
Abstract
Learning and memory were long thought to be the domains of neurons and neural networks, but recent work across brain systems has established that astrocytes play a role in synaptic plasticity and information storage. However, it remains unclear what this role is. In Hebbian and spike timing–dependent plasticity, simultaneous activation of pre- and postsynaptic neurons on a millisecond timescale determines plasticity, yet emerging evidence suggests that slow astrocyte signaling over many seconds is directly linked to memory. Here we take an astrocyte-centric view on plasticity to explore the possible computational and coding roles of astrocytes. We propose that in a 3-body arrangement with pre- and postsynaptic compartments, astrocytes may participate in the synaptic credit assignment problem. This review thus offers a candidate conceptual framework for understanding the interplay between astrocytes and neurons in the context of neural network performance and plasticity.
Introduction
Historically, the roles of astrocytes were considered to be restricted to metabolic support, ionic homeostasis, and neurotransmitter clearance from the synaptic cleft (Volterra and Meldolesi 2005; Halassa et al 2007a). Learning and memory have thus long been thought of as chiefly the domain of neurons and their synaptic connections, thereby establishing a neurocentric view of information storage in the brain. More recently, however, astrocytes have been shown to contribute actively to synaptic plasticity, critical period development, and memory formation (Perea et al 2009; Andrade-Talavera et al 2023b; Murphy-Royal et al 2023; Sánchez Romero and Navarrete 2026).
As the neurocentric view of learning and memory broadens to include a more astrocentric perspective, we first provide a brief overview of the current state of the field. We then frame synaptic plasticity as a 3-body problem, emphasizing the added complexity that arises when astrocytes dynamically interact with pre- and postsynaptic compartments. We focus in particular on how astrocytes might regulate long-term synaptic plasticity, including Hebbian plasticity (Hebb 1949) and spike timing–dependent plasticity (STDP; Markram et al 2012)—2-factor learning rules once thought to be the sole domain of neurons (see Box 1 for a multibody perspective). Finally, we propose that the role of astrocytes in synaptic plasticity may help explain how the brain resolves the credit assignment problem (Box 2).
A multibody perspective on 1-, 2-, and 3-factor learning rules.

Credit assignment in artificial and biological neural networks.

Plasticity in Learning and Memory
Learning and memory are grounded in the brain’s ability to change in response to experience. At the synaptic level, this adaptive capacity is captured by Donald Hebb’s postulate (Hebb 1949), which states that when cooperating presynaptic neurons repeatedly and persistently drive postsynaptic spiking, connections between pre- and postsynaptic cells should be strengthened. These cooperating cells that fire together thus wire together with recipient cells (Shatz 1992). This way, Hebbian plasticity provides a biological substrate for the formation of cell assemblies that represent learned information by forming neural traces, or engrams, of stored memories.
A modern cellular implementation of Hebb’s idea is STDP, where the millisecond temporal order of pre- and postsynaptic spikes governs the direction and magnitude of synaptic change (Markram et al 2012). Typically, long-term potentiation (LTP) is elicited for pre- before postsynaptic spiking within a 10-ms-long temporal window, whereas the opposite temporal ordering yields long-term depression (LTD; Markram et al 2012). As for Hebbian plasticity, presynaptic inputs that help elicit postsynaptic spiking are strengthened. In STDP however, late-arriving inputs from presynaptic cells that therefore fail to drive postsynaptic spiking are additionally weakened. This extension to Hebb’s postulate is important because it enables competition among synaptic inputs (Song et al 2000). STDP has furthermore been widely observed across brain regions and has been linked to connectivity refinement and memory encoding (Markram et al 2012). STDP is consequently widely thought of as a biologically plausible learning rule.
Yet, despite its broad appeal, STDP has been rightfully criticized (Lisman and Spruston 2005, 2010). For instance, its reliance on millisecond-scale spike timing may not align well with the much longer behavioral timescales of learning. More recent work in the hippocampus has uncovered behavioral timescale synaptic plasticity (BTSP), for which bidirectional synaptic changes occur over seconds rather than milliseconds, supporting the formation of place fields (Bittner et al 2017; Milstein et al 2021). Together, STDP and BTSP extend Hebbian principles across timescales and temporal ordering of activity in pairs of connected neurons (Suvrathan 2019).
Conceptually, this invites a computational interpretation: Hebbian-like learning rules such as STDP and BTSP are cases of unsupervised learning, as they depend on local correlations in connected neurons, without reference to explicit reward signals or overall error of the neuronal circuit. They are, in other words, 2-factor learning rules defined by pre- and postsynaptic activity (Box 1). In light of this, it is perplexing why and how the tripartite synapses that involve astrocytes should contribute actively to synaptic plasticity (Perea et al 2009).
In parallel with Hebbian and timing-based plasticity, which primarily serve to encode information by strengthening or weakening selected synapses, the brain employs mechanisms of homeostatic plasticity whose role is not to store memories but to stabilize overall network excitability, ensuring that neuronal activity remains within an optimal dynamic range (Turrigiano 2007, 2011). In homeostatic plasticity, neurons change the excitability of their synapses in response to overall network excitability. In contrast to Hebbian-like learning rules—which are correlation-based 2-factor mechanisms—homeostatic synaptic scaling operates as a 1-factor learning rule, driven solely by deviations of neuronal activity from a target set point (Turrigiano 2007, 2011). Here, it has long been known that glia cells play central roles; for example, astrocyte-derived TNF-α is key to synaptic upscaling (Stellwagen and Malenka 2006). Glial cells are clearly not merely supportive but active contributors to processes that stabilize Hebbian-like learning rules.
For a synapse to be aware of overall network performance and behavioral outcomes, Hebbian-like 2-factor learning is not enough, since the information available to the synapse is only local. Here is where 3-factor learning may provide a solution to improving behavioral learning and network error rates, by gating local synaptic plasticity (Box 1), which can be interpreted as 3 interacting bodies: presynaptic, postsynaptic, and astrocytic compartments. The third factor could arise in the form of network state, reward feedback, or neuromodulation. Three-factor rules can therefore help implement forms of synaptic learning that address the credit assignment problem (Box 2).
Recent studies have brought to light that astrocytes may act as a factor in addition to pre- and postsynaptic activity to regulate plasticity, contributing to a 3-factor learning rule. For example, at hippocampal CA3 → CA1 synapses, a developmental switch occurs after the fourth postnatal week, whereby the same temporal pairings of postsynaptic activity before presynaptic activity that had previously induced timing-dependent LTD (tLTD) up to the third postnatal week now induce timing-dependent LTP (tLTP; Falcón-Moya et al 2020). A similar phenomenon was observed in layer 4 (L4) → L2/3 somatosensory cortex (Martinez-Gallego et al 2022). In both cases, the developmental switch and expression of tLTP were found to critically rely on the release of adenosine and glutamate from astrocytes, which provided the third factor for this form of plasticity (Perez-Rodriguez et al 2019; Falcón-Moya et al 2020; Martinez-Gallego et al 2022).
Taken together, synaptic plasticity encompasses a spectrum of mechanisms—from Hebbian-like 2-factor rules such as STDP and BTSP that encode information to 1-factor homeostatic scaling that stabilizes neuronal firing. Recognizing that these rules involve not only pre- and postsynaptic compartments but also astrocytic processes forces a conceptual shift: learning and memory may not be a 2-body but a 3-body problem of brain computation. This raises a key question: if 1- and 2-factor rules suffice for encoding and stabilization, what computational role does the third astrocytic compartment serve? This question motivates reexamination of plasticity from an astrocentric perspective.
Astrocyte Involvement in Synaptic Plasticity
Since the 2000s, astrocytes have been shown to contribute to different forms of LTP and LTD (Nishiyama et al 2002; Pascual et al 2005; Perea and Araque 2007). These long-lasting changes in synaptic strength are often classified by their dependence on particular receptor mechanisms and signaling pathways. In this view, astrocytes are not merely passive bystanders at synapses but can be active participants in determining whether plasticity occurs and sometimes even in shaping which form of plasticity is expressed.
The involvement of astrocytes in long-term plasticity extends across a variety of brain regions and can act through the release of diverse regulatory molecules, often referred to as gliotransmitters, including
Taking astrocyte-derived glutamate signaling as an example, astrocytes have been reported to modulate LTP (Perea and Araque 2007; Navarrete et al 2012; Park et al 2015) and LTD (Han et al 2012; Min and Nevian 2012; Navarrete et al 2019; Martinez-Gallego et al 2024). In other words, astrocytic signaling has been implicated not only in scaling plasticity up or down but also in shaping whether synaptic strengthening or weakening is expressed under a given induction paradigm.
Communication at the Tripartite Synapse
Astrocytes are integral components of the tripartite synapse and can influence long-term plasticity through pre- and postsynaptic mechanisms. Seminal work showed that astrocytic processes closely appose presynaptic boutons and postsynaptic dendrites, forming a functional neuron–glia unit known as the tripartite synapse (Bushong et al 2004; Halassa et al 2007b). This arrangement places astrocytes in a strategic position to shape bidirectional signaling at synapses.
At tripartite synapses, astrocytes detect neuronal activity and extracellular neurotransmitters via metabotropic and ionotropic receptors, engaging intracellular signaling pathways. In addition, transporters of neurotransmitters and ion channels on astrocyte membranes have been shown to regulate synaptic transmission. For example, glutamate transporters on astrocytes shape neurotransmission, and astrocyte Kir4.1 channels are local regulators of synaptic plasticity (Michaluk et al 2021; Tyurikova et al 2025). Their architecture (Box 3) implies that an individual astrocyte can influence plasticity not only at isolated synapses but across synaptic ensembles embedded within a shared astrocytic territory. Consistent with this view, astrocytes exhibit selective responses to specific cell and synapse types (Mariotti et al 2018; Matos et al 2018), with functional heterogeneity emerging among local astrocyte populations (Martin et al 2015). Ultrastructural studies also describe synaptic clusters organized within astrocyte territories (Salmon et al 2023; Benoit et al 2025), supporting the idea that astrocytes coordinate signaling at spatial scales distinct from purely neuronal circuit architectures.
Astrocyte architecture as a substrate for synaptic plasticity.

These signaling processes can trigger regulated release of gliotransmitters from astrocytes via calcium-dependent and calcium-independent mechanisms, which thereby act on neurons (Goenaga et al 2023). While this review focuses on evidence showing how astrocyte regulates plasticity, it is worth noting that there is ongoing debate regarding gliotransmission and the mechanisms of astrocyte–neuron communication (Sloan and Barres 2014; Chai et al 2017; Fiacco and McCarthy 2018; de Ceglia et al 2023).
Collectively, these properties suggest that astrocytes help determine when and where long-term synaptic plasticity is permitted, positioning them as plausible mediators of a third factor in 3-factor learning rules. In this view, astrocytes regulate the local permissive state that determines when neuron–neuron interactions produce lasting synaptic change.
Presynaptic Mechanisms
Presynaptic expression of long-term plasticity refers to lasting changes in neurotransmitter release from the presynaptic terminal. In practice, this typically reflects altered release probability, reflecting altered vesicle release machinery, presynaptic calcium dynamics, or both (Monday et al 2018).
Evidence across brain regions points to a recurring motif in which astrocytes relay postsynaptic signals back to presynaptic terminals, thereby permitting long-term changes in release probability. At hippocampal synapses, one established route for tLTD begins with glutamate release from the presynaptic terminal, activating postsynaptic metabotropic glutamate receptors (Andrade-Talavera et al 2016). This triggers calcium release from internal stores and promotes endocannabinoid (eCB) synthesis. The eCB signal then engages cannabinoid type 1 receptors on astrocytes, inducing astrocytic
Related mechanisms have been proposed in multiple circuits. In entorhinal–hippocampal pathways, astrocytes respond to postsynaptic eCB signaling by releasing glutamate that may activate presynaptic NMDA receptors, decreasing release probability and contributing to tLTD (Martinez-Gallego et al 2024). Similar motifs appear in neocortex: in barrel cortex, tLTD and astrocyte-induced LTD converge on presynaptic mechanisms dependent on presynaptic NMDA receptors (Min and Nevian 2012). In visual cortex, astrocytic CB1Rs and calcium signaling may likewise determine presynaptic NMDA receptor–dependent tLTD in L5 (Sjöström et al 2003; Thomazeau et al 2025; Watanabe et al 2026), and chemically induced eCB LTD in somatosensory cortex requires astrocytes and presynaptic NMDA receptors (Neubauer et al 2022).
Presynaptic depression can arise through adenosine signaling. At corticostriatal synapses, high-frequency stimulation activates astrocytic metabotropic glutamate receptors, triggering adenosine release that engages presynaptic A1 receptors and produces LTD (Cavaccini et al 2020).
Together, these findings indicate that astrocytes can govern presynaptic plasticity by linking postsynaptic activity to presynaptic release machinery through signals such as nitric oxide, eCBs,
Postsynaptic Mechanisms
Postsynaptic expression of long-term plasticity is typically mediated by changes in the number, properties, or regulation of postsynaptic receptors. These modifications alter synaptic gain, often through changes in quantal amplitude arising from receptor phosphorylation, conductance shifts, or receptor trafficking (Huganir and Nicoll 2013).
Across brain regions, a recurring theme is that astrocytes influence postsynaptic plasticity by regulating receptor activation and trafficking. In the somatosensory cortex, one form of postsynaptic tLTD arises when glutamate activates postsynaptic NMDA receptors and drives eCB synthesis (Andrade-Talavera et al 2024). This eCB signaling is detected by astrocytic CB1Rs, triggering
Astrocytic CB1Rs and metabotropic glutamate receptors are G protein–coupled receptors that can elevate intracellular calcium via production of inositol 1,4,5-triphosphate (Khakh and McCarthy 2015). Activation of 1,4,5-triphosphate receptors on the endoplasmic reticulum then releases calcium from internal stores. In the aforementioned examples, these calcium elevations are associated with gliotransmission onto neurons and contribute to plasticity induction or expression. Although astrocytes can also signal through calcium-independent pathways (Wang Z et al 2000; Goenaga et al 2023), many proposed astrocyte-dependent 3-factor plasticity mechanisms are thought to involve calcium-dependent gliotransmission.
Collectively, these mechanisms indicate that astrocytes participate in postsynaptic plasticity not only by modulating receptor activation in real time but also by influencing receptor trafficking processes that stabilize long-term synaptic change. Astrocytes may therefore regulate synaptic gain from the postsynaptic side by shaping the induction of plasticity and the stabilization of receptor changes that encode it. Astrocytes therefore regulate synaptic weight and synaptic number, linking functional plasticity to structural circuit remodeling (Farhy-Tselnicker and Allen 2018).
How Astrocytes Code in the Brain
Although astrocytes do not fire action potentials, they are active signaling elements that communicate with neurons and shape synaptic function (Nedergaard 1994; Parpura et al 1994; Volterra and Meldolesi 2005). In the field, astrocyte activity is commonly measured by intracellular calcium dynamics. There are other indicators of astrocyte activity, such as intracellular cAMP and sodium levels (Vardjan and Zorec 2015; Verkhratsky et al 2020; Rose and Verkhratsky 2024). Here, we focus on astrocyte calcium signaling and consider how it may contribute to synaptic plasticity across local and global scales.
Astrocyte Calcium Signaling
Astrocyte calcium signaling constitutes the primary physiological substrate through which astrocytes encode and transmit information. Changes in intracellular astrocyte calcium levels occur spontaneously (Nett et al 2002) or in response to neuronal activity (Perea and Araque 2005; Bernardinelli et al 2011). Spontaneous signals are generated intrinsically and often take the form of brief, localized events that can propagate once intracellular calcium crosses the threshold for calcium-induced calcium release (Semyanov et al 2020). These events are developmentally regulated, becoming less synchronous with maturation (Watanabe et al 2023; Figure 1), potentially reflecting a shift toward more spatially restricted control of synaptic function. One mechanism underlying spontaneous activity is the opening of TRPA1 channels in the astrocyte membrane (Shigetomi et al 2013).
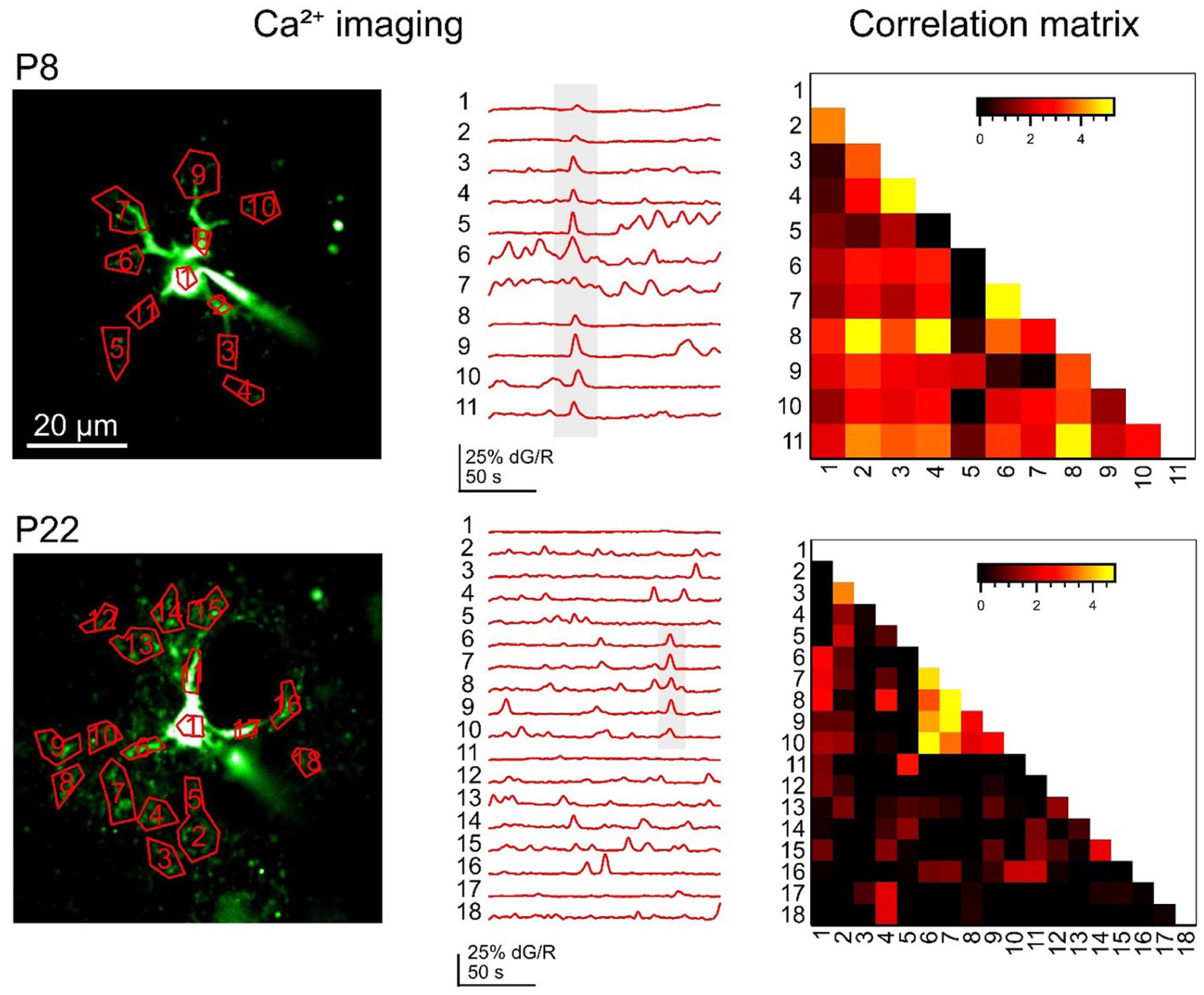
As astrocytes mature, spontaneous calcium activity decorrelates. Left: Region-of-interest segmentation of astrocytic calcium imaging signals shows more synchronous activity across multiple microdomains in a younger astrocyte (postnatal day 8 [P8], top) than in an older astrocyte (P22, bottom). Sample co-active events are highlighted in gray. Right: Pairwise correlation matrices across regions of interest show stronger microdomain correlations in the younger astrocyte (top) as compared with the older astrocyte (bottom). Adapted with permission from Watanabe et al (2023).
By contrast, activity-evoked calcium signaling ranges from rapid and spatially confined events (Stobart et al 2018) to prolonged signals spanning multiple astrocytes (Bernardinelli et al 2011). Such responses are triggered by receptor activation on the astrocyte membrane, including G protein–coupled receptors and NMDA receptors (Bernardinelli et al 2011; Ahmadpour et al 2024). When compared with calcium signals in neurons, those in astrocytes are relatively modest, up to a few hundred nanomolar, but are sufficient to trigger gliotransmission (Parpura and Haydon 2000).
Spatial organization is a defining feature of astrocyte calcium signaling. Calcium elevations may appear as discrete events within fine processes such as branchlets and leaflets or as larger oscillations involving branches and the soma (Box 3). These localized signals, often termed microdomains, can depend strongly on mitochondrial calcium release and may be largely independent of 1,4,5-triphosphate receptors (Agarwal et al 2017; Benoit et al 2025). Astrocytes are further coupled through gap junctions (Giaume et al 2010), enabling calcium signals to propagate across astrocyte networks and extend their sphere of influence.
Astrocyte calcium signals typically unfold over hundreds of milliseconds to seconds (Stobart et al 2018; Goenaga et al 2023). Even the fastest astrocyte calcium events (~100 ms) are much slower than neuronal action potentials and the timescale of STDP. This poses a central question: how can comparatively slow astrocyte signaling shape fast neuronal information processing and synaptic plasticity?
Finally, astrocyte calcium dynamics have been linked to behavior (Martin-Fernandez et al 2017; Yu et al 2018; Nagai et al 2021), suggesting that astrocytic signaling operates across multiple scales. Astrocyte calcium signals can thus originate in local microdomains yet, under behavioral demands, propagate toward somatic integration and broader network coordination.
Local to Global Astrocyte Signaling
Locally, individual astrocytes can contact large numbers of synapses within their territorial domain, and they can remodel their fine processes to alter synaptic coverage over time (Ventura and Harris 1999). Therefore, astrocytes can modulate plasticity at the level of synaptic ensembles within microdomains, rather than at isolated synapses. Consistent with this view, local microdomains of spontaneous calcium signals have been proposed to contribute to memory formation by supporting consolidation of LTP (Losi et al 2025).
Beyond this local scale, astrocyte calcium activity does not necessarily remain confined to distal processes. In response to behavioral events, calcium signals can propagate from fine peripheral processes toward the soma (Rupprecht et al 2024), suggesting that astrocytes can act as slow integrators of neuronal activity and behavioral state. This somatic integration was further shown to be regulated by input from the locus coeruleus (Rupprecht et al 2024), linking astrocyte signaling to global neuromodulatory control. On top of their integrative function, astrocytes discriminate incoming synaptic information in a cell- and synapse-specific manner (Perea and Araque 2005; Martin et al 2015; Mariotti et al 2018; Matos et al 2018). In line with this broader role, astrocytes have been proposed to encode long timescale behavioral variables, including reward location (Doron et al 2022), spatial information (Curreli et al 2022), motivational state (Gau et al 2024), and fear memory (Williamson et al 2024). This suggests the idea that astrocyte calcium signals enable local neurotransmitter events to influence neuronal activity on a global scale.
At the global scale, astrocyte networks allow calcium signals to extend across larger spatial domains. Cahill et al (2024) showed that when glutamate or GABA is applied by uncaging to a spatially restricted area on a single astrocyte, the surrounding astrocyte network responded with minutes-long calcium transients that spread beyond the stimulated cell. Notably, stimulation of the same astrocyte region, with either glutamate or GABA, activated largely distinct regions of the local network, with glutamate responses displaying more propagative activity than GABA responses (Cahill et al 2024). Thus, local transmitter release may drive astrocyte network activity in a neurotransmitter-specific manner (Figure 2). This highlights the importance of astrocyte network extent, as neurons that would not otherwise be directly influenced by activity in a spatially distant region may still be modulated indirectly through astrocyte-mediated signaling. Finally, astrocyte heterogeneity across brain regions suggests that no single mode of astrocyte-mediated plasticity dominates. Instead, distinct astrocyte populations deploy different strategies to shape synaptic change (Schroeder et al 2025).
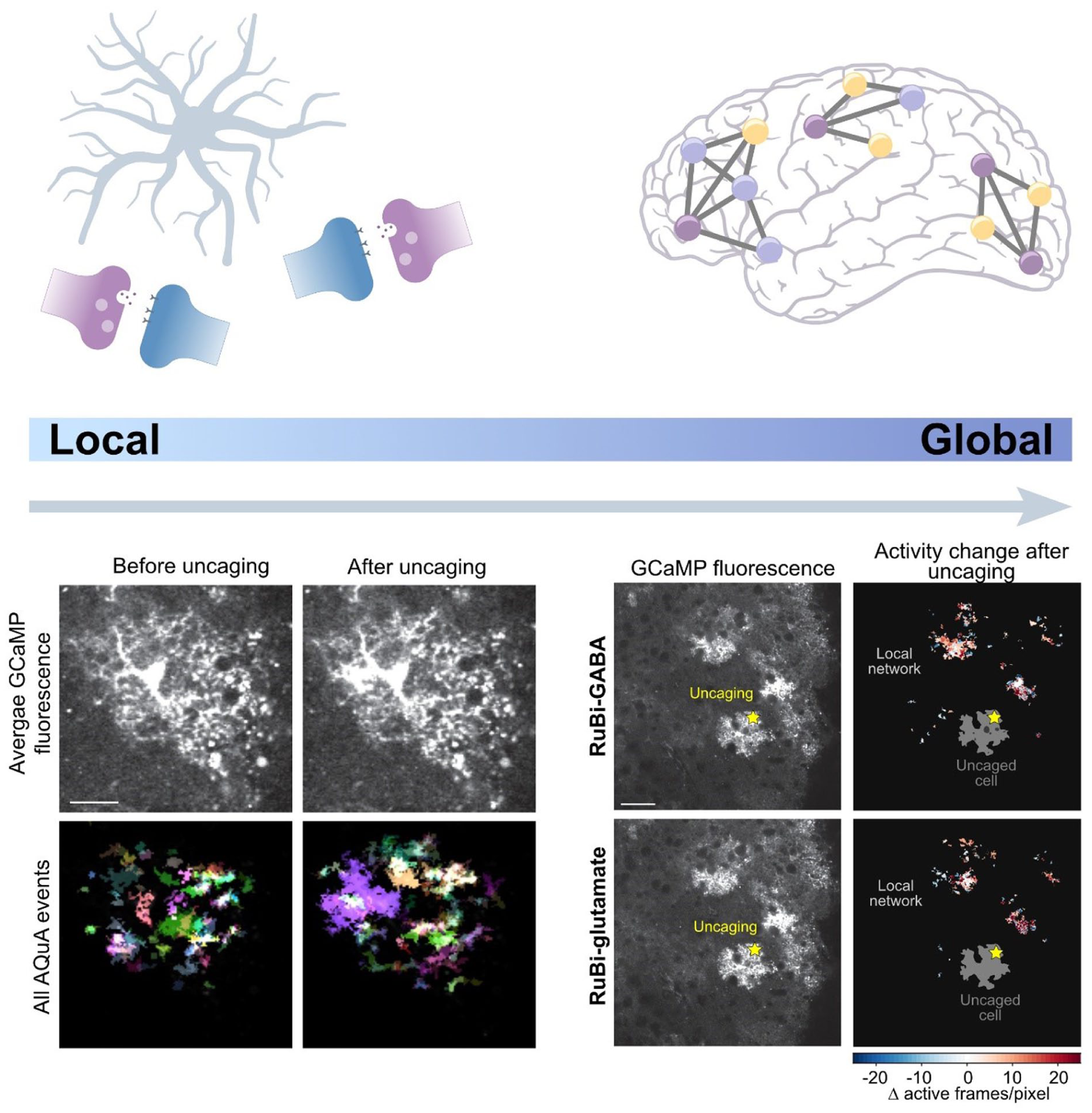
Local to global astrocyte signaling across brain networks. Top: Astrocytes can operate across spatial scales, from local modulation of nearby synapses to coordinated signaling across local astrocyte networks, although evidence for astrocyte networks covering brain regions is emerging (Cooper et al 2026). Bottom: Astrocyte calcium activity was monitored by GCaMP imaging. On the left, images before and after uncaging illustrate that direct stimulation of a single astrocyte via GABA uncaging evokes local increases in calcium activity and alters the spatial pattern of calcium activity. On the right, spatial heat maps show changes in astrocyte network activity following uncaging of either GABA or glutamate, demonstrating that local neurotransmitter stimulation can recruit broader network-level responses beyond the uncaged astrocyte. Scale bars: 20 and 50 µm. Adapted with permission from Cahill et al (2024).
It is worth noting that although astrocytes have long been considered electrically nonexcitable, neuronal activity can nonetheless evoke depolarizations in their fine peripheral processes. Such depolarizations have been shown to inhibit glutamate clearance by astrocytes, thereby revealing astrocyte–neuron coupling beyond calcium-based signaling (Armbruster et al 2022).
These local and network-level signaling modes suggest that astrocytes can integrate information across space and time beyond individual synapses. A natural next question is whether such integration can be organized at the level of astrocyte populations, forming coordinated patterns of activity linked to learning and memory. Recent work has begun to address this possibility by identifying astrocyte ensembles that are engaged during memory encoding and recall.
Astrocyte Ensembles in Learning and Memory
On a larger scale, emerging studies suggest that astrocytes can directly modulate memory. Experimental manipulation of astrocyte signaling pathways, including the activation of Gq protein–coupled receptor signaling and the elevation of intracellular cAMP, alters behavioral performance in memory assays (Adamsky et al 2018; Zhou et al 2021).
Recent work further implicates coordinated astrocyte populations, termed astrocyte ensembles, in memory recall. Williamson et al (2024) showed that fear conditioning induced c-Fos expression in a subset of hippocampal astrocytes that interacted closely with engram neurons. Selective reactivation of these astrocytes elicited freezing behavior consistent with fear memory recall. Serra et al (2025) used a light- and calcium activity–dependent labeling strategy to target astrocyte ensembles encoding a cue–reward association. In response to reward, astrocyte calcium responses gradually increased over 10 days of training, and reactivation of astrocyte ensembles biased behavior toward a reward-associated location. Extending this framework across broader spatial scales, Dewa et al (2025) performed brain-wide tagging of astrocytes during fear conditioning and found that astrocyte ensembles were transcriptionally primed during learning and functionally engaged during recall. Increased noradrenaline receptor expression on astrocytes occurred hours to days after learning, suggesting that astrocytes themselves can be plastic. Silencing these ensembles impaired memory stabilization during repeated recall, suggesting that astrocytes enable neuronal encoding and fear memory retrieval (Williamson et al 2024; Dewa et al 2025; Bukalo et al 2026). Together, these studies suggest that astrocytes participate in memory not only by modulating synapses locally but also through coordinated ensemble-level activity that permits learning and recall. Astrocytes may therefore contribute to memory across scales, from regulating individual synapses to coordinating distributed cellular ensembles.
Disruptions of astrocyte signaling have been linked to disease states involving memory impairment. For example, reduced spontaneous and evoked astrocyte calcium activity has been associated with memory loss and impaired long-term synaptic plasticity in Alzheimer disease (Lia et al 2023).
These observations motivate a shift from a solely neurocentric view toward one that incorporates astrocytic contributions to plasticity (Sánchez Romero and Navarrete 2026). In this framework, astrocytes integrate synaptic signals across their territorial domains and influence brain computation by controlling when and where plasticity is permitted.
What Are the Benefits of Involving Astrocytes in Plasticity?
Neurocentric and Astrocentric Perspectives
Neuroscience has traditionally adopted a neurocentric perspective in which synapses are treated primarily as neuron-to-neuron communication units (Zilberter et al 2009) and plasticity is viewed as a predominantly neuronal process. However, recent work suggests that astrocytes integrate synaptic signals within discrete subdomains (Figure 3) and that the number of synaptic inputs coordinated within a single astrocytic subdomain can be strikingly large (Benoit et al 2025).
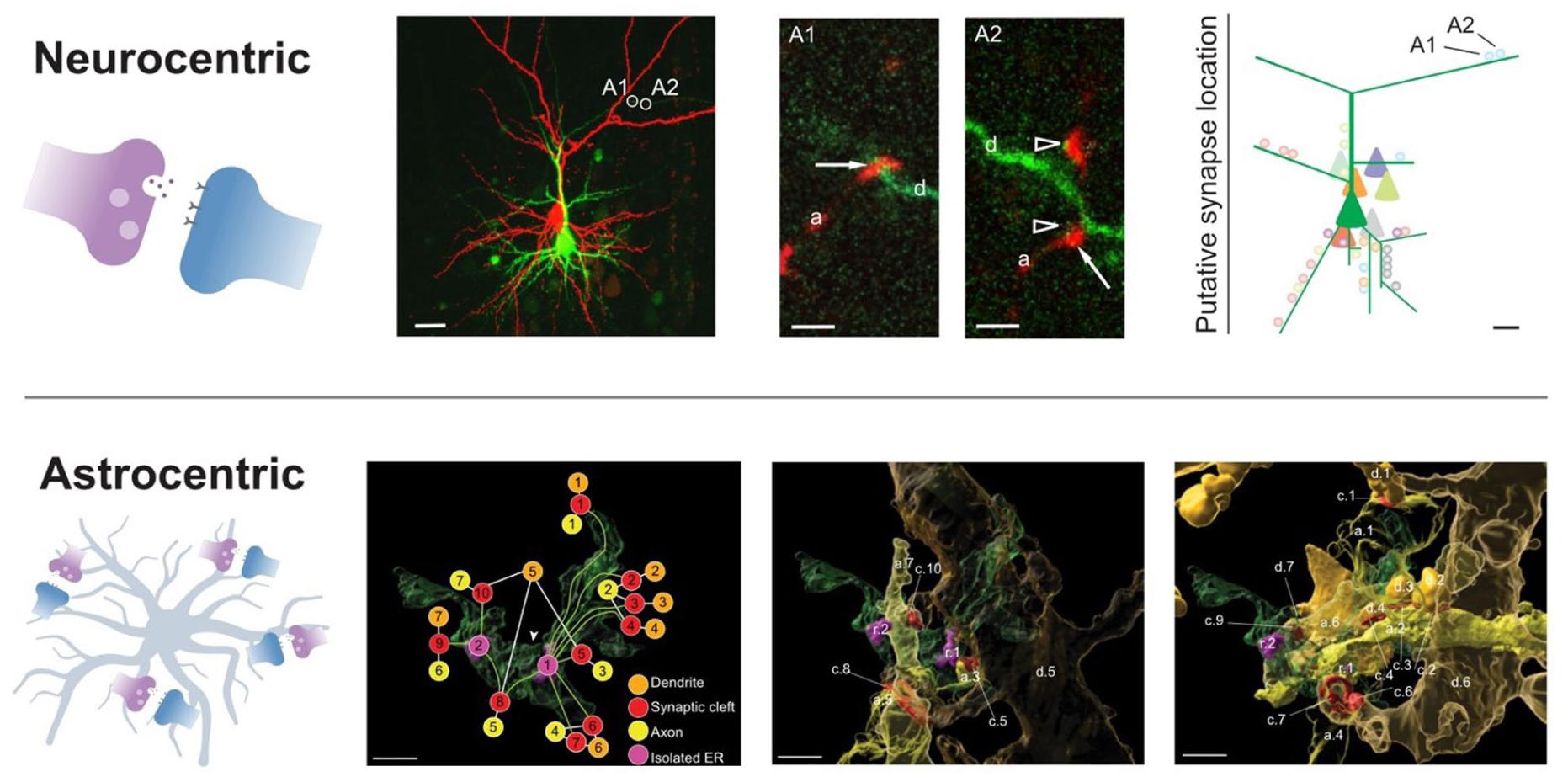
Neurocentric and astrocentric views of synaptic organization. Top: Putative synaptic contacts between 2 pyramidal neurons were mapped by labeling the presynaptic cell with biocytin (red) and the postsynaptic cell with Alexa Fluor 488 (green). Insets highlight presynaptic axons contacting postsynaptic dendritic spines, and reconstructed contact locations were combined across several pairs to generate a spatial map of synapse positions along the postsynaptic dendritic tree. A = synaptic contact; a = axon; d = dendrite. Arrow = synaptic bouton; arrowheads = dendritic spine. Scale bars: 30, 2, and 10 µm. Adapted with permission from Zilberter et al (2009). Bottom: Three-dimensional reconstructions show astrocyte leaflets (transparent green) forming spatially organized microdomains that enwrap multiple excitatory synapses, including dendritic segments (orange), axons (yellow), and synaptic clefts (red). Within a single leaflet domain, distinct connectivity motifs are evident, including convergence of multiple axons onto 1 dendrite and divergence of 1 axon onto multiple dendrites. ER = endoplasmic reticulum. Arrowhead = gap junction. Scale bars: 0.3 µm. Adapted with permission from Benoit et al (2025).
Even if 1- and 2-factor learning rules are sufficient for synaptic encoding and stabilization, incorporating astrocytes into models of plasticity may offer additional computational advantages. Plasticity may therefore reflect not only synapse-specific modifications but also coordinated regulation across synaptic ensembles organized within shared astrocytic territories.
Astrocytes Tune, Gate, and Regulate
Neuromodulators such as norepinephrine signal reinforcement contingencies and behavioral state (Breton-Provencher et al 2022). Astrocytes are increasingly recognized as intermediaries in neuromodulation across circuits, contexts, and species (Chen et al 2025; Guttenplan et al 2025; Lefton et al 2025). For example, norepinephrine signals to astrocyte Gq protein–coupled receptors to induce adenosine triphosphate/adenosine release, which binds to presynaptic adenosine receptors and modulates synaptic transmission (Chen et al 2025; Lefton et al 2025). In sensory systems, astrocyte–neuron feedback loops can contribute to adaptation, positioning astrocytes as active components in state-dependent circuit regulation (Lines et al 2020).
Through these interactions, astrocytes can tune the dynamic range of sensory information processing by shaping cortical network activity, in part through modulation of inhibitory transmission (Miguel-Quesada et al 2023). Beyond synaptic modulation, astrocytes support brain rhythms—the synchronization of neuronal activity associated with behavioral states that are important for cognitive function and information processing (Lee et al 2014; Sardinha et al 2017; Andrade-Talavera et al 2023a).
Astrocytes may thus act as context-dependent gates that bias which synaptic pathways influence circuit output. This view is reminiscent of mixture-of-experts architectures in machine learning, in which a gating network dynamically weights the contribution of specialized subnetworks (Jacobs et al 1991; Jordan and Jacobs 1994). This suggests the possibility that astrocytes play a similar gating role in biological circuits.
Astrocytes have been proposed to act as activity gates, ensuring that only sufficiently strong or salient activity patterns are propagated through local circuits (Lines et al 2024). More broadly, astrocytes may gate modulatory signals and thereby act directly on circuit function (Figure 4A). Astrocytes may thus impose context-dependent constraints on neural computations (Murphy-Royal et al 2023).

Computational roles of astrocytes in plasticity and learning. (A) In the contextual guidance model, astrocyte signaling can reconfigure neuronal activity in a context-dependent manner. Adapted with permission from Murphy-Royal et al (2023). (B) Astrocytes may act as thresholding integrators that accumulate activity-dependent signals across repeated pre- and postsynaptic pairings. Once astrocyte activation reaches a sufficient level, gliotransmitter release enables the induction of timing-dependent long-term depression. Adapted with permission from Min et al (2012). (C) In a model of neuron–astrocyte network architecture that supports encoding, storage, and retrieval of information, an astrocyte network can increase the computational capacity of brain circuits. Adapted with permission from Tsybina et al (2022). (D) Illustration of credit assignment in a simple multilayer network. Neurons in the input layer can be strongly active (dark green), yet reducing output error may require changes in a hidden neuron such as X, whose contribution depends on how downstream units transform its activity. A core idea is that biologically plausible learning separates fast feedforward signaling from slower error feedback signals via an additional compartment such as dendrites (Guerguiev et al 2017). Astrocytes could provide an analogous compartment for integrating delayed teaching signals that gate plasticity, positioning astrocytes as potential contributors to credit assignment in biological circuits. Adapted with permission from Richards and Lillicrap (2019).
Together, these findings suggest that astrocytes can translate neuromodulatory state signals into local circuit tuning and gating. In emotional states such as anxiety, astrocyte activity can encode behavioral state variables across contexts, providing a stable and decodable representation of internal state (Ghenissa et al 2026). By doing so, they may help determine when and where plasticity is permitted, rather than specifying the exact synapses that should change. Beyond circuit tuning, astrocytes may function as slower integrators that determine whether accumulated activity reaches the threshold required for long-term synaptic change.
Astrocytes as Arbiters of Plasticity
Because astrocytes respond to neuronal activity with calcium events at slower timescales than neurons, they could act as spatiotemporal integrators that arbitrate when plasticity should occur. A long-standing idea is that astrocytes detect the collective activity of nearby synapses and integrate these signals to tune neuronal excitability across local networks. Astrocytic signaling can thereby provide a third factor that reflects the level of local neuronal activation and permits synaptic change only when activity is sufficiently strong or sustained.
It has been proposed that during the induction of tLTD (Figure 4B), astrocytes may act as thresholding integrators by sensing the gradual buildup of eCBs generated by repeated pre- and postsynaptic pairings (Min et al 2012). In this view, as eCB signaling accumulates, astrocytic calcium activity ramps up until it crosses a threshold that permits tLTD induction (Min et al 2012). Additionally, astrocytes are able to decode neuronal activity, resulting in distinct calcium elevation patterns and specific gliotransmitter responses (Covelo and Araque 2018; Cahill et al 2024). It remains an open question, however, whether astrocyte responses are typically all-or-nothing or relatively graded.
Astrocytes are sensitive to internal state. Astrocytic calcium activity is relatively weak and long-lasting during sleep but becomes larger, more widespread, and brief during arousal, consistent with a role in state-dependent regulation of circuit plasticity (Wang F et al 2023). Astrocytes can also influence dendritic computations by regulating extracellular
Taken together, astrocyte signaling is well suited to provide a third factor that integrates local activity and internal state, thereby regulating long-term plasticity induction. This form of arbitration is naturally aligned with the slower dynamics of astrocyte calcium and its ability to influence synaptic and dendritic nonlinearities. Astrocytes may therefore define the permissive landscape within which synaptic plasticity occurs. Building on this view, recent modeling work suggests that astrocytes may contribute an additional layer of information processing capable of stabilizing and retrieving neuronal activity patterns.
Astrocytes as an Additional Computational Substrate
Astrocytes have been proposed to provide an additional computational substrate capable of storing and transforming neuronal information over longer timescales (Figure 4C). For instance, Zucker (2012) argued that glia function as information-processing elements, potentially expanding the computational repertoire of neural circuits. In such a framework, activity-dependent changes in astrocyte calcium could encode aspects of recent synaptic and network history, enabling information to be retrieved or reinstated when a relevant stimulus arrives (Mu et al 2019; Sánchez Romero and Navarrete 2026). Consistent with this idea, modeling work has suggested that incorporating astrocyte dynamics can expand the repertoire of network behavior, supporting pattern storage and recall beyond what a purely neuronal architecture can achieve (Tsybina et al 2022).
In a compartmental astrocyte model, the neuronal firing rate was found to determine the rate of local calcium transients in fine astrocyte processes as well as in the soma, whereas correlations among neuronal inputs shaped the occurrence of more global calcium events (Cresswell-Clay et al 2018). This separation of local and global calcium signaling provides a natural substrate for multiscale integration, in which astrocytes simultaneously track local synaptic drive and broader network structure.
Kozachkov et al (2025) proposed that neuron–astrocyte interactions could provide an additional substrate for memory storage. In their theoretical framework, memory patterns can be encoded within astrocytic process connectivity and calcium-dependent signaling, with neuron–synapse–astrocyte dynamics supporting retrieval. Notably, modeling astrocyte processes as computational units increased the predicted storage capacity and scaling of the network.
Together, these models suggest that astrocytes introduce computational degrees of freedom beyond neuronal spiking, enabling information storage and retrieval through slower state variables and compartmental signaling. While some of these hypotheses have received experimental support (Sánchez Romero and Navarrete 2026), their roles in memory encoding and recall remain incompletely understood. Astrocyte dynamics may therefore shape how learning is distributed across space and time in biological circuits, offering an alternative perspective on the synaptic credit assignment problem.
Astrocytes and the Dissolution of the Credit Assignment Problem
Astrocytes are increasingly recognized as active participants in information processing, with roles that extend beyond metabolic support into synaptic regulation, circuit tuning, and plasticity control. These properties have prompted the idea that astrocytes could contribute not only to biological learning but also to improved learning mechanisms in artificial systems that incorporate neuron–glia interactions (Porto-Pazos et al 2011). If astrocytes shape when and where synapses are permitted to change, they may motivate a reframing of one of the central theoretical tensions between modern machine learning and neurobiology, namely the credit assignment problem. This view aligns with recent astrocyte frameworks (Murphy-Royal et al 2023) while emphasizing permissive control of synaptic plasticity. Here we consider whether permissive control of plasticity by astrocytes offers an alternative perspective on the credit assignment problem in biological circuits.
In this view, astrocytes may help resolve the credit assignment problem by regulating when and where synaptic plasticity is permitted. This may seem to raise the question of how credit is assigned to astrocytes themselves. However, rather than requiring precise, local credit assignment, astrocytes may instead integrate slower, population-level signals that reflect behavioral context.
In artificial neural networks, learning requires assigning credit or blame to individual synaptic weights based on their contribution to overall network performance. In practice, this is most commonly achieved by using error backpropagation, in which output errors are propagated backward through successive layers to gradually adjust synaptic strengths (Box 2, Figure 4D). However, this mechanism relies on assumptions that appear biologically unrealistic; for example, synapses do not propagate information backward (Crick 1989).
Furthermore, the need for explicit credit assignment in deep networks does not necessarily imply that biological circuits implement an equivalent mechanism. One key difference is that forward computation in many artificial neural networks is dense, with most units participating continuously, whereas biological computation is typically sparse, with many neurons remaining silent at any given moment (Barth and Poulet 2012). As a result, information processing in the brain is effectively carried by a shifting subset of active neurons, supporting flexible computation while helping constrain energy consumption, so the functional network engaged in a given computation continuously reconfigures itself. This dynamic restructuring may reduce the need for synapse-level precision in credit assignment, instead favoring learning rules that operate over local synaptic clusters under permissive control (Govindarajan et al 2011; Larkum 2013; Richards and Lillicrap 2019; Falcón-Moya et al 2020).
This shifts the focus from how plasticity is implemented to where and when plasticity is permitted. Astrocytes are well positioned to provide a substrate for such permissive, cluster-level control. Artificial neuron–glia networks, where astrocyte-like units provide context-dependent modulation of synaptic activity across local networks, have shown improved performance in computational models (Alvarellos-Gonzalez et al 2012) and could therefore help address credit assignment. Consistent with this view, astrocytes can gate plasticity and influence metaplasticity, and synaptic clustering within astrocyte domains (Benoit et al 2025) supports the idea that they can territorially regulate groups of synapses, potentially through heterosynaptic mechanisms. Recent ultrastructural reconstructions further suggest that astrocytes partition local synaptic space into astrocyte-defined synaptic clusters (Salmon et al 2023). This is consistent with microdomain-level regulation of circuit remodeling.
Another difficulty for credit assignment is that synapses must somehow separate learning-relevant teaching signals from ongoing background activity when both arrive through overlapping pathways. Richards and Lillicrap (2019) proposed that dendritic compartmentalization and computation could help address this problem (Figure 4D). It is tempting to speculate that astrocytes provide a parallel compartment for integrating delayed contextual signals that permissively gate plasticity (Perea et al 2009).
However, these views on the solution to the credit assignment problem in the brain may reflect an incomplete framing. Credit assignment presumes a global output error signal routed to specific synapses, yet no biologically established mechanism for precise backward transport of gradients exists (Crick 1989), although approximate schemes that relax strict weight transport have been proposed in artificial systems (Lillicrap et al 2016). By contrast, known plasticity rules such as Hebbian and STDP are local and state dependent rather than global in the machine learning sense. Multiple circuit configurations may implement similar functions, reflecting degeneracy in neural systems, whereby distinct circuit configurations can give rise to similar outputs (Edelman and Gally 2001; Marder and Taylor 2011), suggesting that identifying a single causal synapse may not be necessary for effective learning.
The brain’s solution to credit assignment may instead rely on fine-grained modularity shaped by local astrocyte leaflets. This perspective reframes the problem in terms of permissive control rather than synaptic blame. At the same time, it does not preclude the possibility that such processes still implement gradient-like optimization at a systems level, as has been argued for biological plasticity more broadly (Richards and Kording 2023).
At its most basic level, learning in biological neural systems requires local activity-dependent synaptic plasticity. Hebbian learning and STDP provide rules by which synaptic efficacy is modified according to the temporal coincidence of pre- and postsynaptic activity (Hebb 1949; Bi and Poo 1998; Sjöström et al 2001). More recently, BTSP links synaptic change to postsynaptic plateau potentials over seconds, providing a local rule better matched to behavioral learning timescales (Bittner et al 2017; Milstein et al 2021). Such 2-factor rules are inherently local, relying only on signals available at the synapse or within the postsynaptic dendritic compartment. However, purely local correlation-based learning is fundamentally ambiguous: it cannot, on its own, distinguish behaviorally meaningful correlations from spurious co-activation caused by noise, common input, or ongoing network dynamics.
This ambiguity may be resolved by neuromodulatory gating of plasticity (Pawlak et al 2010), which links local synaptic eligibility to global behavioral context. Neuromodulatory systems broadcast low-dimensional signals related to reward, salience, arousal, or uncertainty (Aston-Jones and Cohen 2005; Glimcher 2011; Dayan 2012; Schultz et al 1997). These signals do not appear to specify which synapses should change. Rather, they act permissively, stabilizing or enabling plasticity at recently active synapses. This way, neuromodulation may convert Hebbian plasticity into 3-factor learning that is sensitive to behavioral relevance without requiring synapse-specific credit assignment.
A critical intermediate step between local synaptic activity and delayed neuromodulatory signals is provided by synaptic eligibility traces. Eligibility traces are transient synaptic states set by pre- and postsynaptic co-activity that do not themselves cause lasting plasticity but render a synapse eligible for modification if a neuromodulatory signal arrives within a limited temporal window. This framework underlies 3-factor learning rules in which synaptic change depends on the conjunction of local activity, a modulatory signal, and a temporally extended synaptic trace (Izhikevich 2007; Gerstner et al 2018). Experimental evidence for eligibility traces has, for example, been reported in cortex and striatum, where monoamines can selectively stabilize synapses that were active seconds earlier (Yagishita et al 2014; He et al 2015). Eligibility traces therefore provide a biologically plausible solution to the temporal credit problem without requiring explicit backpropagation of error signals.
A further constraint is required to prevent widespread or runaway plasticity during neuromodulatory events. Astrocytes provide this constraint by enforcing local homeostasis through glutamate uptake, potassium buffering, and metabolic regulation (Rothstein et al 1996; Anderson and Swanson 2000; Oliet et al 2001). By regulating glutamate uptake through high-affinity transporters, astrocytes control extracellular glutamate levels and thereby set the conditions under which NMDA receptors are engaged (Rose et al 2018). Transient activity-dependent slowing of astrocytic glutamate clearance can prolong extracellular glutamate signaling and enhance NMDA receptor activation (Armbruster et al 2016), thereby modulating downstream plasticity. In parallel,
Finally, effective learning in biological circuits may rely on redundancy and degeneracy, rather than precise synapse-level credit assignment. Multiple distinct circuit configurations can support similar functional outputs, and learning need only bias a subset of co-active synapses within a redundant ensemble to achieve behavioral change (Edelman and Gally 2001; Marder and Taylor 2011). In this view, plasticity may therefore operate at the level of ensembles and synaptic clusters within microdomains, not individual causal synapses. Neuromodulatory broadcast, eligibility traces, and astrocytic gating may exploit this redundancy, stabilizing patterns of activity across groups of synapses while maintaining robustness to noise, partial failure, and ongoing circuit reconfiguration.
From this perspective, the brain may dissolve the credit assignment problem not by assigning credit to individual synapses but by regulating when and where plasticity is permitted, with astrocytes helping define the local permissive state for synaptic change.
Conclusions
In this review, we propose that the brain does not solve the credit assignment problem in the same way that it is framed in machine learning. Instead, astrocytes may help reframe it by selectively permitting plasticity in local circuit microdomains, reducing the need for synapse-level credit assignment. In this sense, synaptic change may be governed less by neuron–neuron interactions alone and more by a 3-body dynamic in which astrocytes help determine when and where learning is allowed to occur.
Importantly, these perspectives need not be mutually exclusive. Biological learning processes may still approximate gradient-based optimization, even if implemented through local and distributed mechanisms rather than explicit error backpropagation (Richards and Kording 2023). However, forms of one-shot learning, in which lasting memory traces can arise from a single or very brief experience, suggest that at least some biological learning operates outside a regime of gradual error-driven updates and may instead depend on permissive gating of plasticity in space and time (Piette et al 2020).
More broadly, this view highlights the need for caution when comparing engineered and biological learning systems. As Dijkstra (1984) remarked, “the question of whether a computer can think is no more interesting than the question of whether a submarine can swim.” In a similar spirit, Richard Feynman noted that planes do not flap their wings like birds, reminding us that engineered systems need not replicate biological mechanisms to achieve functional success. Accordingly, although deep learning relies heavily on explicit backward propagation of error, biological circuits may instead depend on spatially and temporally structured plasticity that is selectively permitted rather than explicitly assigned.
Recent evidence indicates that dendritic compartments can encode vectorized error-related signals necessary for learning (Francioni et al 2026), supporting the idea that the cortex can implement structured credit-related signaling within single neurons. Rather than negating a role for astrocytes, these findings suggest that instructive dendritic signals and astrocyte-mediated permissive control may operate at distinct but interacting levels, jointly shaping where and when plasticity is expressed.
Viewed in this broader context, insights from machine learning remain valuable, even if the underlying implementations differ. Machine learning has become a powerful tool for revealing structure in complex brain data. For instance, Azabou et al (2025) showed that large-scale calcium-imaging models can generalize across brain regions and neuronal cell types, extracting latent structure without explicit labels. As astrocyte calcium datasets scale, similar approaches may help reveal what astrocytes encode across local and global brain states. Such approaches will help test theoretical frameworks such as the one proposed here. Biological and engineered systems need not share implementation to yield mutually informative insights. For example, machine learning analyses of large-scale recordings have revealed structure in cortical population activity that informs circuit theory (Cowley et al 2026).
Beyond their contributions to plasticity, astrocytes contribute to many core functions across brain physiology. They engage in reciprocal signaling with microglia that shapes astrocyte morphology and synapse remodeling (Faust et al 2025; Gu et al 2025; Li et al 2025). Astrocytes also associate with oligodendrocytes, blood vessels, and the extracellular environment to exert coordinated actions across neural tissue (Kwon et al 2025).
Here, we deliberately adopt an astrocentric perspective on plasticity, focusing on the potential roles of astrocytic compartments in determining plasticity. However, plasticity is governed by multiple interacting cellular compartments. Interneurons, for example, exert powerful inhibitory control over plasticity (McFarlan et al 2023). Likewise, dendrites gate and modulate plasticity through a reciprocal interplay with synaptic signaling that shapes plasticity rules (Sjöström et al 2008). Specifically, pyramidal dendrites can act as associative compartments where coincident inputs trigger calcium spikes and burst firing that support memory formation (Larkum 2013). Moreover, there are forms of plasticity that do not involve astrocytes at all (Rodríguez-Moreno et al 2013). Rather than competing explanations, these processes likely operate in concert so that plasticity emerges from coordinated interactions across distinct neuronal and glial compartments.
Astrocyte dysfunction has been implicated in disorders with disrupted plasticity and memory, including Alzheimer disease, where synapse loss strongly tracks cognitive decline and astrocytes may actively contribute to synaptic vulnerability (Hulshof et al 2022). As a result, astrocytes are increasingly being explored as therapeutic targets for restoring circuit stability and cognitive function (Chierzi et al 2023; Alfonso-Triguero and Arranz 2026). Understanding how astrocytes regulate where and when plasticity can occur may therefore prove important not only for theories of learning but also for strategies aimed at repairing it. Astrocytes thus emerge as a plausible organizing principle for learning in biological circuits, reframing credit assignment as a problem of permissive control rather than synaptic blame.
Footnotes
Acknowledgements
We thank Alanna Watt, Aparna Suvrathan, Pouya Bashivan, Anmar Khadra, Paul Masset, Kaleem Siddiqi, Albert Hiu Ka Fok, Tabish Syed, Haley Renault, and Sjöström laboratory members for help and useful discussions.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: A.W. received awards from Fonds de recherche du Québec – Santé (FRQS; 335452), Quebec Bio-Imaging Network, Research Institue of the McGill University Health Centre (RI-MUHC), and Healthy Brains Healthy Lives. C.G. received awards from Natural Sciences and Engineering Research Council of Canada (NSERC) Undergraduate Student Research Awards (USRA) (2023-584320), FRQ – Nature et Technologie USRA Supplement (342221), RI-MUHC, Canadian Institutes of Health Research (CIHR) Canada Graduate Research Scholarship-Master’s program (524050), and FRQS (BF1 346922). P.J.S. was funded by the Montreal General Hospital Foundation, Canada Foundation for Innovation Leaders Opportunity Fund (28331), CIHR (Project Grants 156223, 191969, 191997), FRQS Chercheurs-boursiers (254033), and NSERC Discovery Grant/Discovery Accelerator Supplement (2024-06712, 2017-04730, 2017-507818). P.J.S. is a recipient of a Donald S. Wells Distinguished Scientist Award.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.