
Abstract
During June and July 2012, Buffalo Pound Lake and Blackstrap Lake in Saskatchewan, Canada were visited biweekly and surveyed for sick and dying fish. During this investigation, 2 fish kills were identified. Buffalo Pound experienced a large die-off of yellow perch (Perca flavascens) in June, while Blackstrap experienced a die-off of lake whitefish (Coregonus clupeaformis) in July. In excess of 50 fish were examined for gross lesions at each lake, and dead and moribund fish consistently had 1 or more of the following lesions: multifocal petechial cutaneous hemorrhage, skin ulceration, or branchial necrosis. Of these, 17 fish were collected for necropsy, and major tissues were submitted for histology. Aerobic bacterial culture was performed on 16 out of 17 fish. In 7 out of 8 (88%) yellow perch, the body wall had multiple areas of pale discoloration that corresponded to erosion and ulceration of the skin. Seven out of 8 (88%) whitefish had severe necrotizing branchiitis, and 8 out of 8 (100%) had severe epicardial parasitism, consistent with Ichthyocotylurus erraticus. Wet mounts of skin and gill lesions demonstrated filamentous bacteria with gliding motility, which often formed haystack-like arrangements. Flavobacterium columnare and Aeromonas hydrophila were cultured from skin and gill lesions of all fish. Based on the characteristic appearance and distribution of lesions, mortality was attributed to columnaris disease with secondary infection with A. hydrophila. The current study demonstrates that columnaris disease is an important contributor to fish kills in southern Saskatchewan lakes. However, further research is needed to determine what role environmental factors play in outbreaks of columnaris disease in prairie lakes.
In July 2011, thousands of dead fish, mainly small yellow perch (Perca flavascens) and few sticklebacks (family Gasterosteidae) were found in Buffalo Pound Lake, and specimens were subsequently submitted frozen to the Canadian Wildlife Health Cooperative (Saskatoon, Saskatchewan, Canada). Complete necropsies revealed large numbers of myxosporidial spores throughout the spinal cord and brain of yellow perch, consistent with Myxobolus neurophilus. Based on the field observations of abnormally swimming fish, and the location and severity of lesions, it was hypothesized that mortality may have been due to parasitic infection; however, M. neurophilus was subsequently identified in clinically normal perch from additional lakes in Saskatchewan. 16 The findings indicate that other factors may have contributed to the die-off in Buffalo Pound Lake. The objective of the current study was to monitor 2 southern Saskatchewan lakes for fish kills and identify infectious pathogens that contribute to these events in prairie lakes.
Buffalo Pound Lake (50°39′N 105°30′W) and Blackstrap Lake (51°47′N 106°25′W) are man-made reservoirs, which are part of the Qu’Appelle drainage system in southern Saskatchewan. Both lakes are shallow (depth 5–9 m), exhibit little to no thermal stratification, are eutrophic, and have experienced fish kills in the past. In June and July 2012, these lakes were monitored for fish kills once every 2 weeks during daylight hours, or once per week when die-offs were occurring. The length of each reservoir was explored by boat in order to survey for dead and dying fish.
Dead and sick fish were examined for lesions, and a subset of fish was collected by dip net and subsequently euthanized in a water bath supersaturated with tricaine methanesulfonate (MS-222) a buffered with sodium hydroxide. Complete necropsies were conducted on site, and representative sections of all major tissues were fixed in 10% neutral buffered formalin, decalcified with 5% formic acid, trimmed into 5-µm-thick sections, and stained with hematoxylin and eosin for routine histologic examination. A subset of slides was additionally stained with Brown and Brenn (Gram stain). Unstained wet mounts of external lesions were examined under light microscope.
Fresh samples of gill and/or skin, kidney, spleen, and less frequently heart and liver, were aseptically cultured onto blood agar, MacConkey agar, and Cytophaga agar supplemented with neomycin b (5 μg/ml) and polymyxin B c (10 IU/ml). Plates were incubated at room temperature (22–25°C) for 48 hr and then subcultured until pure bacterial colonies were isolated. Bacteria were tested for the production of catalase and cytochrome oxidase by using a commercial kit d and 3% hydrogen peroxide, respectively. A commercial rapid identification system e was used to further characterize the biochemical phenotype of isolated bacteria.
Colonies that were morphologically consistent with Flavobacterium columnare were additionally tested for flexirubin-like pigment and Congo red absorption by flooding plates with 20% potassium hydroxide c and Congo red indicator, f respectively. Casein degradation was determined by streaking F. columnare–suspect colonies on milk agar plates.
Genomic DNA was isolated from 2 isolates from each lake, which were morphologically and biochemically compatible with F. columnare, using a commercial kit g following the manufacturer suggested protocol for animal tissue. Amplification of a segment of the 16S ribosomal RNA (rRNA) gene known to be specific for F. columnare was performed using the primers FvpF1 (5′-GCCCAGAGAAATTTGGAT-3′) and FvpR1 (5′-TGCGATTACTAGCGAATCC-3′) from a published study. 1 A known F. columnare strain, confirmed by sequencing of the 16S rRNA PCR product from the species-specific PCR 1 and a no-template control were used as positive and negative controls, respectively.
Based on field observations and reports made by members of the public, Buffalo Pound Lake experienced a large fish die-off during June 8–19. Yellow perch were primarily affected, but small numbers of fathead minnows (Pimephales promelas), three-spined sticklebacks (Gasterosteus aculeatus), and white suckers (Catostomus commersonii) were also found dead. Dead fish were scattered on the shoreline, many of which were autolyzed and covered by white fungal plaques (consistent with Saprolegnia sp.). Several sick yellow perch were found floating or weakly swimming in dorsal recumbency.
In excess of 50 fish were examined for external lesions, all of which consistently had multifocal cutaneous petechial hemorrhages and erosion and ulceration of the skin. Of these, 8 moribund yellow perch (mean fork length 13.7 cm) and 1 male fathead minnow (fork length 3 cm) were collected for necropsy and bacterial culture. Of the yellow perch collected, 2 were female, 1 was male, and 5 were undetermined. Seven out of 8 (88%) yellow perch had areas of pale discoloration on the body wall (Fig. 1), corresponding to loss of surface scales, erosion of the skin, degeneration of the underlying musculature, and infiltration of Gram-negative filamentous bacteria. One yellow perch had mild necrotizing branchiitis (Fig. 2). In 50% (4/8) of yellow perch, there were large parasitic pseudocysts and free myxospores distributed throughout the spinal cord and brain, consistent with M. neurophilus. The internal viscera were grossly normal.

Yellow perch (Perca flavascens). There are locally extensive areas of skin discoloration, which correspond to areas of erosion and ulceration.

Body wall; yellow perch (Perca flavascens). There is a loss of surface scales, and the epidermis, dermis, and underlying musculature is diffusely necrotic. Hematoxylin and eosin. Bar = 200 µm.
Based on field observations and reports made by the public, Blackstrap Lake experienced a large die-off of lake whitefish (Coregonus clupeaformis) between July 24 and August 4. Dead fish were found in all areas of the lake, and many were water-logged and autolyzed. Moribund fish were floating at the water surface with flared opercula and, in most instances, were nonresponsive when handled. There was a small amount of green algae near the shoreline, and although water temperature was not recorded, it was subjectively warmer compared to earlier in the summer.
In excess of 50 fish from Blackstrap Lake were examined for external lesions, all of which were consistently covered by large amounts of mucous and had multifocal cutaneous hemorrhages; severe gill necrosis was present in the majority of fish examined. Of these, 7 moribund and 1 dead white fish were collected for necropsy and bacterial culture; 6 males and 2 females, with an average weight of 2.15 kg and fork length of 48 cm. All fish were covered by an excessive amount of mucous, and the base of the fins was erythematous. In 88% (7/8) of fish collected, up to 25% of the gill filaments were eroded and replaced by brown necrotic debris, hemorrhage, and stacked aggregates of filamentous Gram-negative bacteria, small Gram-negative rods, and few algae (Figs. 3, 4); motile filamentous bacteria were noted on wet mount preparations. In 100% (8/8) of fish, the epicardium was diffusely covered by 1-mm-diameter white nodules, which corresponded to metacercariae surrounded by granulomatous inflammation. In 100% (8/8) of whitefish, there was marked lymphoid depletion in the spleen, and melanomacrophage centers were abundant.

Gills; lake whitefish (Coregonus clupeaformis). The trailing edge of gill filaments are eroded and replaced by brown necrotic debris.
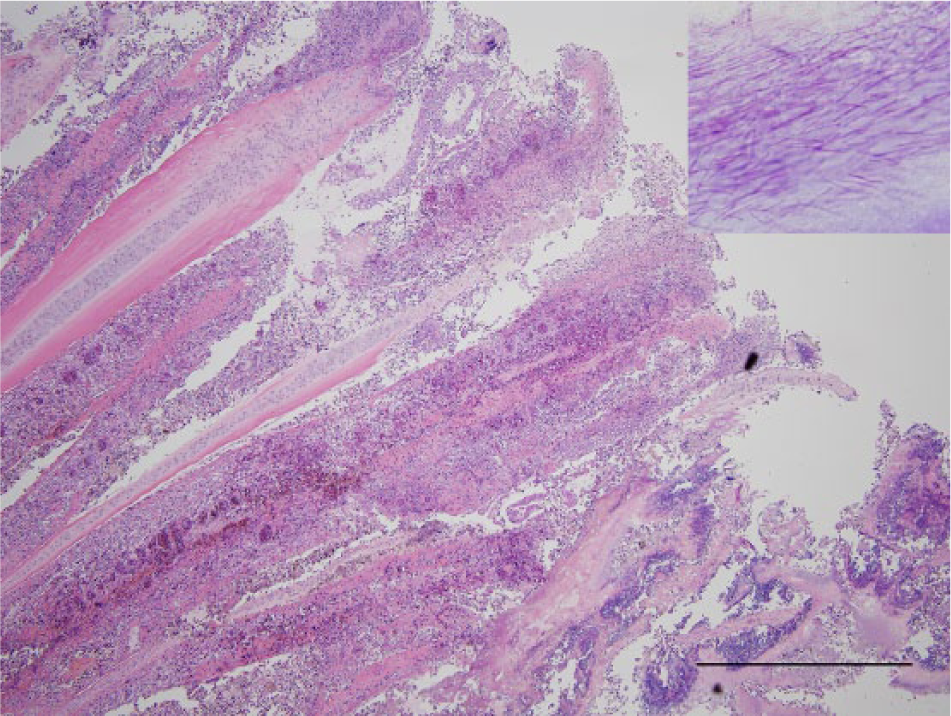
Gills; lake whitefish (Coregonus clupeaformis). The normal gill architecture is replaced by necrotic debris. There is hemorrhage and severe congestion of the primary gill filaments. Hematoxylin and eosin. Bar = 1 mm. Inset: large numbers of filamentous Gram-negative bacteria form haystack-like arrangements, consistent with Flavobacterium columnare. Brown and Brenn stain.
The commercial rapid identification system identified Aeromonas hydrophila and a “Non-fermenter” spp. in all fish in which culture was performed. The isolates identified as Non-fermenter spp. formed yellow rhizoid colonies and had phenotypic properties that were in agreement with F. columnare in previous studies in North America and Europe, with the exception that sodium thiosulfate was not produced.8,11 The species-specific PCR identified these Non-fermenter spp. isolates as F. columnare. The characteristic features of F. columnare, which are noted in the current report, include production of flexirubin-like pigment, Congo red absorption, digestion of gelatin, reduction of nitrates, and formation of strongly adherent, yellow rhizoid colonies. The discrepancy in sodium thiosulfate in the present study compared to others may be explained by differences in the methodology used in other studies, or possibly due to differences in F. columnare strains. For example, the biochemical profile of F. columnare strains identified in South America differs from that described in North American and European fish.4,6,14,17
Flavobacterium columnare and A. hydrophila were cultured from 100% and 86% of body wall lesions in yellow perch, respectively, and both were cultured from the gills in 100% of lake whitefish. In 100% of fish, 1 or a combination of F. columnare and A. hydrophila was isolated from 1 or more internal organs. Shewanella putrefaciens was cultured from the kidney in 1 dead whitefish that was moderately autolyzed.
The species selectivity and the duration of the fish kills observed in the Buffalo Pound and Blackstrap fish kills was suggestive of environmental perturbation with subsequent opportunistic infection. 12 Identifying bacterial disease in wild fish is often complicated by rapid autolysis and overgrowth by saprophytic bacteria. However, these issues were circumvented in the present study by actively surveying lakes and collecting live fish, which exhibited lesions similar to those present on dead fish. 8 Based on postmortem and histologic findings, in combination with bacteriologic results, both fish kills were attributed to an outbreak of F. columnare that was likely secondary to suboptimal environmental conditions.
Flavobacterium columnare, the etiologic agent of columnaris disease, is ubiquitous in freshwater habitats, and has been reported as part of the normal bacterial microflora of fish. 3 Opportunistic infection arises when fish are stressed due to suboptimal water conditions, including: increasing water temperatures, high levels of nitrate and/or organic matter, low levels of dissolved oxygen, and high stocking density.2,3,18,19 Clinical infection is associated with high morbidity and mortality, and is manifested as necrosis of the gills and erosion and ulceration of the skin. Skin lesions typically originate at the base of the dorsal fin and extend ventrally on either side of the body forming a circumferential pale band, creating the classic “saddle back” lesion that is often associated with columnaris disease. 3
In many cases, columnaris disease is complicated by secondary bacterial and fungal infections, which can likely be attributed to a combination of immunosuppression and damage to the natural barrier provided by the skin. 4 A mixed infection of F. columnare and A. hydrophila was the ultimate cause of death in several thousand common carp (Cyprinus carpio carpio) in the St. Lawrence River, 13 which is similar to what was found in the Buffalo Pound and Blackstrap die-offs reported in the current study. Flavobacterium columnare was considered the primary infectious agent in the present study due to the characteristic appearance and distribution of lesions.
The difference in fish species affected in the Buffalo Pound and Blackstrap die-offs likely reflect differences in underlying factors that predisposed fish to columnaris disease. In Buffalo Pound, oxygen levels did not appear to be depleted because large fish were not affected, ambient air temperature was cool, and there appeared to be adequate surface wind mixing based on field observations. A fishery stock assessment, which was conducted at the same time as the fish kill, revealed that the yellow perch population in Buffalo Pound was extremely abundant based on catch per unit effort (Jennifer Merkowski, personal communication, 2012). This is of particular interest because not only does high stocking density enhance horizontal transmission of bacteria, but also it acts as a stressor, predisposing yellow perch to columnaris disease. 15 An outbreak of columnaris disease occurred previously in white suckers in Buffalo Pound Lake in June 1972; however, factors that may have triggered the outbreak were not discussed. 21
Based on field observations and the fact that large fish were targeted during the Blackstrap fish kill, the whitefish mortality was probably due in part to increasing water temperatures and reduced oxygen availability. The epicardial metacercaria in whitefish from Blackstrap is consistent with Ichthyocotylurus erraticus based on the location of parasites and host species. 9 The severity of this lesion may interfere with cardiovascular function and the delivery of oxygenated blood in whitefish. 5 A similar parasite, Apatemon gracilis, is reported to cause restrictive pericarditis in rainbow trout (Oncorhynchus mykiss), and reduce stroke volume by as much as 50%. 20 A combination of environmental stress and chronic parasitism may have accounted for the splenic lymphoid depletion seen in the current study.
Although cyanobacteria are known to reside in nutrient-rich lakes, microcystin toxicity was considered unlikely in the current study because algal blooms were not observed, and fish did not have characteristic lesions. 7 Most fish kills caused by agricultural and industrial toxins involve several different species of fish and occur within a short time frame, 12 neither of which were features of die-offs in the present study. Toxic levels of hydrogen sulfide and ammonia were also considered an unlikely cause of mortality due to an absence of typical lesions. 10 In conclusion, the current study identified F. columnare as an important contributor to fish kills in southern Saskatchewan lakes.
Footnotes
Acknowledgements
The authors would like to thank Saskatchewan Ministry of Environment staff for their assistance in collecting yellow perch in summer 2011. The authors would also like to thank the Animal Health Centre, Plant and Animal Health Branch, BC Ministry of Agriculture, Abbotsford, British Columbia, for performing the polymerase chain reaction assays, and Dr. Frederick A. Leighton for reviewing earlier versions of this article.
a.
Syndel Laboratories Ltd., Qualicum Beach, British Columbia, Canada.
b.
EMD Bioscience Inc., La Jolla, CA.
c.
Sigma-Aldrich, St. Louis, MO.
d.
DrySlide oxidase slides, BD Diagnostic Systems, Sparks, MD.
e.
API20E rapid identification system, bioMérieux Canada Inc., St. Laurent, Quebec, Canada.
f.
Ricca Chemical Co., Arlington, TX.
g.
DNeasy blood & tissue, Qiagen GmbH, Hilden, Germany.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Funding for the study was provided by the Saskatchewan Fish and Wildlife Development Fund.