
Abstract
Dengue is the most important mosquito-borne viral infection of humans worldwide. Genetic ancestry of the host is a significant risk factor for severe dengue, but the mechanisms are not known. Recent findings using human skin explants from genetically defined donors reveal that dengue virus (DENV) replication and spread in skin increases with increasing proportion of European ancestry of the donor, associated with a strong inflammatory response and local myeloid cell infiltration, infection, and migration. In contrast, African ancestry is associated with substantially reduced cutaneous inflammation and cell infiltration following virus inoculation, resulting in reduced infection and migration of infected cells. These findings help explain the long-standing observation that individuals of African descent are relatively protected against severe dengue, while individuals of European descent are not. In this essay, we review DENV infection, focusing on human skin and the influence of genetic ancestry on the cutaneous innate response and virus spread.
Introduction
Dengue is the most prevalent mosquito-borne viral infection of humans worldwide (Bhatt et al., 2013). The disease has rapidly expanded in incidence around the globe in the past three decades, associated with climate warming and increased population mobility (Zhang et al., 2025). In 2024, the number of reported dengue cases in the Americas alone approached 13 million (Pan American Health Organization, 2024). Infection with any one of the four serotypes of dengue virus (DENV) is usually asymptomatic, while in about 25% of infections, patients present with an acute self-limited febrile illness known as dengue (prior to World Health Organization reclassification of dengue cases in 2009, this was called dengue fever) (World Health Organization, 2009). A small proportion of cases rapidly progress to severe dengue (traditionally known as dengue hemorrhagic fever and/or dengue shock syndrome), characterized by systemic vascular leakage syndrome that can be fatal (Wilder-Smith et al., 2019). The factors that contribute to the increased risk of severe disease in individuals infected with DENV are many and varied. In this Bits essay, we will discuss recent work that sheds light on one of these factors: the genetic ancestry of the host.
Dengue Risk Factors
The most well-known risk factor for severe dengue is secondary infection. In this scenario, antibodies raised against the primary infecting serotype bind to but do not neutralize a heterologous serotype upon secondary infection. This leads to virus-antibody engagement with Fcγ receptors on myeloid cells, which then have enhanced uptake and replication of virus, a process termed antibody-dependent enhancement or ADE (Halstead and O’Rourke, 1977). Cases of secondary infection are most likely to develop into severe dengue disease when baseline preinfection antibody titers fall within a relatively narrow range (Katzelnick et al., 2017). Similarly, infants born to mothers with immunity to one DENV serotype may develop severe dengue upon infection with a heterologous serotype when maternal antibodies decline to subneutralizing concentrations (Chau et al., 2009; Kliks et al., 1989). Other well-described risk factors for severe dengue are advanced pregnancy and advancing age, certain underlying comorbidities (Wilder-Smith et al., 2019), and single nucleotide polymorphisms (SNPs) in a diversity of genes including innate immune mediators, Fcγ receptors, cytokines, and human leukocyte antigens (Alagarasu et al., 2013; Frota et al., 2021; Khor et al., 2011; Lin et al., 2009; Loke et al., 2002; Perez et al., 2010; Sakuntabhai et al., 2005; Vejbaesya et al., 2009).
Host Genetic Ancestry as a Dengue Risk Factor
Beginning more than four decades ago, epidemiological observations during dengue outbreaks in South and Central American and Caribbean countries with genetically admixed populations revealed that severe dengue disease was most likely to arise in individuals of European, rather than African, descent, identifying host ethnicity as a dengue risk factor (Blanton et al., 2008; Bravo et al., 1987; Chacon-Duque et al., 2014; Halstead et al., 2001; Sierra et al., 2007). In the 1981 dengue epidemic in Cuba, dengue hemorrhagic fever was more than 10 times more likely to be diagnosed in children of European versus African ancestry (Bravo et al., 1987). Population differences in disease severity were not due to differences in infection rates or hospitalization preferences (Guzman et al., 1990). In the mid-1990s in Haiti, dengue hemorrhagic fever was absent from Haitian children despite hyperendemic DENV transmission (Halstead et al., 2001). In contrast, dengue fever cases were relatively common among visiting U.S. military personnel assigned to Haiti during this period, with onset of illness beginning as soon as 7 days after deployment (Centers for Disease Control and Prevention, 1994). Polymorphisms in oxysterol binding protein-like 10 (OSBPL10) and retinoid x receptor alpha (RXRA) genes have been identified as conferring African-ancestry protection from severe dengue (Sierra et al., 2017). Additional studies with blood cells from individuals homozygous for either the European or African haplotypes of OSBPL10 indicate that the European haplotype enhances DENV infection associated with elevated expression of RXRA (Granela et al., 2024). Defining the role of genetic ancestry in dengue disease severity is complicated by the socioeconomic, cultural, and environmental factors that may affect different populations differently and thus impact the outcome of DENV infection (Blanton et al., 2008; Carabali et al., 2015). As such, biological processes that influence dengue disease as a function of genetic ancestry have been difficult to identify.
DENV Replication and Spread in Skin
The skin is the primary site of human DENV infection following the bite of an infected female Aedes mosquito, and the biology of virus infection and spread in the skin has been relatively well-studied. Virus is deposited within the epidermis and dermis, and virus replication begins in the region of basal keratinocytes, whether it is delivered by a bifurcated needle or mosquito bite (Castanha et al., 2024; Duangkhae et al., 2018; Limon-Flores et al., 2005). Productive virus replication occurs in Langerhans cells in the epidermis and fibroblasts, dendritic cells (DC), macrophages, endothelial cells, and mast cells in the dermis (Cerny et al., 2014; Duangkhae et al., 2018; Huang et al., 2000; Montes-Gomez et al., 2020; St John et al., 2011; Wu et al., 2000). A critical innate immune response to infection within skin that promotes DENV replication and spread is the elaboration of proinflammatory cytokines and chemokines by infected cells in both epidermis and dermis, as these result in local infiltration of additional Langerhans cells, DC, and macrophages to the site of virus inoculation that subsequently become infected (Fig. 1) (Cerny et al., 2014; Duangkhae et al., 2018; Surasombatpattana et al., 2011). Mosquito bites themselves induce edema and inflammation that stimulate local and systemic recruitment of myeloid cells to the site of infection (Castanha et al., 2024; Pingen et al., 2016). Infection leads to the induction of an antiviral response through the production of type I interferons (IFN) (Cerny et al., 2014; Duangkhae et al., 2018; Surasombatpattana et al., 2011). DENV inoculation results in an increase in migration out of skin of infected myeloid cells that disseminates virus (Fig. 1) (Duangkhae et al., 2018; Wu et al., 2000). Antibody-dependent enhancement of infection also occurs in human skin, as demonstrated by increased replication and spread of DENV-2 in skin in the presence of DENV-3 immune sera (Castanha et al., 2020).

Biology of DENV infection in human skin. Virus is deposited into the epidermis and dermis via the bite of an infected Aedes mosquito and replicates first in basal keratinocytes, followed by spread to Langerhans cells in the epidermis and DC, macrophages, and fibroblasts in the dermis. Infection induces an inflammatory response, including release of cytokines and chemokines that promote infiltration of resident Langerhans cells, DC, and macrophages, which in turn become infected, amplifying the response. Infected cells migrate from both epidermis (Langerhans cells) and dermis (DC and macrophages) through lymphatics to disseminate DENV throughout the host. Recent findings indicate that this biology is strongly influenced by host genetic ancestry. DC, dendritic cells.
Genetic Ancestry of the Host Influences Cutaneous Replication and Spread of DENV
Biological differences in DENV infection and cutaneous immunity in individuals of various ancestral backgrounds could well contribute to differences in dengue disease severity, as the magnitude of virus dissemination from skin likely has a direct impact on the extent and severity of resulting disease. To test this hypothesis, our group compared DENV infection and immunity in skin explants from a group of genetically defined healthy, dengue-naïve donors of African or European ancestry (Castanha et al., 2025). Genetic ancestry was quantified by analyzing each donor’s DNA using a panel of ancestry informative SNPs, allowing donors to be placed on a continuum of European ancestry from 0 to 1. As the proportion of European ancestry of donors increased and African ancestry decreased, there was a marked increase in DENV (all serotypes) and Zika virus infection and replication in both epidermis and dermis (Fig. 2A) (Castanha et al., 2025). Importantly, the inflammatory response and subsequent infiltration of resident Langerhans cells, DC, and macrophages from outside of the inoculated region into the site of primary virus replication increased with European ancestry and was essentially absent in African ancestry skin. Moreover, the migration of infected cells out of skin into media (and, by extension, the host) was seen only in DENV infection of skin explants from European donors. Cutaneous infection with DENV-2 was enhanced in the presence of pooled anti-DENV-3 immune serum, a model of secondary infection and ADE, to a significantly greater extent in European ancestry skin as compared with African ancestry skin (Fig. 2B) (Castanha et al., 2025). These new findings reveal that our basic understanding of DENV replication and spread in human skin, as depicted in Figure 1, is really the case only for individuals of European ancestry. Indeed, in reports that have defined DENV infection and spread using human skin explants or primary human skin cells, three used skin donors that were exclusively of European descent (Castanha et al., 2024; Castanha et al., 2020; Duangkhae et al., 2018), while others did not define donor ethnicity (Cerny et al., 2014; Limon-Flores et al., 2005; Surasombatpattana et al., 2011; Wu et al., 2000).
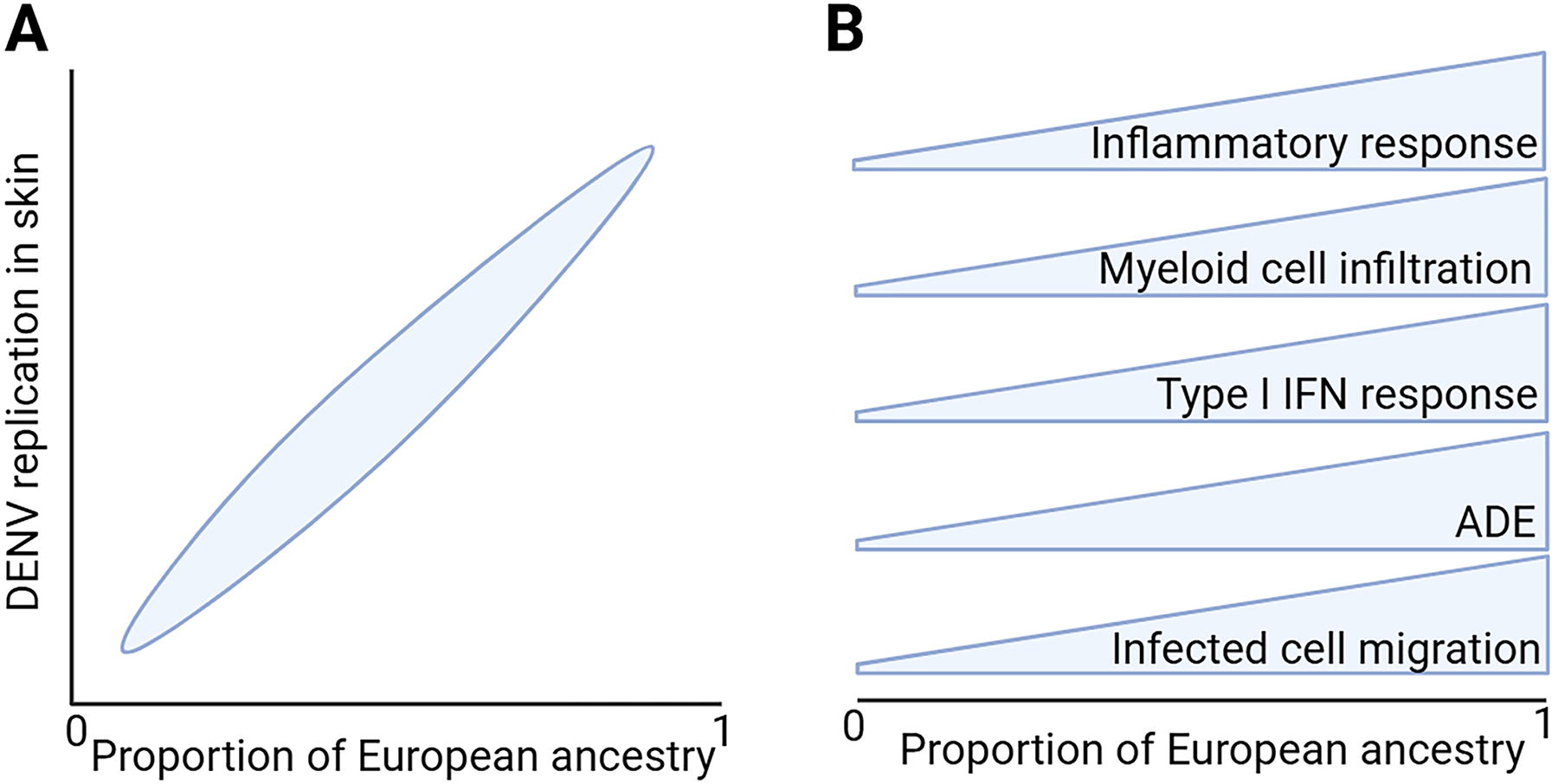
Schematic illustrating the effect of ancestry on DENV replication in human skin explants.
The Cutaneous IFN Response to DENV Infection
Viral infections induce type I IFN responses that initiate expression of a host of IFN-stimulated genes (ISGs), which then serve as effectors of the antiviral response, although several ISGs actually enhance replication of certain viruses (Schoggins et al., 2011). DENV infection of human skin cells also induces a type I IFN response (Cerny et al., 2014; Duangkhae et al., 2018; Surasombatpattana et al., 2011). As expected, in DENV infection in human skin explants, European ancestry was associated with a significant enrichment of genetic pathways associated with the type I IFN response, yet this enrichment failed to reduce the number of infected cells in skin (Fig. 2B) (Castanha et al., 2025). Polymorphisms in oligoadenylate synthetase (OAS) 1–3 and RXRA were present at greater frequencies in European donors, and were also associated with increased virus replication in skin, yet OAS3 expression in response to both type I IFN and DENV infection was substantially greater in European donors relative to African donors (Castanha et al., 2025). These data suggest that the type I IFN response, and the OAS response in particular, may promote rather than restrict DENV infection in skin, which could contribute to the increased virus replication seen in European relative to African ancestry skin. In human peripheral blood mononuclear cells, influenza virus infection also induces a stronger type I IFN response in European relative to African ancestry donors, yet, in contrast to skin, this response is associated with a subsequent reduction in virus titers (Randolph et al., 2021).
Outstanding Questions Concerning Genetic Ancestry and Dengue
Numerous questions remain to be answered before we can more completely understand the relationship between the genetic ancestry of the host and susceptibility to DENV infection and disease. First, with respect to mechanisms for enhanced infection and replication in European ancestry skin, what are the cellular and molecular factors driving this difference, and is the skin a unique tissue compartment when compared with blood in its innate response to viral infection across different human populations? Second, is there a real difference in the susceptibility of African versus European descendants to ADE, as noted in skin and suggested by observations in blood cells (Morier et al., 1987), and if so, what is the mechanism? Third, what are the molecular factors involved in the different susceptibilities of African and European descendants to dengue disease? For this to be answered, access to archived samples from individuals with well-characterized dengue disease who have also been characterized for their genetic ancestry will be needed. Several innate immune pathways and genes, including OSBPL10 and RXRA (Sierra et al., 2017), are candidates for systemic differences to dengue disease severity in different ancestral groups. Dysregulation of the alternative complement pathway has been correlated with dengue disease severity (Nascimento et al., 2009), and SNPs in genes encoding complement regulatory factors that predict either increased risk of infection or increased risk of inflammation have been identified (Harris et al., 2012). Interestingly, many of the SNPs in complement regulatory factors differ significantly in allele frequency across human populations (Ermini et al., 2012). Could ancestral differences in complement factors and complement function be contributing to population differences in dengue severity? Finally, ancestral differences in infection with and immunity to DENV could potentially impact the efficacy and adverse events associated with dengue vaccination. All dengue vaccines that are in advanced development are live-attenuated tetravalent vaccines that result in low-level virus replication in the recipient (Durbin, 2020; Saez-Llorens et al., 2025). Could vaccine-induced responses—both with respect to vaccine efficacy and complications—also differ across human populations? A detailed analysis of immune responses and adverse events, as well as innate immune and inflammatory markers in vaccinees that have been genetically typed for ancestry markers, could begin to address this important question.
Summary
We are now beginning to understand the biological basis for differences in dengue disease severity in different human populations that were first recognized in the early 1980s. Further research to define the ancestral influences on dengue and other arboviral diseases could lead to better patient management and vaccine utilization and may identify novel therapeutic targets for intervention.
Footnotes
Funding Information
No funding was received for this article.
Disclosure Statement
No competing financial interests exist.