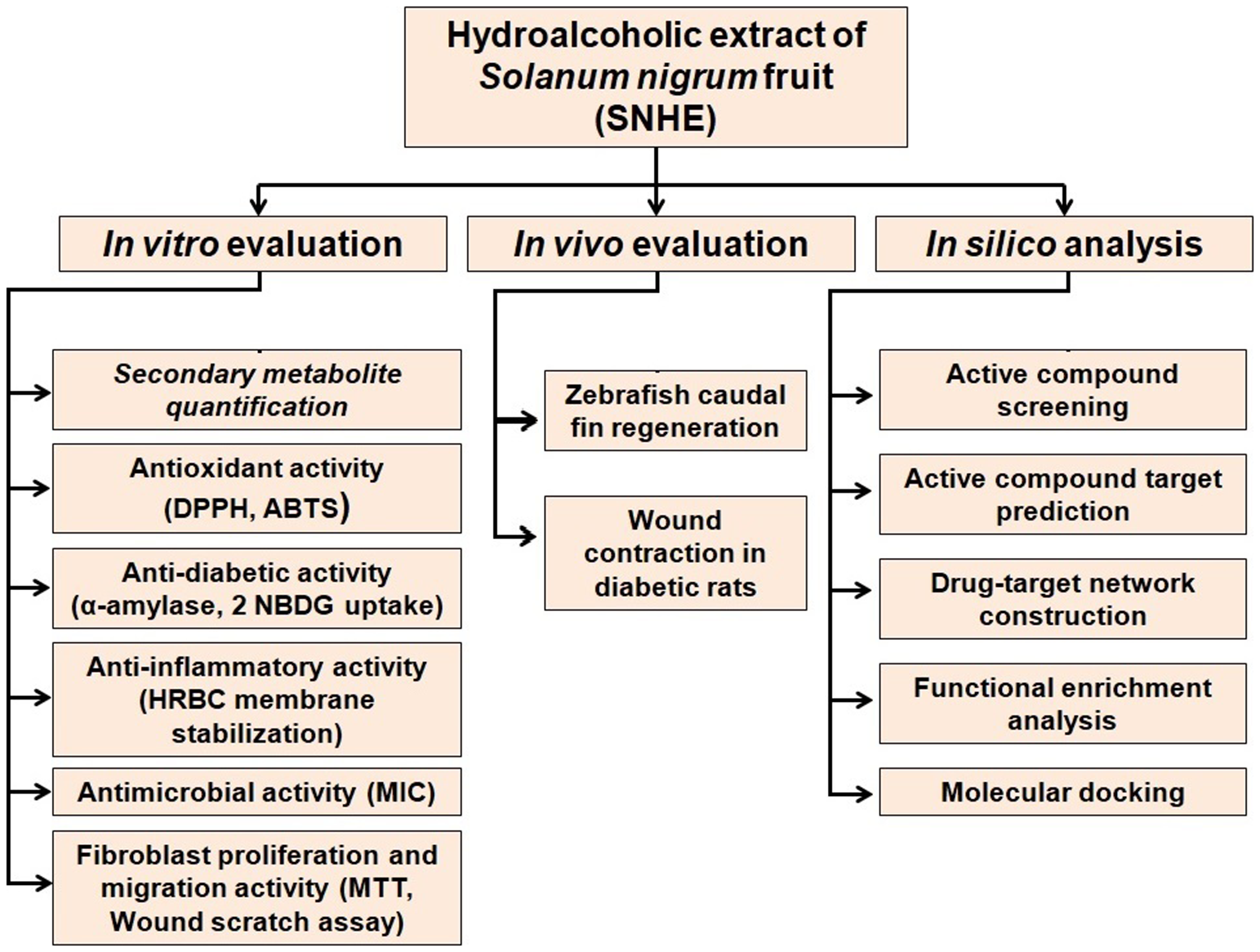
Flowchart illustrating the sequential methodology used in the study.
Materials
Himedia supplies included Dulbecco's modified eagle medium (DMEM), 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide (MTT), penstrep, Fetal bovine serum (FBS), Mueller-Hinton broth (MHB), and 96-well bottom plates. 2-(N-(7-Nitrobenz-2-oxa-1,3-diazol-4-yl)Amino)-2-Deoxyglucose (2-NBDG) Molecular Probes were procured from Invitrogen, while Sigma Aldrich provided other chemicals. McCoy fibroblasts were procured from the National Centre for Cell Science, Pune, India and maintained in DMEM with 10% FBS and 1% penstrep at 37°C with 5% CO2.
Quantification of phenolic and flavonoid content
The Folin-Ciocalteau assay was utilized to quantify the total phenolic content of SNHE, with Gallic acid serving as the standard reference. 13 The aluminium chloride calorimetric method was used to determine the total flavonoid content of SNHE. 14
Anti-oxidant activity of SNHE
In order to assess the antioxidant activity of SNHE, the DPPH and ABTS radical scavenging tests were employed. The standard calibration curve for the DPPH assay was established using various concentrations of butylated hydroxytoluene (BHT). 15 Furthermore, the ABTS technique was also employed to evaluate the antioxidant potential of SNHE. 15 The standard calibration curve for this assay was established using a range of ascorbic acid concentrations.
Inhibition percentage for DPPH and ABTS inhibition was calculated as follow:
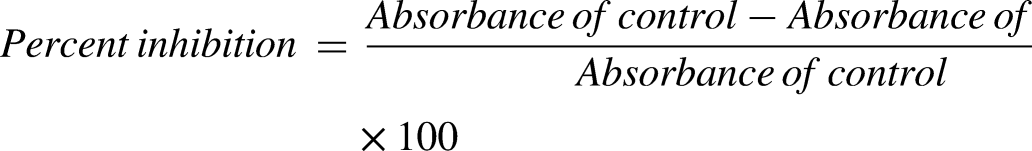
α-amylase inhibitory assay
The DNSA method was used to examine the anti-diabetic properties of SNHE, with acarbose serving as a positive control. 16 Different concentrations of SNHE were mixed with 2 units/mL of α-amylase and incubated at 30°C for 10 min. After that, starch and DNSA reagent were added, and resultant mixture was subjected to boiling. After that, the absorbance at 540 nm was measured.
The α-amylase inhibitory activity was expressed as percent inhibition and was calculated as follow:

Glucose uptake in McCoy fibroblasts
McCoy fibroblasts were cultivated in 96-well plates and subjected to an overnight treatment with 0.1 mg/mL of calendula cream and 0.05 and 0.1 mg/mL of SNHE. Afterwards, fresh medium containing 100 μM of the fluorescent glucose derivative 2-NBDG was added to the cells and incubated for an hour. Following a wash with cold phosphate buffer saline (PBS), fluorescence was observed and analyzed using a fluorescent microscope (Nikon Eclipse Ti-U) and ImageJ software (National Institutes of Health, USA).
Human red blood cell (HRBC) membrane stabilization study
The in vitro anti-inflammatory activity of SNHE was evaluated by HRBC membrane stabilization assay.
17
A reaction mixture consisting of 50 µL of SNHE, 50 µL of erythrocyte suspension (10% v/v), and 2.9 mL of PBS was incubated at 54°C for 20-min. Afterward, it was centrifuged at 2500 rpm for 3 min. The optical density of the resulting supernatant was measured at 540 nm, with Aspirin serving as the positive control. The percent protection was calculated using the provided formula:
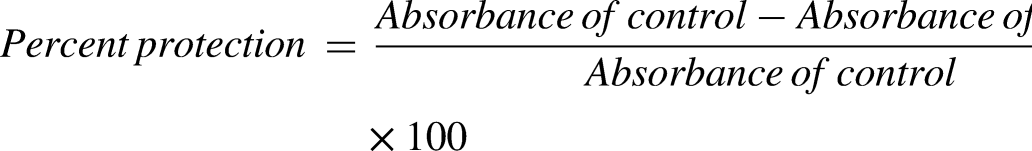
Minimum inhibitory concentration (MIC) of SNHE
The potency of SNHE against common wound infection bacteria (Staphylococcus aureus MTCC3160, Bacillus subtilis MTCC121, Escherichia coli ATCC25922, and Salmonella typhi MTCC98) was determined in accordance with CLSI (2012) guidelines. 18 The broth micro-dilution method was used to determine the MIC of SNHE. In sterile 96-well plates containing MHB, serial dilutions of SNHE were used along with bacterial suspension. After overnight incubation at 37°C, 0.04% w/v Resazurin dye was added to detect bacterial growth.
Effect of SNHE on fibroblast proliferation and migration
The MTT assay was used to measure cell growth. 19 In a 96-well flat bottom plate with 0.12% FBS-supplemented DMEM, McCoy cells (104 cells/well) were seeded and exposed to different SNHE concentrations. After 24 h, MTT (5 mg/mL) was added, followed by 4 h incubation at 37°C. Absorbance was measured at 580 nm.
The wound scratch assay was employed to measure the effect of SNHE on cell migration. 19 Using a sterile 100 µL pipette tip, linear wounds were created in McCoy fibroblast monolayers. Treatments administered to cells consisted of control (no treatment), Calendula cream (0.1 mg/mL, positive control), and SNHE (0.05 mg/mL and 0.1 mg/mL). For the purpose of evaluating migration, pictures of scratched areas were taken after 24 h of incubation at 37°C. ImageJ software was used to analyze image data.
In vivo diabetic wound healing study
SNHE was evaluated in vivo for its ability to promote fin regeneration in Zebrafish and heal diabetic wounds in Wistar rats, with Calendula cream serving as a control. In order to evaluate the effectiveness of SNHE in treating wounds, adult Zebrafish were employed as a model system.
20
After two weeks of acclimatization, adult Zebrafish were divided into control, standard, and experimental groups. A hyperglycemic environment was generated by exposing the fish to a 2% glucose solution, and the fish were anaesthetized for a 5 mm caudal fin transection. For seven days, SNHE (2 and 4 mg) and Calendula cream (4 mg) were applied to fish. Fin regeneration was monitored and calculated as:
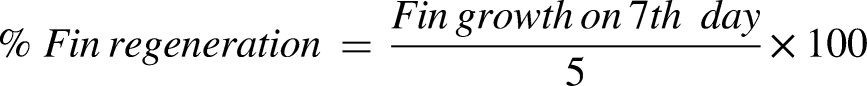
Furthermore, by administering streptozotocin intraperitoneally at a dose of 70 mg/kg body weight, diabetes was induced in Wistar rats, resulting in high plasma glucose levels (200 mg/dL). On the dorsal region of rats, a 5 mm wound was generated. For a period of 14 days, treatments were administered once daily to rats, including water as the negative control, Calendula cream (10 µg) as the standard, and SNHE (10 µg). Wound healing assessment involved the measurement of wound areas.
21
Wound closure percentages were calculated as:
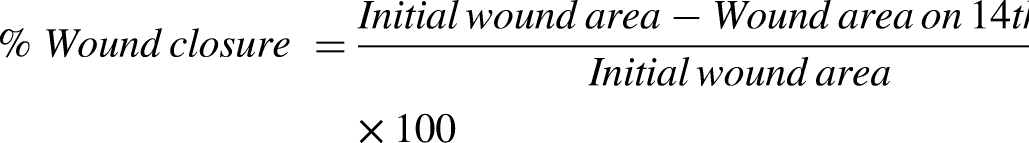
Molecular mechanism of SNHE on DFUs using network pharmacology
Pharmacokinetic screening of active compounds of S. nigrum: The phytoconstituents of S. nigrum were extracted from IMPAT (https://cb.imsc.res.in/imppat/home) and KNApSAcK (http://www.knapsackfamily.com/KNApSAcK) databases. SwissADME (http://www.swissadme.ch), molsoft (https://www.molsoft.com/mprop), and protox II (https://tox-new.charite.de) were used to evaluate the pharmacokinetic parameters, such as oral bioavailability (OB) ≥ 30%, drug-likeness (DL) ≥ 0.18, and toxicity of chemical constituents obtained from the IMPAT and KNApSAcK, in order to identify the bioactive compounds of S. nigrum. The rate, percentage, and extent of an oral medication that reaches the anticipated location of therapeutic action are measured by OB, an important factor in drug research. According to Ononamadu et al., 2021, DL is an indication that is used to determine whether a compound has the potential to be turned into a medicine based on its chemical and physical features. 22
Prediction of targets of active compounds
The targets of the active molecules of S. nigrum were identified through the Swiss Target Prediction database (http://swisstargetprediction.ch) and the STITCH database (http://stitch.embl.de).23,24
Genes associated with DFU
The term “diabetic foot ulcer” was employed as the primary keyword in the GeneCards (https://www.genecards.org/) and OMIM (https://www.omim.org) databases to identify and screen relevant targets in humans.25,26 The targets obtained from these databases were integrated to eliminate duplicate results, resulting in a compilation of potential DFU targets. Common genes associated with active compounds of S. nigrum and DFU were identified using a Venn Diagram (https://bioinformatics.psb.urgent.be/webtools/Venn/).
Network construction
Cytoscape version 3.10.1 (https://cytoscape.org) was employed to construct a drug target network, with efforts made to minimize duplication. 27 To identify hub genes and establish a targets-protein-protein interaction (PPI) network, the STRING database (https://string-db.org) was utilized to generate potential targets. 28 According to Tai et al., 2022, hubs were identified in the analysis as nodes having degrees that were twice or higher than the mean node degree. 29
Gene ontology (GO) enrichment and KEGG analysis
The targets associated with DFU identified from the hub genes related to “homo sapiens” were used for GO enrichment and KEGG analysis using the DAVID resource system with a threshold of p ≤ 0.05. 30
Molecular docking
Receptor and ligand preparation: Three-dimensional (3D) structure of the top 4 genes obtained from PPI-network was obtained from the Protein Data Bank database (www.rcsb.org). The 3D structures of bioactive phytocompounds present in S. nigrum were downloaded in SDF format from Pubchem (https://pubchem.ncbi.nlm.nih.gov/) and converted to PDB format using Open Babel (http://openbabel.org/).
Molecular docking: The selected ligands (phytocompounds) were docked with the receptor hub proteins- Serine/threonine-protein kinase AKT (AKT1), Signal transducer and activator of transcription 3 (STAT3), Peroxisome proliferator-activated receptor gamma (PPARG), and Cyclooxygenase-2 (PTGS2) using the AutoDock Vina Software (https://vina.scripps.edu).
The binding mode that had the highest docking score in our molecular docking analysis was chosen to examine the interaction between the protein receptor and its anticipated active chemicals. The grid box was centered to cover the active binding site and all essential residues. For the AKT1, grid box (46 Å × 58 Å × 54 Å) centered at (11.99, −13.67, −13.79) Å, for the STAT3, grid box (126 Å × 60 Å × 126 Å) centered at (0.23, 28.88, 33.56) Å, for the PPARG, grid box (40 Å × 40 Å × 40 Å) centered at 55.17, −27.91, 19.65) Å, for the PTGS2, grid box (76 Å × 94 Å × 90 Å) centered at (33.269, −4.946, 7.759) Å. The structural interactions between receptor and ligand, as well as molecular visualization, were examined using the BIOVIA Discovery Studio Visualizer.
Statistical analysis
The experiments were conducted with three replicates each, and the data were presented as the mean ± standard deviation (S.D.). Data analysis was carried out using SPSS version 20.0. To evaluate the statistical significance between groups, the t-test was applied, and significance was determined at the P < 0.05 level.
Results and discussion
Phenolic and flavonoid content in SNHE
Using calibration curves generated using gallic acid and quercetin standards, the phenolic and flavonoid contents of the SNHE were determined. Using the equation y = 0.0031x + 0.064 from a standard curve with an R2 of 0.997, the phenol analysis of SNHE revealed a concentration of 68.30 mg of gallic acid equivalents (GAE)/g of SNHE. A calibration curve (R2 = 0.9908) was used to determine the flavonoid content of SNHE. The results were expressed as quercetin equivalents (QE), with a total of 62.2 mg QE/g of SNHE. Phenolic and flavonoid molecules constitute key secondary metabolites in medicinal plants, widely known for their antioxidant, antibacterial, and anti-inflammatory activities. Together, they support tissue repair by promoting angiogenesis, re-epithelialization, collagen synthesis, and modulation of inflammatory responses through multiple mechanisms and signaling pathways, such as Wnt/β-catenin, TGF-β, NF-κB, MAPK/ERK, and PI3K/Akt.31,32 In the management of chronic wounds, where infection and oxidative stress are major challenges to healing, this multi-targeted activity becomes highly essential. Our results are consistent with phytochemical profiling of S. nigrum reported in studies from Saudi Arabian and Nigerian, where bioactive constituents like γ-tocopherol, β-sitosterol, and different fatty acid esters were found along with high total phenolic content and total flavonoid content.33,34 As a rich source of medicinal phytochemicals, S. nigrum has consistently surpassed other Solanum species, including Solanum cordatum and Solanum schimperianum, in both phenolic and flavonoid content. The pharmacological significance of the higher phenolic and flavonoid levels found in SNHE emerges from the fact that polyphenols have been demonstrated to mitigate oxidative damage in chronic wounds by counteracting reactive oxygen species through electron or hydrogen atom donation. 35 Furthermore, certain flavonoids and phenolics have antibacterial properties against pathogens associated with wounds, aiding to prevent infections and promote faster tissue regeneration.
Anti-oxidant activity of SNHE
Oxidative stress contributes significantly to the chronicity of diabetic wounds by surpassing antioxidant defensesleading to prolonged inflammation, impaired angiogenesis, and delayed re-epithelialization. Antioxidant-based approaches that target reactive oxygen species may aid in redox balance restoration, inflammation management, and tissue repair. 36 Anti-oxidants balance free radicals by donating electrons or hydrogen atoms. The ability of SNHE to transfer hydrogen atoms was evaluated using the DPPH and ABTS assays. The DPPH method, renowned for its simplicity and sensitivity, 37 assesses anti-oxidant activity by determining the IC50, which is the concentration required reducing the initial concentration of DPPH by 50%. DPPH radical scavenging activities for SNHE and BHT were 0.031 mg/mL and 0.026 mg/mL, respectively (Table 1). ABTS radical scavenging activity of SNHE at different concentrations, with IC50 values for SNHE of 0.051 mg/mL and ascorbic acid of 0.044 mg/mL is listed in Table 1. These findings demonstrate that SNHE possesses considerable antioxidant activity, as evidenced by IC50 values that are comparable to those of standard antioxidants— ascorbic acid in the ABTS assay and BHT in the DPPH assay. Both studies displayed a dose-dependent increase in radical scavenging activity, thereby suggesting a potent potential for electron or hydrogen atom transfer. This activity is most likely due to SNHE's high phenolic (68.30 mg GAE/g) and flavonoid (62.20 mg QE/g) content, which can neutralize free radicals via hydrogen donation and resonance stabilization. These results align with previous studies that demonstrated the strong in vitro antioxidant activity of S. torvum and S. nigrum extracts, with solvent selection affecting potency. 38 Furthermore, study on the leaves of S. nigrum by Lim et al. (2025) showed that solvent–water combinations greatly improved the extraction of phytochemicals, especially phenolics and saponins, which subsequently contributed to stronger DPPH and ABTS radical scavenging activities. 39 The high anthocyanin content of purple eggplant peel extract (S. melongena L.) further supports the antioxidant potential of phenolic-rich Solanum species. This extract demonstrated strong antioxidant activity with 74.26% inhibition in the β-carotene bleaching assay. 40
Antioxidant activity of Hydroalcoholic extract of S. nigrum determined by DPPH and ABTS scavenging assay.

Results are expressed as means ± standard deviation (n = 3).
In vitro anti-diabetic activity of SNHE
α-amylase is a metalloenzyme that breaks down polysaccharides into maltose and maltotriose, leading to increased postprandial glucose levels. Inhibiting this enzyme can slow the release and absorption of glucose, reducing postprandial hyperglycemia. 41 Therefore, the hypoglycemic effect of SNHE was evaluated by α-amylase inhibitory activity (Figure 2). The hypoglycemic effects of SNHE were dose-dependent, with 0.1 mg/mL extract exhibiting a 72.6% inhibition. Acarbose had an IC50 value of 0.042 mg/mL, whereas SNHE had an IC50 value of 0.061 mg/mL. In a previous study, in diabetic rats, an aqueous extract of S. nigrum fruit improved blood glucose, lipid profile, and vascular reactivity, potentially by repairing pancreatic β-cells or increasing Glu4 translocation. 42 Antioxidants found in the fruit extract of S. nigrum may also be contributing to the antidiabetic effect through regulating a number of enzyme activities. α-amylase, one such enzyme, increases blood sugar levels and results in type II diabetes by converting starch to glucose. Natural compounds may assist in managing diabetes by improving insulin action. 43 Hence, the suppression of α-amylase activity by SNHE could potentially contribute to diabetic wound healing by decreasing the conversion of starch into glucose. This could potentially lower high blood glucose levels, generating a more favorable environment for diabetic wound repair.
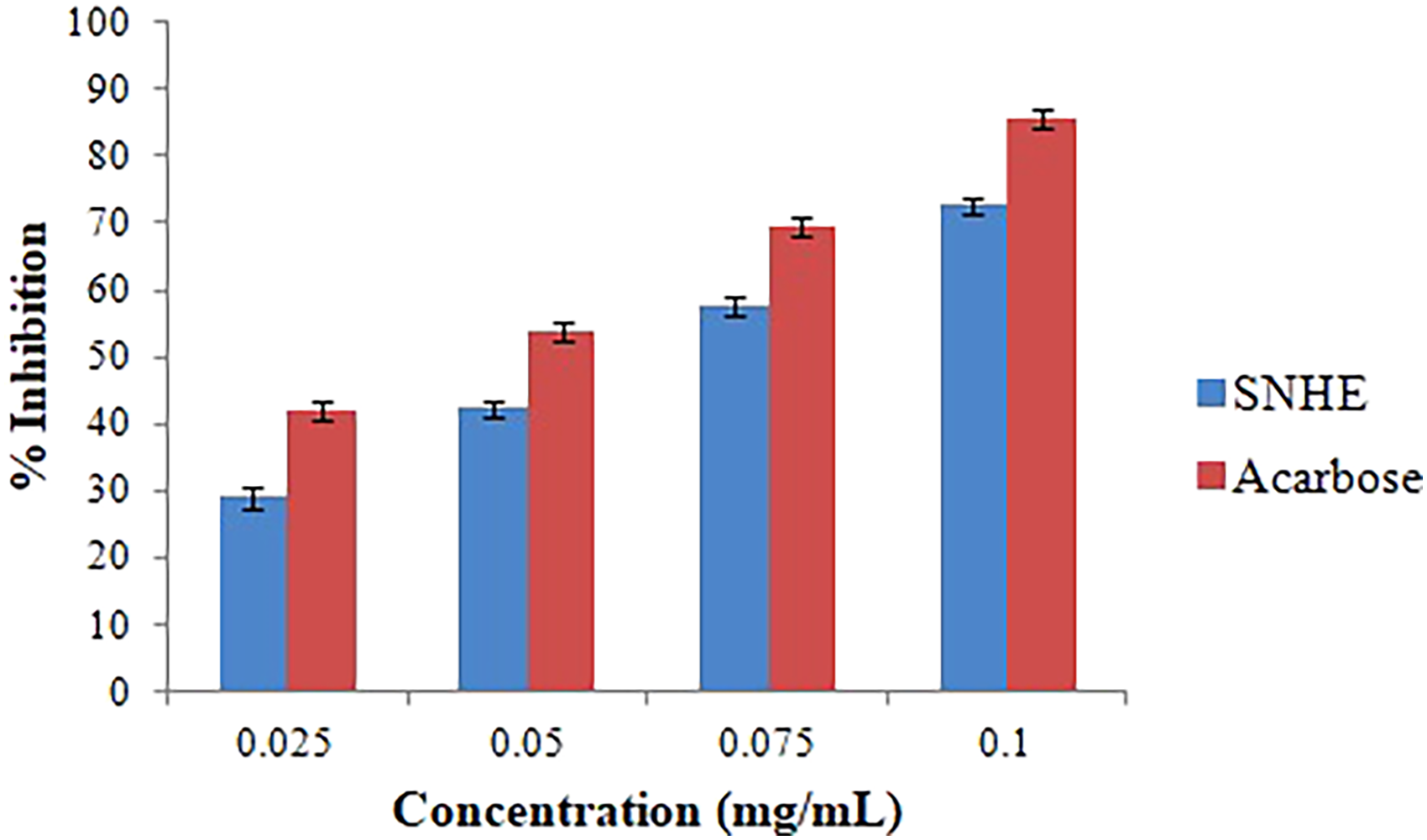
Anti-diabetic activity of SNHE: graphical representation of inhibition of alpha amylase activity of Acarbose and SNHE at various concentrations. The results were expressed as mean ± S.D. of independent replicates and error bars represent the standard deviation from the mean values. Significance was determined at the P < 0.05 level based on t-test. *SNHE- Solanum nigrum fruit hydro-alcoholic extract.
One of the fundamental metabolic dysfunctions in diabetes is reduced cellular glucose uptake due to insulin resistance, which results in persistent hyperglycemia. 44 Enhancing glucose uptake can help remove excess glucose from the bloodstream, enhance cellular energy availability, and lower glucotoxicity. The fluorescent glucose analog 2-NBDG was used to evaluate glucose uptake in fibroblast cells in order to determine the effect of SNHE on glucose uptake. SNHE increased 2-NBDG uptake in McCoy cells, as evidenced by the pronounced fluorescence observed (Figure 3(a)-(e)). Compared to untreated cells, 2-NBDG uptake was significantly enhanced in McCoy cells treated with SNHE, with increment of 1.74-fold and 2.45-fold at concentrations of 0.05 and 0.1 mg/mL, respectively. Calendula cream also exhibited a 1.47-fold increase in 2-NBDG uptake at concentration of 0.1 mg/mL. In a comparable study on another Solanum species, S. lasiocarpum, glucose uptake was enhanced in L6 myoblasts, with absorption rates reaching 118.19% at 31.25 µg/mL and peaking at approximately 132.02% at 62.5 µg/mL, compared to untreated cells. 45 Increased glucose uptake can improve cellular glucose accessibility and lower blood glucose level, potentially offering advantages in diabetes treatment. Results suggest that SNHE may present a promising therapeutic choice for diabetes management.
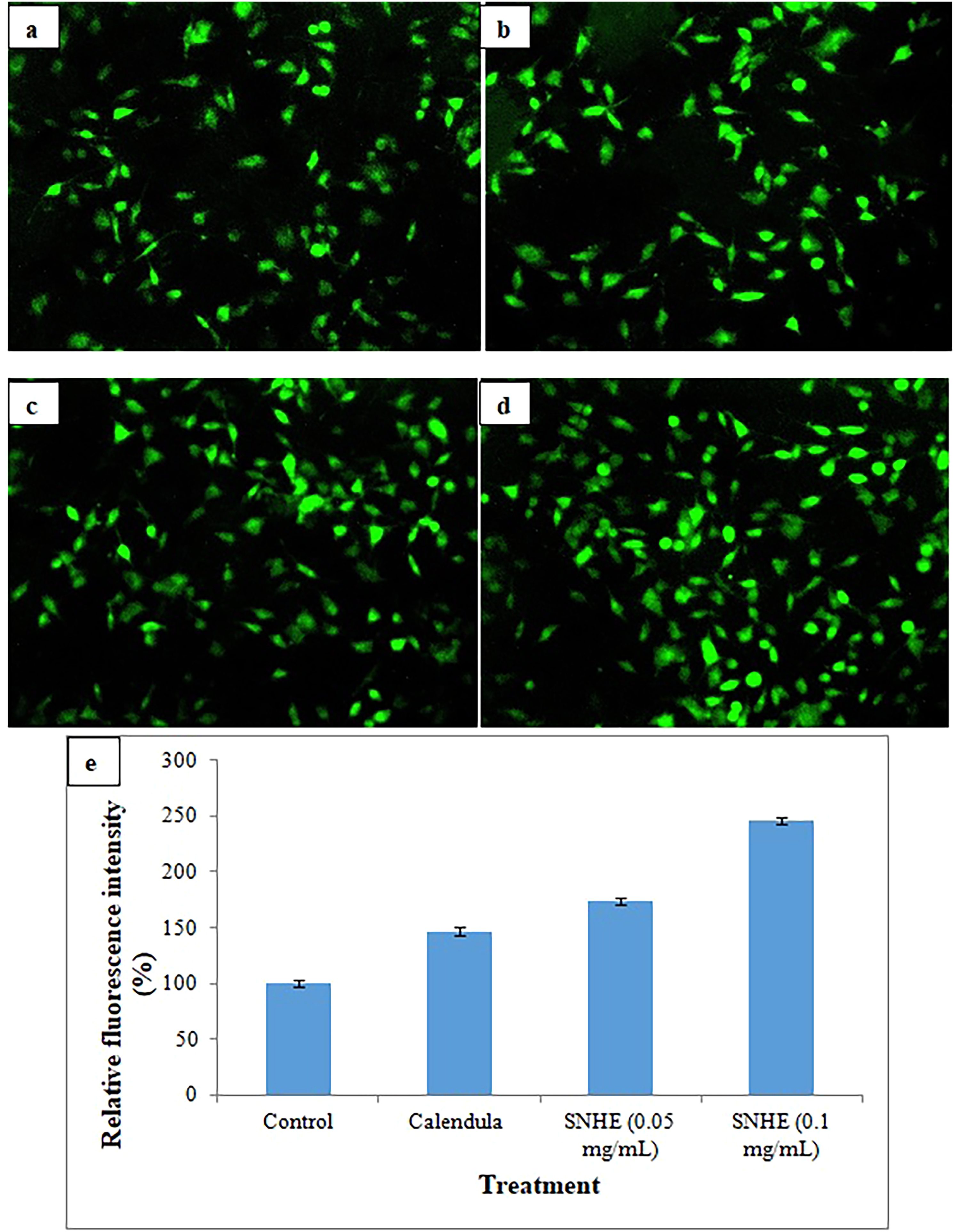
2-NBDG uptake assay in McCoy fibroblast cells by fluorescent microscopy: (a) untreated cells, (b) treated with Calendula cream, (c) treated with 0.05 mg/ml of SNHE, (d) treated with 0.1 mg/ml of SNHE, and (e) fluorescence intensity graph. The results were expressed as mean ± S.D. of independent replicates and error bars represent the standard deviation from the mean values. Significance was determined at the P < 0.05 level based on t-test. *SNHE- Solanum nigrum fruit hydro-alcoholic extract.
Anti-inflammatory activity of SNHE
The HRBC membrane stabilization assay was used to evaluate the anti-inflammatory activity of SNHE due to the structural similarities between erythrocyte and lysosomal membranes. Lysosomes play an important role in inflammation by releasing mediators leading to tissue damage. Excessive fluid inflow under hypotonic conditions results in red blood cell lysis, simulating the release of proteins and enzymes during inflammation. Preventing such lysis indicates that the extract may stabilize lysosomal membranes and inhibit the release of inflammatory mediators in vivo. 17 SNHE effectively inhibited heat-induced hemolysis. This impact might prevent neutrophil lysosomal content from being released at inflammatory sites. SNHE inhibited the heat-induced hemolysis to varying extents at various concentrations (Figure 4), with a maximal inhibition of 64.35% at 0.2 mg/mL. Aspirin, which was utilized as a control, demonstrated 77.12% inhibition. The anti-inflammatory potential of SNHE was remarkably comparable to Aspirin, with IC50 values of 0.127 mg/mL and 0.083 mg/mL, respectively. Chronic wounds are characterized by persistent inflammation, which hinders the proliferative phase of healing. This chronic inflammatory condition affects tissue repair mechanisms, contributing to slow healing of the wounds. 46 Several bioactive compounds found in S. nigrum have been demonstrated to exhibit anti-inflammatory effects. Solanine, a glycoalkaloid, can effectively reduce inflammation by restricting edema formation. Solasodine inhibits arachidonic acid metabolism through COX and 5-lipoxygenase pathways. It has been demonstrated that steroidal glycosides inhibit the formation of nitric oxide in activated macrophages, whereas flavonoids including apigenin, kaempferol, luteolin, rutin, and quercetin control inflammation by a variety of molecular mechanisms. 47 The anti-inflammatory action observed in SNHE may be attributed to these compounds. Candlenut leaf extract (Aleurites moluccanus) inhibits TNF-α and promotes wound healing in vivo, 48 comparable to the anti-inflammatory effects demonstrated with SNHE in our study.

Anti-inflammatory activity of SNHE: HRBC membrane protection/ stabilization potential of SNHE and Aspirin. The results were expressed as mean ± S.D. of independent replicates and error bars represent the standard deviation from the mean values. Significance was determined at the P < 0.05 level based on t-test. *SNHE- Solanum nigrum fruit hydro-alcoholic extract.
Anti-bacterial activity of SNHE
Infected DFUs are challenging to treat due to their polymicrobial nature and poor response to prolonged antibiotics. Chronic diabetic wounds are more susceptible to microbial colonization due to impaired progression through the physiological phases of healing. Pathogens further interfere with processes necessary for tissue repair by releasing chemicals and modifying immune responses. 49 Therefore, it is crucial to eliminate and prevent microbial growth because infections can hinder the healing process. S. aureus exhibited the highest sensitivity to SNHE, followed by B. subtilis, S. typhi, and E. coli. In comparison to ampicillin, which had MIC values between 0.019 and 0.0312 mg/mL, SNHE exhibited MIC values ranging from 0.781 to 3.125 mg/mL (Table 2). These results align with earlier reports on Solanum species antibacterial properties. Strong activity of S. nigrum against foodborne pathogens was reported by Beigomi and Valizadeh (2025), with MIC values ranging from 12.5 to 100 mg/mL, particularly effective against B. cereus and S. aureus. 50 Other Solanum species- S. anguivi, S. elaeagnifolium, S. incanum, S. xanthocarpum, S. trilobatum, and S. torvum, also exhibit significant antibacterial action against both Gram-positive and Gram-negative bacteria. 51 Likewise, with inhibition zones of up to 22 mm at 100 mg/mL and MIC values as low as 0.01 mg/mL for E. coli and 0.1 mg/mL for S. aureus, purple eggplant peel extract (S. melongena L.) displayed antibacterial activity against both Gram-positive (S. aureus) and Gram-negative (E. coli) strains. 40 Susceptibility to infections in diabetic patients can hinder wound healing. The anti-bacterial properties of SNHE can aid in preventing or treating infections in diabetic wounds, accelerating the healing process.
Anti-bacterial activity of SNHE against pathogenic bacteria.
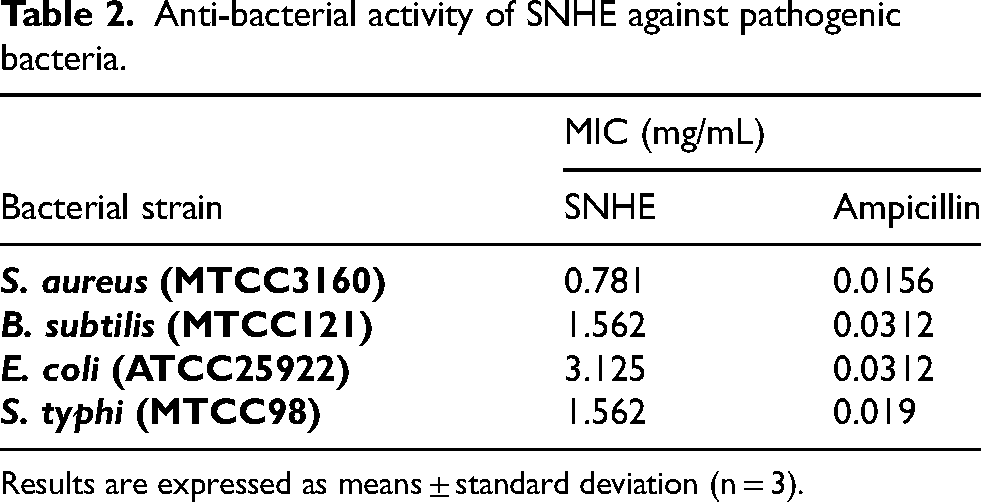
Results are expressed as means ± standard deviation (n = 3).
Fibroblast proliferation and migration
Fibroblasts play a crucial role in wound healing by migrating to the wound site and proliferating to restore tissue integrity. Their migration and proliferation, as well as the coordinated movement of fascia-resident fibroblasts, are critical for wound closure and efficient tissue repair. 52 Fibroblasts contribute to collagen synthesis by building the structural framework of new tissue. As shown in Figure 5, SNHE enhanced fibroblast proliferation in the range of 0.025–0.1 mg/mL, peaking at 0.1 mg/mL (139.12%), while Calendula cream significantly stimulated fibroblast proliferation at 0.1 mg/mL (111.6%). These findings demonstrate that SNHE, as compared to untreated and Calendula cream-treated cells, has a positive impact on fibroblast proliferation.

Effect on proliferation of McCoy fibroblast: graphical representation of the evaluation of Calendula cream and SNHE effects on McCoy fibroblast proliferation. The results were expressed as mean ± S.D. of independent replicates and error bars represent the standard deviation from the mean values. Significance was determined at the P < 0.05 level based on t-test. *SNHE- Solanum nigrum fruit hydro-alcoholic extract.
Scratch assay images on McCoy cells are shown in Figure 6(a)–(e), illustrating the progression of wound healing over a 24 h period. Compared to untreated cells, cells exposed to SNHE exhibited enhanced migration and wound closure. Calendula cream (0.1 mg/mL) treated cells demonstrated 14.3% wound closure, while SNHE-treated cells achieved wound closure rates of 9.97% and 27.29% at concentrations of 0.05 and 0.1 mg/mL. In Figure 6(f), the percentage of the wound closure area for the experimental and control groups is graphically represented. The bioactive constituents of SNHE, such as flavonoids and phenols, promote fibroblast migration and proliferation, which are essential processes in wound healing. By promoting these processes, the extract can accelerate tissue repair, enhance collagen synthesis, and facilitate faster wound closure. SNHE's phenolic and flavonoid-rich composition may enhance proliferation and migration by protecting fibroblasts from oxidative stress, stimulating TGF-β-mediated collagen synthesis, and promoting extracellular matrix remodeling.53,54 By promoting fibroblast proliferation, SNHE can hasten granulation tissue formation, improve collagen deposition, and improve early wound contraction.
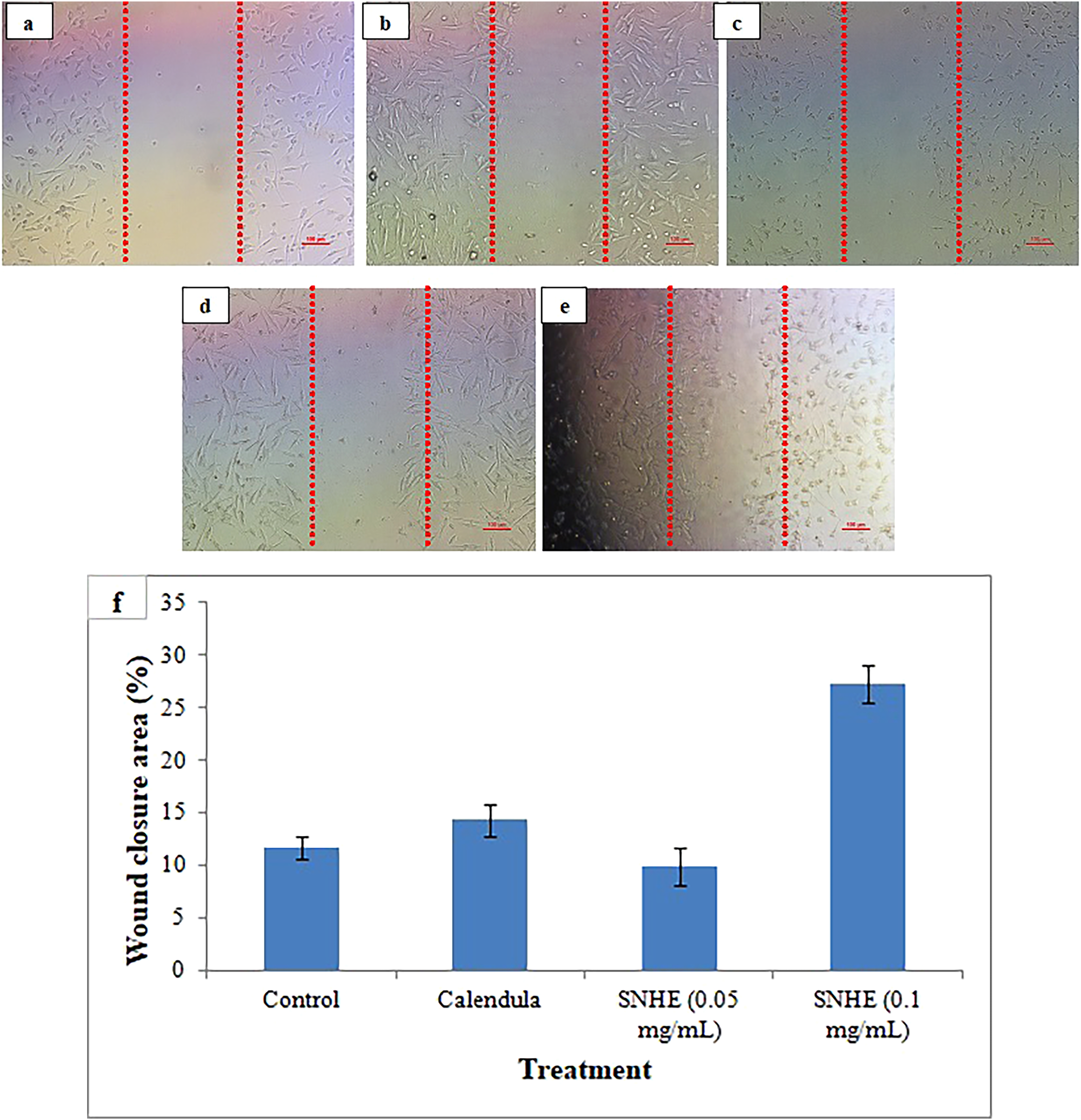
Effect on migration of McCoy fibroblast: micrograph from scratch assay: (a) fibroblasts immediately after scratch creation, (b) untreated fibroblasts 24 h post scratch, (c) Calendula cream-treated fibroblasts 24 h post scratch, (d) 0.05 mg/ml of SNHE treated fibroblasts 24 h post scratch, (e) 0.1 mg/ml of SNHE treated fibroblasts 24 h post scratch, and (f) quantification of fibroblast that migrated toward the scratch area. The results were expressed as mean ± S.D. of independent replicates and error bars represent the standard deviation from the mean values. Significance was determined at the P < 0.05 level based on t-test. *SNHE- Solanum nigrum fruit hydro-alcoholic extract.
In vivo wound healing study of SNHE in Zebrafish and diabetic rats
The in vivo wound healing potential of SNHE was evaluated by using adult Zebrafish and diabetic rats. Adult Zebrafish are valuable models for studying wound healing because of their fully developed immune system and robust regenerative abilities, which facilitate the assessment of tissue regeneration potential following fin amputation. 55 Figure 7(a)–(d) shows images of the regeneration of the caudal fin following amputation and treatment with SNHE (2 and 4 mg) and Calendula cream (2 mg). In contrast to the control group, caudal fins that had been exposed to SNHE exhibited significant regeneration (p < 0.05). A remarkable 50.67% regeneration rate was attained by a fish group exposed to 4 mg of SNHE (Figure 7(e)). The untreated group had a regeneration rate of 35%, while the groups that received Calendula cream and 2 mg of SNHE had regeneration rates of 45% and 46.67%, respectively. Similarly, in diabetic rats, SNHE-treated group showed increased wound contraction percentages with significant differences (p < 0.05) (Figure 8(a)–(d)). On the 14th day, SNHE application resulted in a wound closure area of 85.54%, compared to 71.52% for the water-treated group and 76.85% for the calendula-treated group. These results demonstrate that SNHE administration improves tissue regeneration, increases epithelization, and accelerates up wound closure. The potential of SNHE to cure wounds is consistent with findings from Owusu et al. (2024), who reported that an ethanolic leaf extract from S. mauriatianum promoted complete wound closure (100 ± 0.00%) by day 18, outperforming standard sulphadiazine treatment. 56 Similarly, in excision wound models, Zafar et al. (2024) demonstrated that methanolic extracts of Periploca aphylla and S. nigrum significantly improved wound healing, surpassing framycetin ointment. 57 These findings highlight the therapeutic potential of Solanum species extracts, particularly SNHE, as promising candidates for wound healing. Increased fibroblast activity, stimulation of angiogenesis, accelerated regeneration of epithelial tissue, and inhibition of inflammatory responses are likely the factors responsible for the wound-healing effects.

vComparative evaluation of fin regeneration on the 7th day post transection: (a) untreated Zebra fish, (b) Zebra fish exposed to Calendula cream, (c) Zebra fish exposed to 0.05 mg/ml of SNHE, (d) Zebra fish exposed to 0.1 mg/ml of SNHE, and (e) graphical representation illustrating the effects of various treatments on fin regeneration of Zebra fish. The results were expressed as mean ± S.D. of independent replicates and error bars represent the standard deviation from the mean values. Significance was determined at the P < 0.05 level based on t-test. *SNHE- Solanum nigrum fruit hydro-alcoholic extract.

Comparative evaluation of wound healing in diabetic Wistar Rats on 14th day: (a) water treated group, (b) Calendula cream treated group, (c) SNHE treated group, (d) graphical representation illustrating the effects of various treatments on wound healing in diabetic rats. The results were expressed as mean ± S.D. of independent replicates and error bars represent the standard deviation from the mean values. Significance was determined at the P < 0.05 level based on t-test. *SNHE- Solanum nigrum fruit hydro-alcoholic extract.
Screening of active chemical compounds and druggable targets in S. nigrum
A total of 117 phytochemicals of S. nigrum were retrieved from databases, including IMPAT and KNApSAcK. Consequently, four active molecules for S. nigrum were identified, such as Beta-Ionone, Ascorbic acid, Citric acid, and Nicotinamide (Figure 9, Supplementary 1). These compounds belong to different chemical classes, including terpenoids, vitamins, and organic acids, which highlight S. nigrum's phytochemical diversity. Each compound has reported bioactivities associated with the diabetic wound healing: beta-ionone possesses antibacterial, anti-inflammatory, and antioxidant properties 58 ; ascorbic acid is crucial for reducing oxidative stress, collagen synthesis, tissue regeneration, and keratinocyte differentiation 59 ; citric acid has antimicrobial effects 60 ; and nicotinamide has antibacterial, anti-inflammatory, and antioxidant qualities. 61

Active compounds present in S. nigrum (a) Beta-Ionone, (b) Ascorbic acid, (c) Citric acid, and (d) Nicotinamide.
The Swiss target prediction database and the STITCH database were subsequently employed to identify a comprehensive set of 249 targets corresponding to the aforementioned active compounds. The diversity of these targets, which include enzymes, receptors, and signaling molecules, supports a multitarget pharmacological profile. This is consistent with the broad therapeutic effects of S. nigrum in traditional medicine, particularly in the treatment of diabetic foot ulcers.
Potential targets of S. nigrum against DFU
3095 targets were identified from GeneCards and OMIM databases which were found to be associated with DFU. Further, 117 targets were observed to be common between the 3095 disease targets and 249 drug targets (Figure 10(a)). To elucidate the intricate relationships between these targets, a network was constructed and visualised using Cytoscape, displaying the target gene network and active compounds of S. nigrum (Figure 10(b)). Notably, with 110 targets each, beta-ionone and nicotinamide exhibited the highest degree of network connectivity, followed by citric acid and ascorbic acid, each with 56 targets.
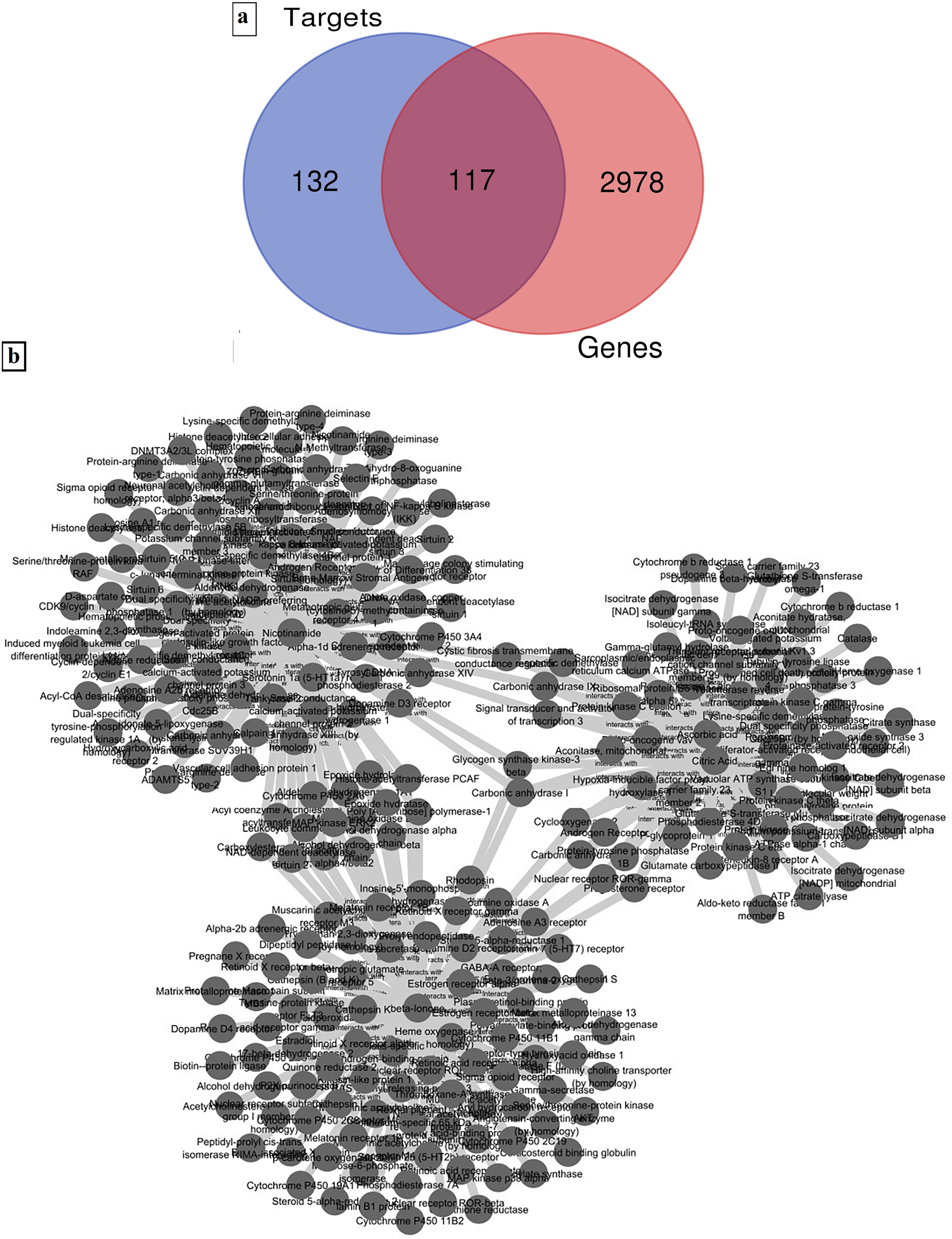
(A) Venn diagram of targets of S. nigrum active compounds and DFU genes, (b) Active compound-target genes network of S. nigrum.
High network degree values indicate that beta-ionone and nicotinamide may act as essential bioactive components, exerting wide pharmacological effects through multiple molecular interactions. In DFU treatment, where several pathological factors—oxidative stress, chronic inflammation, poor angiogenesis, and microbial infection—occur concurrently, the ability to act on numerous targets is particularly advantageous. The notable target coverage of citric acid and ascorbic acid suggests they may support and enhance the primary wound-healing actions of beta-ionone and nicotinamide.
The PPI map of the common targets of S. nigrum and DFU
The 117 drug targets mentioned earlier underwent further analysis through the Protein-Protein Interaction (PPI) network on the STRING platform. Following this, the data was imported into Cytoscape version 3.10.1 to create the PPI map, comprising 844 edges and 117 nodes (Figure 11). Our findings revealed an average node value of 14.4 for the target proteins. The existence of hubs—that is, nodes with relatively high degrees that typically have significant involvement in the cellular activities inside a network—was then investigated. Hub nodes included AKT1, STAT3, PPARG, PTGS2, ESR1, SIRT1, PARP1, HMOX1, and CYP3A4 among S. nigrum's DFU-associated targets (Supplementary 2).

Protein–protein interaction network of the 117 common target genes of S. nigrum and DFU.
GO and KEGG enrichment analysis
Hub gene targets were analyzed and enriched through the DAVID and STRING databases. These targets included biological process (BP), molecular function (MF), and cell composition (CC). We observed that S. nigrum could interact with 76 biological processes, 23 molecular functions, and 11 cell components associated to DFU. In Figure 12(a), 12(b), and 12(c), a bubble chart displays the ten most enriched terms in BP, MF, and CC. Among them, some biological processes were closely related to diabetic wound healing, including transforming growth factor beta receptor signaling pathway, response to oxidative stress, angiogenesis, positive regulation of nitric oxide biosynthetic process, positive regulation of endothelial cell proliferation, cellular response to insulin stimulus, glucose homeostasis, positive regulation of angiogenesis, regulation of cell proliferation, inflammatory response, positive regulation of cell migration involved in sprouting angiogenesis, cellular response to nerve growth factor stimulus, positive regulation of fibroblast proliferation, and regulation of inflammatory response. These mechanisms are consistent with our experimental findings, in which S. nigrum stimulated fibroblast proliferation, migration, and antioxidant activity, implying a molecular foundation for the reported wound-healing potential.
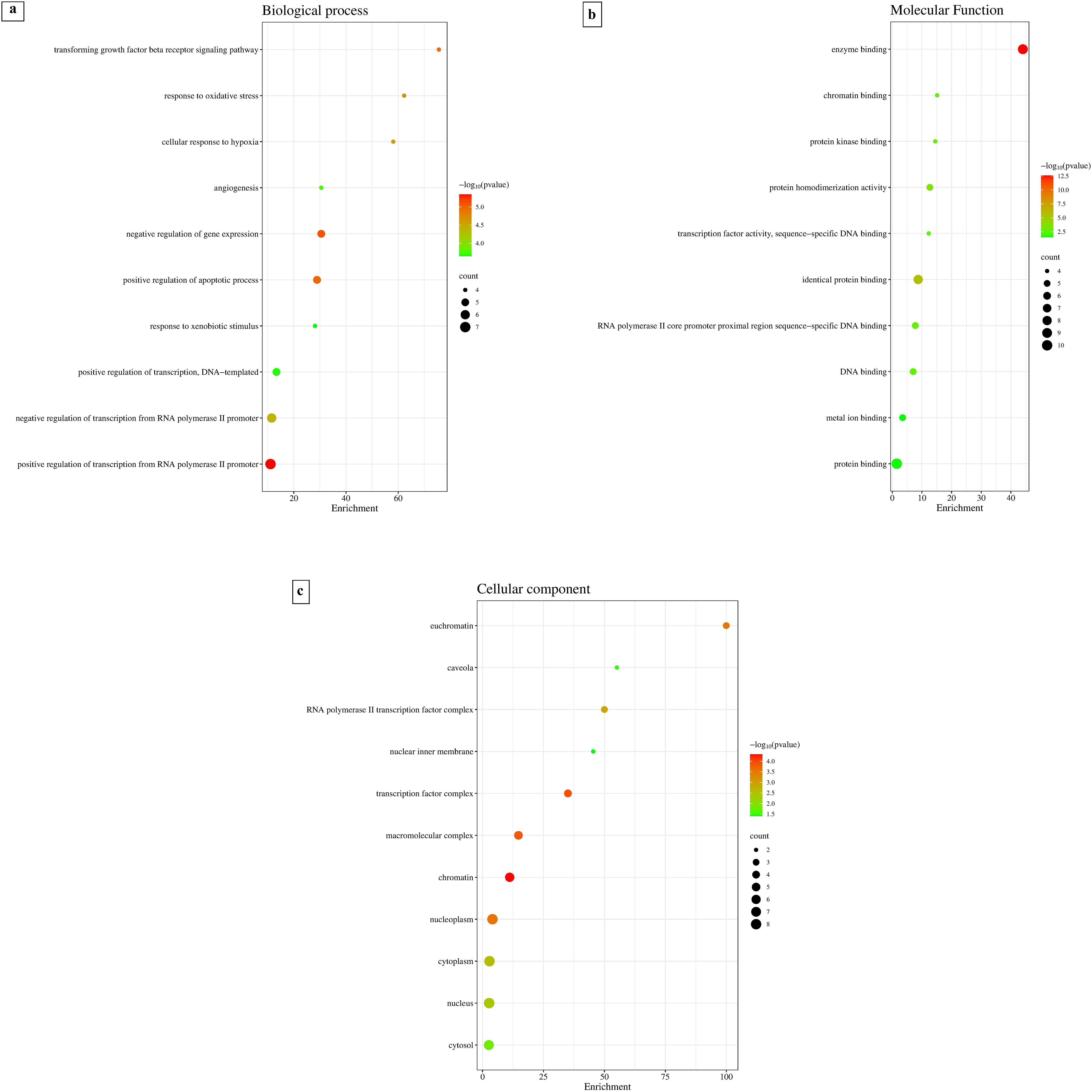
Go enrichment analysis of intersection targets of S. nigrum (a) Biological processes, (b) Molecular functions, and (c) Cell components.
46 pathways in all were enriched as a result of data analysis. Table 3 shows that 27 channels were selected by setting the parameter, p ≤ 0.05.
KEGG enrichment analysis of targets.

The development, metastasis, and drug resistance of DFUs are closely correlated with certain signalling pathways, including Pathways in Cancer, Chemical Carcinogenesis - Receptor Activation, Kaposi Sarcoma-Associated Herpesvirus Infection, Lipid and Atherosclerosis, and Prolactin Signalling Pathway, to varying degrees. The lipid and atherosclerosis pathways reflect the vascular complications associated with DFU pathogenesis, whereas the cancer-related pathways highlight the risk of malignant transformation in chronic, non-healing ulcers. DFUs are typically caused by three factors: arterial occlusive disease, trauma with secondary infection, and diabetic neuropathy. The primary cause of DFU is atherosclerosis driven on by peripheral vascular disease, particularly in relation to the femoral-poplitea blood vessels and the tiny blood vessels below the knee. Patients with diabetes have a twice-the-risk of developing ulcers in this condition as compared to those without diabetes. 62 DFU have the potential to turn cancerous. They set up a favorable environment for the growth of cancer. Additionally, a number of characteristics like enlargement, verrucous everted edges, and contact bleeding raise the possibility that a persistent wound represents malignant development. 63
Molecular docking of the screened components of S. nigrum and top four hub genes
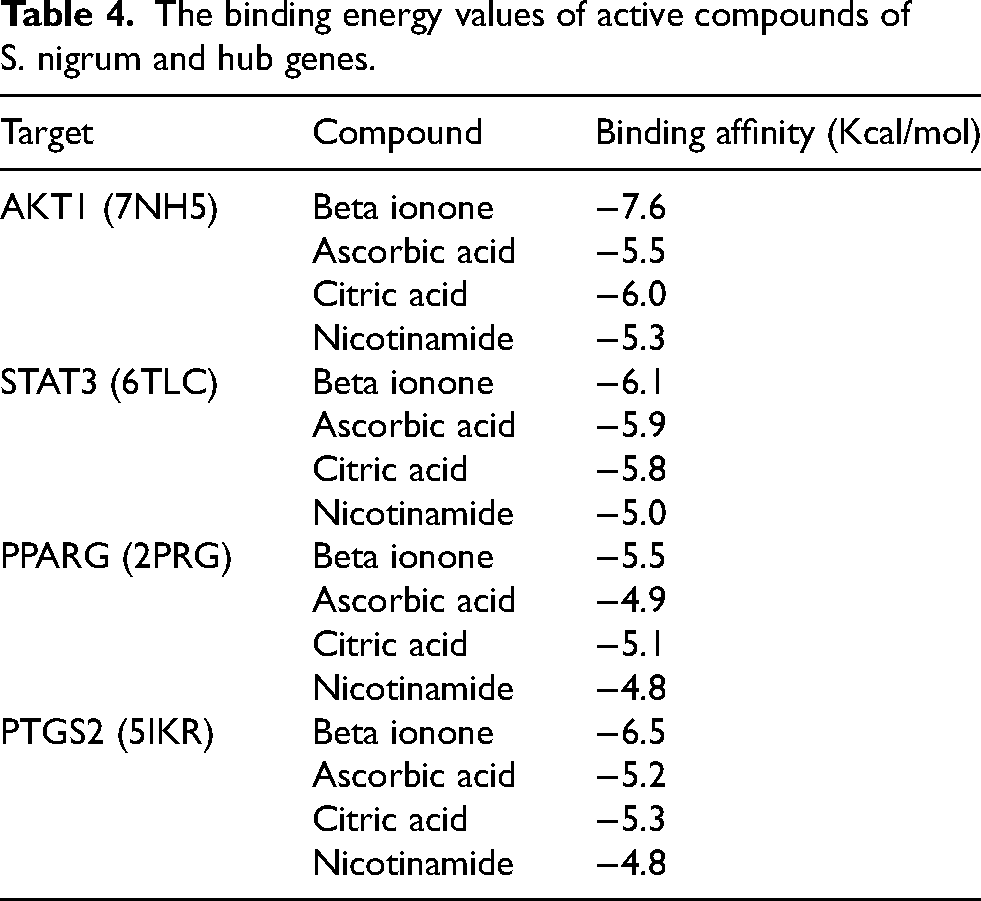
Molecular docking in conjunction with network pharmacology can yield strong data regarding the molecular mechanisms behind herbal medicine. To perform molecular docking, 4 active compounds in S. nigrum and top 4 hub gene targets of DFU- AKT1 (PDBID: 7NH5), STAT3 (PDB ID: 6TLC), PPARG (PDBID: 2PRG), and PTGS2 (PDBID: 5IKR) were selected. Every compound-target pair has a docking score of less than −4.8 kcal/mol, suggesting that all four active compounds—nicotinamide, ascorbic acid, citric acid, and beta-Ionone—have strong binding affinities to their respective targets (Table 4). Such binding energies indicate stable ligand-target interactions, implying that these drugs may possess biological effects associated with management of DFU control through these targets. In conclusion, our analysis of the PPI network revealed critical hub nodes linked with S. nigrum's DFU-related targets, such as AKT1, STAT3, PPARG, and PTGS2. These hubs have a remarkable significance when considering the pathogenesis of diabetic foot ulcers (DFUs).
The binding energy values of active compounds of S. nigrum and hub genes.
AKT1 is a signaling pathway junction protein that affects a variety of biological processes, including angiogenesis, growth, survival, proliferation, and cell metabolism, by serine or threonine phosphorylating a range of downstream substrates. 64 STAT3 signalling is a crucial intrinsic mechanism for cell proliferation since it is frequently activated in wounded tissues. Numerous genes that are regulated by STAT3 encode growth factors and cytokines, and their receptors then trigger the same STAT3 pathways, controlling the proliferation of cells. 65 PPARG is essential for transcriptionally coordinating the transition of pro-inflammatory macrophages to anti-inflammatory macrophages, which is necessary for reducing inflammation and resolving homeostasis. It also stimulates the proliferation of fibroblasts. 66 One mechanism linked to the pathophysiology of DFU is the glucose-mediated modification of the cyclooxygenase pathway activity, which leads to decreased prostaglandin synthesis and function. PTGS2, an isoform of inducible cyclooxygenase, is linked to the activation of subsequent inflammatory reactions. 67 Targeting these essential hub proteins is crucial, as the thorough understanding of these molecular pathways illuminates possible therapeutic paths for treating DFU, further investigation is warranted.
Conclusion
SNHE exhibits strong potential as a topical treatment for DFUs owing to its anti-inflammatory, antioxidant, hypoglycemic, and antibacterial properties. It promotes fibroblast proliferation and expedite wound closure in cellular investigations, and accelerate wound contraction in animal models. Our study demonstrates a systematic investigation of S. nigrum using in vitro, in vivo, and network pharmacology analyses, identifying four bioactive compounds (beta-ionone, ascorbic acid, citric acid, nicotinamide) that target AKT1, STAT3, PPARG, and PTGS2, offering mechanistic insight. Hence, S. nigrum has the potential for systemic as well as topical strategies and can be used as an adjunct therapy for chronic non healing wound conditions associated with diabetes mellitus.
Future scope
Further investigations could focus on development and comprehensive evaluation of S. nigrum topical formulations, along with molecular mechanism studies to better understand the action of its bioactive constituents. Furthermore, exploring synergistic effects with other conventional or natural therapies could broaden its potential for managing diabetic wounds.
Abbreviations
2-NBDG, 2-(N-(7-Nitrobenz-2-oxa-1,3-diazol-4-yl)Amino)-2-Deoxyglucose; ABTS, 2–2′-azino-di-(3-ethylbenzthiazoline sulfonate); DFU, Diabetic foot ulcer; DM, Diabetes mellitus; DMEM, Dulbecco's modified eagle medium; DNSA, 3,5-dinitrosalicylic acid; DPPH, 2
Supplemental Material
sj-docx-1-mgc-10.1177_10241221251380070 - Supplemental material for Topical application of Solanum nigrum ameliorate impaired wound healing associated with diabetes mellitus: A network pharmacology and experimental approach
Supplemental material, sj-docx-1-mgc-10.1177_10241221251380070 for Topical application of Solanum nigrum ameliorate impaired wound healing associated with diabetes mellitus: A network pharmacology and experimental approach by Diksha Manhas and Udayabanu Malairaman in Main Group Chemistry
Footnotes
Acknowledgements
We would like to express our gratitude to Jaypee University of Information Technology, Waknaghat, H.P., India for providing research facilities for this work. Further, the authors thank Dr Ritesh Jain, Chouksey Engineering College for the animal studies.
Ethics statement
All experiments were carried out in accordance with CPCSEA guidelines and Institutional Animal Ethical Committee, Chouksey Engineering College, Bilaspur (Ref. SOP/IAEC/2023/March/11), and Institutional Ethics Committee, Jaypee University of Information Technology (Ref. IEC/Project No-02/2023).
Author contribution
Udayabanu Malairaman: Conceptualization, Writing- review and editing, Validation, Visualization, Supervision, Formal analysis. Diksha Manhas: Conceptualization, Methodology, Investigation, Formal analysis, Writing- review and editing, Writing- original draft, Software, Visualization.
Funding
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Declaration of interests
The authors declare that this manuscript has not been published or currently under review for publication elsewhere. No potential conflict of interest was reported by the authors.
Data availability
All data generated or analyzed during this study are included in this published article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.