
Abstract
Liver fibrosis (LF) is a fibrotic and inflammatory condition resulting from chronic liver damage, and the activation of hepatic stellate cells (HSCs) is the central event. Our prior research has revealed that the circAno6/miR-296-3p/Toll-like receptor (TLR4) pathway is a key signaling axis in the pathogenesis of LF. Shugan Jianpi Formula (SGJPF) is a traditional Chinese medicinal treatment widely used to treat LF. We hypothesized that SGJPF exerts anti-LF effects by modulating the circAno6/miR-296-3p/TLR4 signaling axis, attenuating inflammatory responses, and inhibiting HSC activation, thereby ultimately mitigating LF. In this study, an SGJPF extraction solution was first prepared and administered to rats via gavage at a specified dosage. Blood samples were subsequently collected from the abdominal aorta to prepare SGJPF-containing serum. Optimal SGJPF intervention conditions were determined using the Cell Counting Kit-8 assay. Flow cytometry was used to assess JS-1 cell cycle progression. The levels of interleukin-1 beta (IL-1β) and IL-18 were measured by enzyme-linked immunosorbent assay. The expression levels of collagen type I (collagen I), α-smooth muscle actin (α-SMA), circAno6, TLR4, miR-296-3p, and NOD-like receptor protein 3 (NLRP3) were evaluated using Western blotting, immunofluorescence, and real-time quantitative polymerase chain reaction. The results indicated that the optimal intervention condition involved serum containing 20% SGJPF, which was administered for a duration of 48 h. SGJPF-containing serum has been shown to reduce the viability of JS-1 cells and decrease the cell count in the G2 + S phase, thereby affecting HSC function. Furthermore, SGJPF-containing serum effectively suppressed the expression levels of NLRP3, IL-1β, IL-18, collagen I, α-SMA, circAno6, and TLR4 while upregulating miR-296-3p expression. These findings suggest that SGJPF can modulate the activity of the circAno6/miR-296-3p/TLR4 signaling axis, attenuate inflammatory responses, and inhibit HSC activation, thereby mitigating LF.
Keywords
Introduction
Liver fibrosis (LF), characterized by fibrotic and inflammatory processes, is a consequence of chronic liver injury (Zhang et al., 2016). Following hepatic damage, inflammatory cytokines stimulate the activation and transdifferentiation of hepatic stellate cells (HSCs) into myofibroblasts. These myofibroblasts subsequently produce collagen type I (collagen I) and α-smooth muscle actin (α-SMA), thereby facilitating the progression of LF (Chen et al., 2022; Liu et al., 2022).
As a novel form of endogenous noncoding RNA, circular RNAs (circRNAs) exhibit highly conserved and tissue-specific expression patterns and are involved in the regulation of transcription, translation, protein modification, and other processes (Li et al., 2023; Zhang et al., 2023). A substantial body of evidence indicates that circRNAs are significantly involved in the onset and progression of LF through mechanisms such as inflammatory damage, the activation of HSCs, and the acceleration of the fibrotic process (Jin et al., 2023; Xu et al., 2023). On the basis of our previous research, the pathway involving miR-296-3p and Toll-like receptor 4 (TLR4), in conjunction with circAno6, may serve as a regulatory mechanism that impacts the inflammatory response, which, in turn, leads to the activation of HSCs. Consequently, targeting circAno6 might be a promising therapeutic strategy for the treatment of LF (Li et al., 2024).
Shugan Jianpi Formula (SGJPF), also known as “Gan Le granule” or “Gan Le infusion,” was developed by the First Affiliated Hospital of Anhui University of Chinese Medicine for the treatment of LF (Jiang et al., 2017a). SGJPF is composed of 11 distinct herbs. To ensure quality, a fingerprint profile of the SGJPF extracts was established utilizing ultra-performance liquid chromatography in our previous study. Comprehensive details can be found in the Supplementary Data. SGJPF has been demonstrated to reduce inflammatory factor levels and inhibit both the activation and apoptosis of HSCs (Fan et al., 2020; Zhou et al., 2025). Furthermore, our previous research revealed that SGJPF mitigated LF induced by CCL4 in both mice and rats (Jiang et al., 2017a). These findings suggest that SGJPF may have therapeutic potential for the treatment of LF. However, the precise mechanisms through which SGJPF exerts its therapeutic effects on LF remain largely unclear.
Accordingly, the aim of this study was to investigate the potential mechanism by which SGJPF modulates the circAno6/miR-296-3p/TLR4 signaling axis, reduces proinflammatory factor production, and prevents the overactivation of HSCs, which could offer therapeutic benefits for LF therapy.
Materials and Methods
Cell culture
JS-1 mouse HSCs were obtained from Hunan Fenghui Biotechnology Co., Ltd., and cell culture techniques were conducted in accordance with established protocols (Zhang et al., 2020). To activate the JS-1 cells for modeling purposes, they were maintained in medium supplemented with 10 ng/mL transforming growth factor-β1 for 48 h (Peng et al., 2018).
Preparation of medicinal serum
Healthy male specific pathogen-free (SPF) Sprague–Dawley (SD) rats, each weighing ∼200 g with a tolerance of ± 20 g, were obtained from Liaoning Changsheng Biotechnology Co., Ltd. (SCXK2020-0001). All animal experiments were approved by the Experimental Animal Ethics Committee of Anhui University of Chinese Medicine (Ethics Approval No. AHUCM-rats-2024011). Thirty SPF-grade male SD rats were randomly assigned to a blank group (n = 10) and an SGJPF drug-containing serum group (n = 20). The rats in the SGJPF group were gavaged at 20 times the clinical dose, with 2 mL per 100 g given daily for 7 days. One hour after the final administration, the drug-containing serum was prepared according to the method described in the literature (Liu et al., 2025; Zhou et al., 2025). Finally, the serum was inactivated for 30 min in a water bath set to 56°C.
Cell Counting Kit-8
Cell Counting Kit-8 (CCK8) test kits (Servicebio, G4103) were used to evaluate the cell viability of normal resting HSCs and activated HSCs according to the manufacturer’s instructions.
Flow cytometric analysis
The cells were preserved in 90% ethanol at −20°C for 1 h after being rinsed with phosphate-buffered saline (PBS). The subsequent step involved washing the cells three times with PBS and resuspending them in a staining solution of propidium iodide and RNase, including 20 μL of RNase A. Prior to analysis using a BD LSRII flow cytometer, the samples were incubated for 30 min at 37°C. The resulting data were analyzed using FlowJo version 10.
Enzyme-linked immunosorbent assay
The levels of interleukin (IL)-1β and IL-18 present in the supernatants of the cell culture media were quantified using specific enzyme-linked immunosorbent assay kits designed for these cytokines. The kits employed in this study were RX203063M (for IL-1β) and RX203064M (for IL-18), which were obtained from RuiXin Biotech. All measurements were performed strictly according to manufacturer’s protocols to ensure the accuracy and reliability of the results obtained.
Real-time quantitative polymerase chain reaction
TRIzol (Invitrogen) was used to extract total RNA. The relative quantification system and the 2−ΔΔCt method were used for gene expression analysis, with β-actin as the reference gene. Primer Premier 5.0 software was used to design primers for NOD-like receptor protein 3 (NLRP3), collagen I, α-SMA, TLR4, and circAno6, whereas the stem–loop method was employed to design miR-296-3p primers. The primers used for PCR are listed in Table 1.
The Primer Sequence

α‐SMA, α‐smooth muscle actin; NLRP3, NOD‐like receptor protein 3; TLR4, Toll‐like receptor.
Western blotting
Cells were lysed in radioimmunoprecipitation assay (RIPA) buffer and subjected to centrifugation, followed by protein separation via sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS–PAGE). The proteins were then transferred to polyvinylidene fluoride (PVDF) membranes to obtain total protein. The membranes were incubated overnight with primary antibodies specific for several proteins, including β-actin, collagen I, NLRP3, TLR4, and α-SMA. The concentrations used for these antibodies were 1:1000 for β-actin, 1:500 for collagen I, NLRP3, and TLR4, and 1:2000 for α-SMA. Prior to this incubation, the membranes were treated with a blocking solution containing 5% skim milk powder for 2 h to prevent nonspecific binding. The secondary antibody was incubated following the same protocol and proportion for a duration of 2 h. Subsequently, the enhanced chemiluminescence (ECL) luminescent solution was prepared at a 1:1 ratio and applied to the PVDF membrane, and the signals were analyzed using the appropriate instrumentation.
Immunofluorescence experiments
The cells were fixed with a 4% paraformaldehyde solution at room temperature for 10 min. After fixation, the cells were permeabilized for 15 min. To avoid nonspecific antibody binding in later stages, a blocking solution containing 5% goat serum diluted in PBS was applied to the cells at room temperature for 1 h. Afterward, the cells were incubated with the primary antibody, followed by the application of the secondary antibody. The cell nuclei were stained using 4,6-diamidino-2-phenylindole, after which the cells were washed twice with PBS. After being mounted with DABCO (Sigma-Aldrich), the samples were subsequently examined using confocal laser scanning microscopy at 568 nm (Olympus, Tokyo, Japan).
Statistical analysis
Statistical analysis of the data was conducted using SPSS Statistics version 25.0, and the findings are expressed as the mean values accompanied by their respective standard deviations. To assess differences between two groups, a Student’s t-test was carried out. Furthermore, for comparisons involving multiple groups, a one-way analysis of variance was applied. A p value < 0.05 was considered indicative of a statistically significant difference in the results.
Results
Screening for the optimal concentration and treatment time of SGJPF-containing medicinal serum
According to the results of the CCK-8 assay, SGJPF-containing serum exhibited inhibitory effects on the viability of HSCs at various concentrations. When analyzed at different concentrations and time intervals, the optical density (OD) values at 48 and 72 h did not show any significant difference. However, a statistically significant difference was observed between 24 and 48 h, indicating that the inhibitory effect of SGJPF-containing serum on HSCs was greatest at 48 h (Fig. 1A). Furthermore, as illustrated in Figure 1B, the IC50 value of SGJPF-containing serum at 48 h was determined to be 20.3%. Consequently, a serum concentration of 20% SGJPF was identified as the optimal intervention condition for subsequent experiments following a 48-h exposure period. Notably, the selected condition (20% serum, 48 h) did not significantly affect normal resting HSCs (Supplementary Fig. S1), indicating that the subsequent observed effects were due to therapeutic inhibition rather than general cell cytotoxicity.

Screening of optimal intervention conditions of SGJPF medicinal serum.
Effect of SGJPF on the function of HSCs
To examine the effects of serum containing SGJPF on HSC viability and the cell cycle, CCK8 assays were conducted, as depicted in Figure 2A–C. Interestingly, a notable increase in HSC viability was observed in the model group compared with the control group, along with a marked decrease in the number of cells present in the G1 phase of the cell cycle. In contrast, compared with the model group, HSC viability in the SGJPF-containing serum and sh-circAno6 groups decreased by 22.6% and 34.8%, respectively. Additionally, there was a significant increase in the proportion of cells in the G1 phase, accompanied by a decrease in the proportion of cells in the G2 + S phases, which decreased by 20.1% and 28.8%, respectively.

Effect of SGJPF on the function of HSCs.
The outcomes of the IF, real-time quantitative polymerase chain reaction (RT-qPCR), and Western blotting (WB) assays are shown in Figure 2D–H. Compared with the control group, the model group demonstrated a marked increase in the fluorescence intensity as well as mRNA and protein expression levels. In contrast, compared with the model group, the groups treated with serum containing SGJPF and sh-circAno6 showed significant decreases in α-SMA fluorescence intensity as well as mRNA and protein expression levels. A similar pattern was observed for collagen I, as illustrated in Figure 2I–M.
Effect of SGJPF on inflammatory markers
As illustrated in Figure 3A–E, compared with the control group, the model group had significantly higher NLRP3 fluorescence intensity, as well as elevated mRNA and protein expression levels. In contrast, compared with the model group, both the SGJPF-containing serum group and the sh-circAno6 group exhibited significant decreases in NLRP3 fluorescence intensity as well as mRNA and protein expression levels. Figure 3F–G illustrates that compared with the control group, the levels of IL-1β and IL-18 were significantly increased in the model group, whereas the levels in the SGJPF-containing serum and sh-circAno6 groups were significantly reduced compared with the model group.
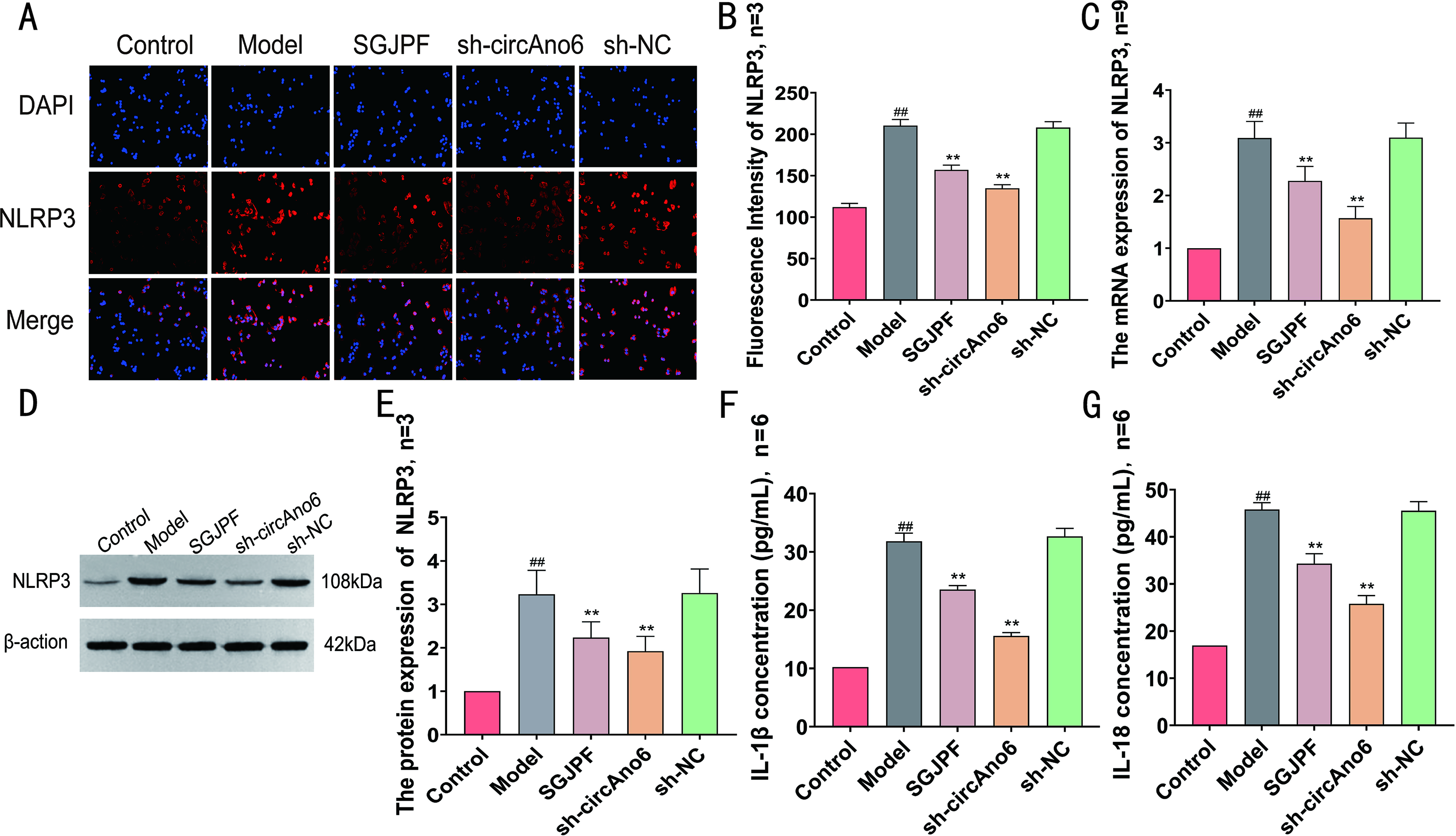
Effect of SGJPF on inflammatory markers.
Effect of SGJPF on the circAno6/miR-296-3p/TLR4 signaling axis
The findings from the RT-qPCR analysis revealed a significant increase in circAno6 levels in the model group along with a considerable decrease in the expression of miR-296-3p. In contrast, compared with the model group, both the SGJPF-containing serum group and the sh-circAno6 group exhibited significantly lower circAno6 levels. Conversely, a notable increase in miR-296-3p expression levels was detected in these groups (Fig. 4A, B). IF, RT-qPCR, and WB assays revealed significant increases in TLR4 fluorescence intensity as well as mRNA and protein expression levels in the model group. However, different trends were observed between the SGJPF-treated and sh-circAno6 groups (Fig. 4C–G).
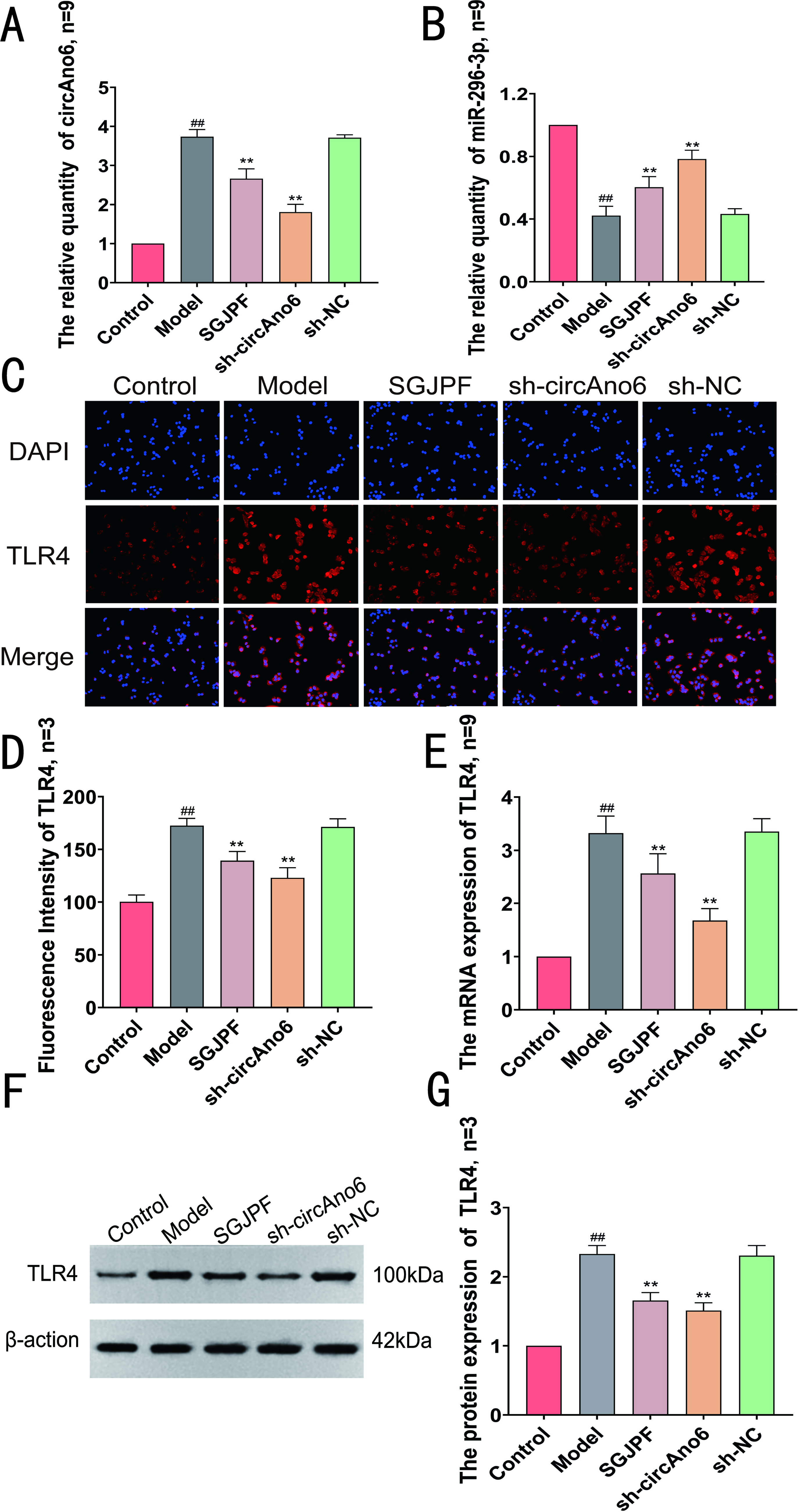
Effect of SGJPF on circAno6/miR-296-3p/TLR4 signaling axis.
SGJPF can rescue the effects of circAno6 overexpression on HSC function and inflammatory reactions
Finally, to investigate whether SGJPF can rescue HSC function and the inflammatory response by regulating circAno6, we conducted a recovery experiment. Notably, as illustrated in Figure 5A–E, both the model group and the pc-circAno6 groups exhibited an increase in cell viability, whereas the overexpression group that received SGJPF treatment reversed this trend. Furthermore, flow cytometry analysis indicated that SGJPF effectively rescued cell cycle arrest in the G2 + S phase induced by circAno6 overexpression. Moreover, SGJPF significantly reduced the levels of the inflammatory cytokines IL-1β and IL-18, thereby alleviating the inflammatory response.

SGJPF can rescue the effects of overexpression of circAno6 on the HSC function and inflammatory reaction.
Discussion
Traditional Chinese medicine(TCM) has long been utilized as a therapeutic strategy for addressing LF (Tao et al., 2019). Notably, owing to its comprehensive approach and syndrome differentiation, TCM presents distinct advantages in LF management (Jiang et al., 2017b). The SGJPF, a classical TCM formulation, is composed of Chaihu (CH, Bupleuri Radix), Baishao (BS, Paeoniae Radix Alba), Zelan (ZeL, Lycopus lucidus Turcz), Baizhu (BZ, Atractylodis Macrocephalae Rhizoma), Fuling (FL, Poria), Huangqi (HQ, Astragali Radix), Yiyi Ren (YYR, Coicis Semen), Banlan Gen (BLG, Isatidis Radix), Yinchen (YC, Artemisiae Scopariae Herba), Zhuling (ZL, Polyporus), and Zhiqiao (ZQ, Aurantii Fructus) and has been extensively utilized in the clinical treatment of liver disorders. This investigation revealed that, by regulating the circAno6/miR-296-3p/TLR4 signaling axis, SGJPF can inhibit the production of proinflammatory factors and prevent excessive activation of HSCs, potentially explaining its mechanism in LF treatment.
First, our experimental results revealed that SGJPF significantly reduced the viability of HSCs and increased the proportion of HSCs in the G1 phase of the cell cycle. These observations suggest that SGJPF can inhibit both cell viability and proliferation. Considering the well-established link between LF and the activation and proliferation of HSCs, it is evident that the activation of HSCs results in the upregulation of α-SMA expression and overproduction of collagen, thereby aggravating the progression of LF (Carson et al., 2018). Subsequently, we found that SGJPF-containing serum can significantly reduce the levels of α-SMA and collagen I.
Numerous studies have established a significant association between circRNAs and the occurrence and progression of LF (Chen et al., 2017; Cheng et al., 2018; Jin et al., 2023). Specifically, circAno6 has been identified as a differentially expressed circRNA that is closely linked to LF (Zhou et al., 2019). In our previous work, we demonstrated the involvement of the circAno6/miR-296-3p/TLR4 signaling axis in triggering inflammatory responses and contributing to LF pathogenesis (Li et al., 2024). Notably, serum containing SGJPF significantly reduced the level of circAno6 expression, which indicates that SGJPF may inhibit LF through the modulation of circAno6.
MiR-296-3p is part of the miR-296 gene cluster, which is involved in regulating numerous physiological and pathological functions, such as cell growth, migration, and programmed cell death, and has been linked to the development of several diseases (Bravo-Parra et al., 2020; Wang et al., 2019). Initial experimental findings revealed the presence of a miR-296-3p binding site within the circAno6 sequence, suggesting a potential interaction between circAno6 and miR-296-3p (Li et al., 2024). Moreover, the findings indicated that SGJPF promoted miR-296-3p expression, suggesting that SGJPF exerts a regulatory effect on miR-296-3p.
TLR4 is a type I transmembrane receptor located on the cell membrane that consists of extracellular, transmembrane, and intracellular domains. It is instrumental in the pathogenesis of LF (Li et al., 2021). A substantial body of academic research has demonstrated that TLR4 is capable of initiating inflammatory responses, encouraging HSC activation and proliferation, and inducing excessive ECM production and secretion, which facilitates the onset and progression of LF (Jourdan et al., 2013; Li et al., 2021; Wu et al., 2018; Zhou et al., 2022). TLR4 is integral to the progression of LF, with evidence suggesting that inhibition of the TLR4 signaling pathway mitigates liver injury and subsequently reduces fibrosis (Liang et al., 2016; Tian et al., 2017). In this study, we discovered that SGJPF significantly reduced TLR4 expression. On the basis of previous research, it is plausible to speculate that SGJPF modulates LF by regulating the circAno6/miR-296-3p/TLR4 signaling axis.
Liver inflammation originates from the production of proinflammatory cytokines that promote the activation of HSCs and contribute to the progression of LF and cell death linked to NASH (Manne et al., 2018). Studies have indicated that IL-1β enhances LF, whereas IL-18 belongs to the IL-1 superfamily of cytokines and has a structural resemblance to IL-1β (Arend et al., 2008; Kaplanski, 2018). The NLRP3 inflammasome is essential in various liver conditions, such as fatty liver disease, drug-induced liver injury, ischemia/reperfusion damage, and fibrosis (Artlett, 2012; Henao-Mejia et al., 2012; Zhu et al., 2011). Research has provided empirical evidence indicating that the NLRP3 inflammasome activates caspase-1. This activation is a crucial step in a biochemical cascade that leads to the maturation of cytokines. Specifically, caspase-1 catalyzes the conversion of pro-IL-1β and pro-IL-18, the inactive precursors of these important inflammatory cytokines, into their active forms, IL-1β and IL-18. This process is essential for the initiation and regulation of inflammatory responses (Essam et al., 2019; Gan et al., 2022; Li et al., 2019; Tavakoli Dargani and Singla, 2019). The results of this study revealed that SGJPF significantly decreased the levels of IL-1β, NLRP3, and IL-18, indicating its potential to suppress inflammatory responses. Notably, TLR4 plays a critical role in activating the NLRP3 inflammasome by delivering a strong priming signal that facilitates the synthesis of new pro-IL-1β and increases the expression level of NLRP3. This process is vital for triggering the inflammasome and maturation of the proinflammatory cytokine IL-1β (Tsutsui et al., 2010).
Conclusion
Consequently, as depicted in Figure 6, SGJPF exerts anti-LF effects through modulation of the circAno6/miR-296-3p/TLR4 signaling axis, attenuation of inflammatory responses, and inhibition of HSC activation, thereby ultimately mitigating LF.
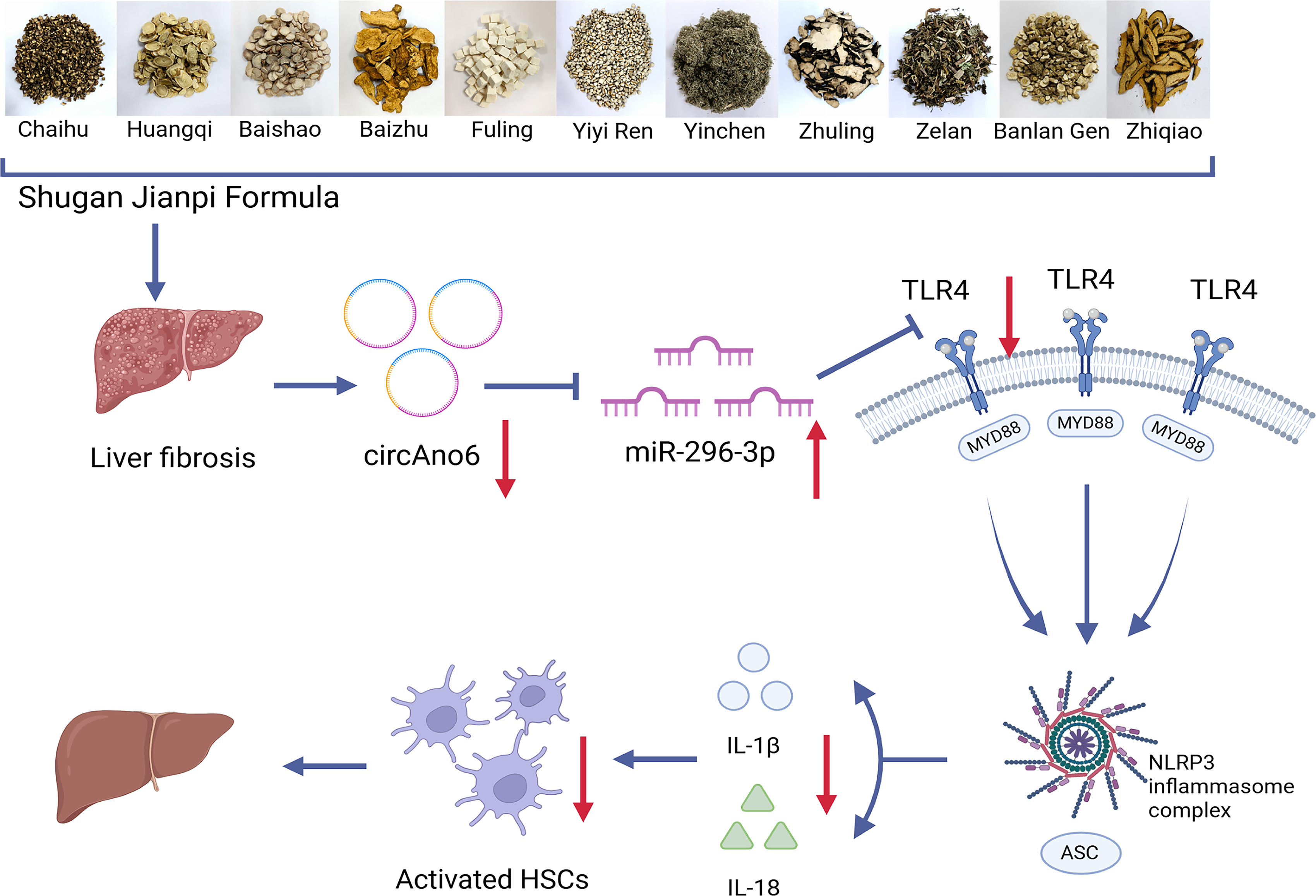
Mechanism diagram of regulating the circAno6/miR-296-3p/TLR4 signaling axis with SGJPF to alleviate inflammation cascade and inhibit HSCs activation.
Authors’ Contributions
Q.W. and T.L.: Writing—original draft. J.Q.: Data curation. C.F.: Supervision and funding acquisition. L.Z.: Supervision and writing—review and editing. H.J.: Funding acquisition, supervision, writing—review and editing, and conceptualization.
Footnotes
Acknowledgments
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by the
Disclosure Statement
The authors affirm that they have no recognized financial conflicts of interest or personal relationships that could have potentially influenced the research presented in this article.
Funding Information
This study was funded by National Natural Science Foundation of China (grant no. 82304995) and Natural Science Research Project of Anhui Educational Committee (grant no. 2024AH040165).
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.