
Abstract
Objectives:
Lipopolysaccharide (LPS) from oral Gram-negative bacteria induces inflammatory responses involving the chemokine CXCL1. Although CXCL1 is known to mediate inflammation, its role in disease processes remains incompletely understood. This study aimed to investigate the biological function of CXCL1, evaluate shRNAs as inhibitors, and determine whether CXCL1 suppression is associated with reduced LPS-induced cytokine production.
Materials and Methods:
Human CXCL1 cDNA was cloned into recombinant plasmid DNA and transfected into THP-1 monocyte-like cells. CXCL1-regulated cytokines, including tumor necrosis factor alpha (TNF-α) and IL-1β, were quantified by enzyme-linked immunosorbent assay (ELISA) following treatment with pharmacologic and molecular inhibitors. Multiple CXCL1-targeting shRNAs were designed, synthesized, and tested. The specificity of CXCL1 shRNA-mediated inhibition of LPS-induced proteins was analyzed using ELISA and a hybrid method analysis.
Results:
Treatment with a Janus kinase inhibitor or CXCL1 shRNA#2 significantly suppressed CXCL1 expression, resulting in reduced production of TNF-α and other pro-inflammatory cytokines, suggesting a regulatory role for CXCL1. CXCL1 shRNA#2 also markedly inhibited LPS-induced chemokines, kinases, transcription factors, and apoptotic genes, demonstrating broad anti-inflammatory effects.
Conclusions:
CXCL1 appears to play a regulatory role in LPS-dependent inflammatory pathways, and its suppression by shRNA or pharmacologic inhibitors effectively reduces cytokine production. These findings provide insight into CXCL1-associated mechanisms while highlighting the need for further mechanistic studies to elucidate the precise signaling pathways involved and may inform the development of potential therapeutic strategies for inflammatory diseases.
Introduction
High levels of chemokine ligand-1 (CXCL1) protein have been detected in sites of various diseases (Kuo et al., 2012; Sahingur and Yeudall, 2015; Verbeke et al., 2011; Wen and Wang, 2022). CXCL1 is also implicated in lipopolysaccharide (LPS)-induced inflammatory responses (Lu et al., 2022). Inflammation is a critical defense mechanism against harmful stimuli or pathogens; however, excessive inflammatory responses can lead to immune system disorders (Tang et al., 2011). LPS, a component of Gram-negative bacteria, induces the production of cytokines and chemokines such as TNF-α and CXCL1 in macrophages, thereby initiating an inflammatory response (Elgreu et al., 2022; Naruishi and Nagata, 2018; Rath-Deschner et al., 2020; Tang et al., 2018).
CXCL1 regulates the recruitment and migration of neutrophils during infection (Liu et al., 2018; Liu et al., 2022). Its activity can be inhibited using receptor antagonists C-X-C motif chemokine receptor 2 (CXCR2), toll-like receptor 4 antagonists, nuclear factor kappa B (NF-κB) inhibitors, and JNK inhibitors (Nair et al., 2012); however, the specificity of these inhibitors remains unclear (Huang et al., 2021; Tam et al., 2023). However, it remains unclear whether CXCL1 functions solely as a downstream marker of inflammation or plays a direct regulatory role in cytokine production. Recently, RNA interference (RNAi) has emerged as a powerful approach to overcome nonspecific inhibition by selectively knocking down gene expression (Chen et al., 2023). RNAi employs small interfering RNAs that, through the RNA-induced silencing complex, cleave target messenger RNA (mRNA) in a sequence-specific manner, thereby reducing protein expression (Tanner et al., 2022). Vector-based short hairpin RNAs (shRNAs) provide an effective means of implementing RNAi (Arruda et al., 2022) and have demonstrated therapeutic potential in treating various diseases, including cancer (Barresi et al., 2022).
In this study, we aimed to use shRNAs to inhibit CXCL1 and determine whether its suppression is associated with reduced LPS-induced TNF-α production. We cloned recombinant plasmid DNA containing human CXCL1 cDNA to analyze its biological function. CXCL1 overexpression induced TNF-α and IL-1β production, and our data indicate that Janus kinase (JAK) is involved in this pathway. Among 10 different CXCL1-targeting shRNAs, shRNA #2 was identified as the most effective inhibitor of LPS-induced cytokine production.
Materials and Methods
Cell lines
Human THP-1 monocyte-like cells (Cat# TIB-202™, ATCC®) were cultured in RPMI 1640 medium (Cat#: 11875-093, Life Technologies, NY) under standard conditions.
Protein array
Cytokine arrays were used to detect multiple analytes in cell culture supernatants. THP-1 cells (1 × 10^5 cells/well) were seeded in 6-well plates and either left untreated as a control or treated with 100 ng/mL LPS for 24 h. Supernatants (100 µL) from each well were applied to the cytokine arrays. Array images were acquired and analyzed using the LI-COR Odyssey Infrared Imaging System. Human cytokine array kits were obtained from R&D Systems (Cat# DGR00B, MN, USA).
CXCL1 cDNA cloning
CXCL1 cDNA, provided by Harvard University (Clone ID: HsCD00326770), was amplified by PCR using the primers 5′-ATGTCGGGGGACACCTGCCTG-3′ and 5′-TCAGTTGGATTTGTCACTGTT-3′. The amplified fragment was cloned into the pcDNA3 plasmid to generate recombinant plasmid pcCL7. Cloning was confirmed by DNA sequencing.
Inhibitors
The following inhibitors were purchased from Calbiochem and prepared according to the manufacturer’s instructions: BAY11-7082 (NF-κB inhibitor, Cat#: 481406), LY294002 (phosphoinositide 3-kinase (PI3K)/protein kinase B (Akt) inhibitor, Cat#: 440202), SB202190 (p38 mitogen-activated protein kinase inhibitor, Cat#: 559389), AG490 (JAK inhibitor, Cat#: 658401), and U0126 (mitogen-activated protein kinase kinase (MEK) inhibitor, Cat#: 662005).
CXCL1 shRNA synthesis
Ten 29-mer CXCL1-targeting shRNAs were designed based on the human CXCL1 mRNA sequence (Origene, MD). After screening, shRNA #2 (29-mer, Cat#: TF315486, Origene, MD) was selected as the most effective. The sequences used are as follows:
shRNA #2: 5′-GGCGGAAAGCTTGCCTCAATCCTGCATCC-3′. Control shRNA: 5′-GCACTACCAGAGCTAACTCAGATAGTACT-3′.
ELISA analysis
Supernatants from treated THP-1 cells were analyzed by ELISA for CXCL1 (ab190805, Abcam), caspase 3 (ab220655, Abcam), TNF-α (ab181421, Abcam), and IL-1β (A35574, Invitrogen). Absorbance was measured using a microplate reader (Model 680, Bio-Rad, Portland, ME), and data were graphed accordingly.
Protein array analysis
Protein array analysis was performed as previously described (Edabashi et al., 2023).
Statistical analysis
All experiments were performed in triplicate. Data were analyzed using the SAS software package. Single mean comparisons were conducted by analysis of variance followed by Student’s t-test. P values of <0.05 were considered statistically significant.
Results
TNF-α and IL-1β are known to be induced in LPS-treated THP-1 monocyte-like cells. We investigated whether other cytokines or chemokines are also induced by LPS using a protein array. As shown in Fig. 1, one chemokine, CXCL1, was significantly upregulated by LPS, producing approximately three times more protein than in untreated cells.
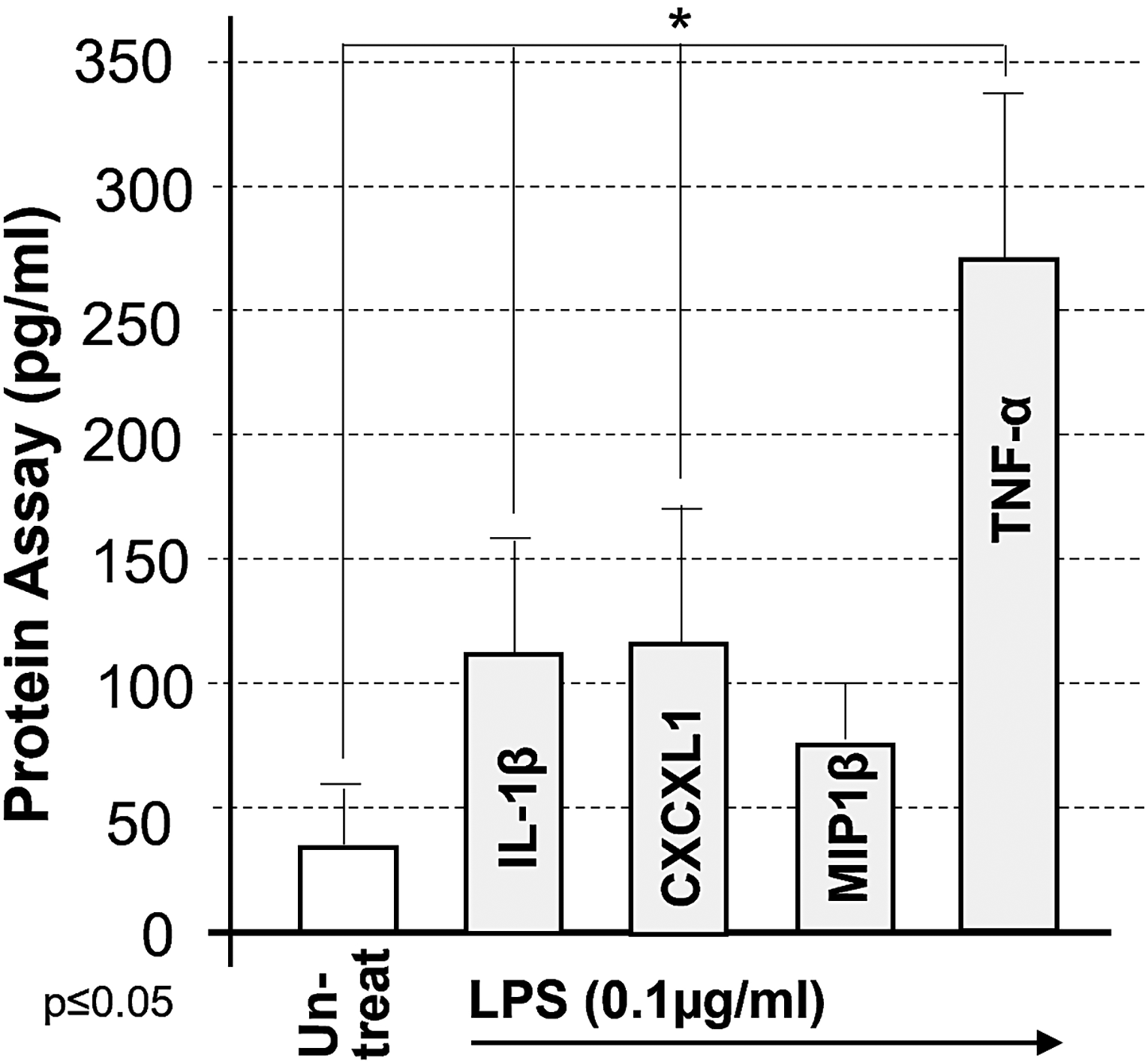
Protein array of LPS-treated cytokine/chemokine. 1 × 105 THP-1 cells were untreated as the control and treated with 100 ng/mL Escherichia coli LPS for 24 h. Cell culture supernatants were collected and analyzed by protein array. Induction of cytokines/chemokines as the test group was compared with the control and graphed. LPS, lipopolysaccharide.
To study the biological function of CXCL1, a recombinant plasmid (pcCL7) containing human CXCL1 cDNA (324 bp) was constructed, with pcDNA3 serving as the backbone plasmid. The plasmid’s physical map, with a total length of 5,724 bp, is shown in Fig. 2.
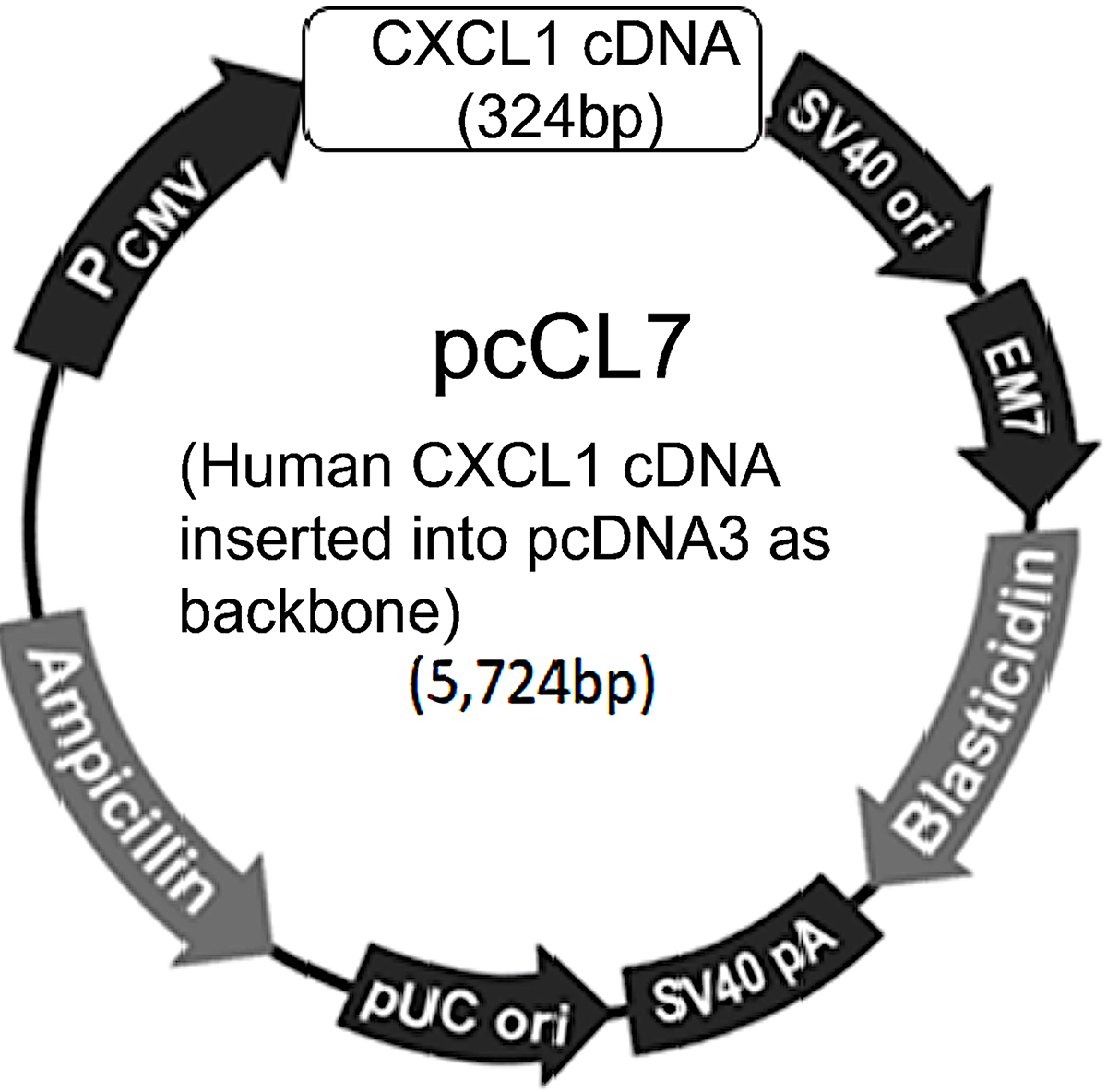
The physical map of pcCL7 recombinant plasmid DNA.
Since LPS induces both TNF-α/IL-1β and CXCL1, we examined whether additional factors regulate TNF-α and IL-1β production. As shown in Fig. 3, overexpression of pcCL7 alone (inducing CXCL1 protein) significantly increased TNF-α levels (Fig. 3, No. 2). Cotreatment with pcCL7 and a JAK inhibitor reduced CXCL1-induced TNF-α production by approximately 22% compared to the positive control (Fig. 3, No. 6). This inhibitory effect was not observed with other inhibitors (Fig. 3, Nos. 3–5, 7, and 8), suggesting that JAK may be involved in CXCL1-regulated TNF-α and IL-1β production.
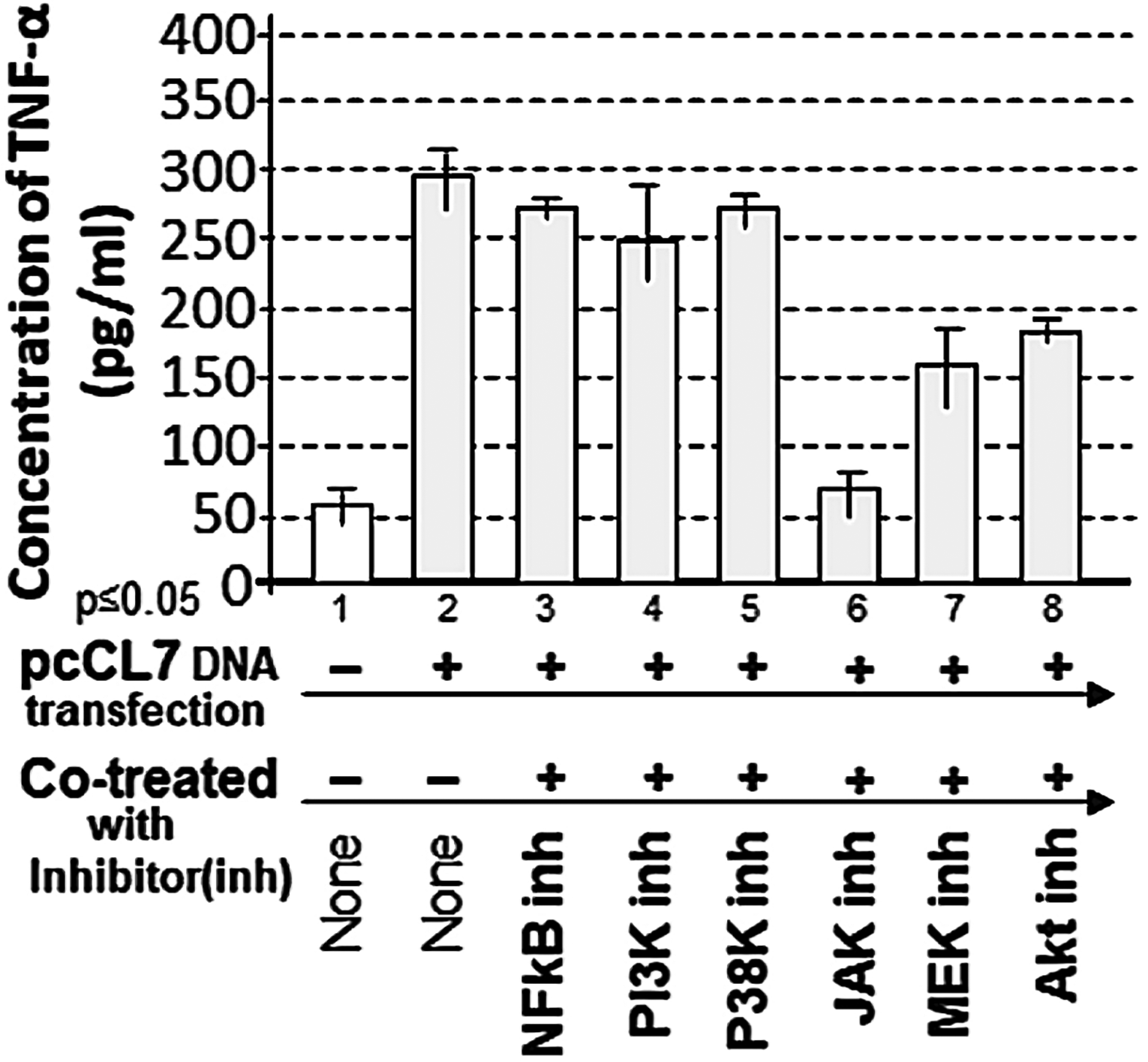
Analysis of the effects of inhibitors on pcCL7 overexpression (CXCL1)-mediated TNF-α production. 1 × 105 THP-1 cells were either untreated as the negative control, transfected with 1 µg/mL pcCL7 alone as the positive control, or cotreated with 1 µg/mL pcCL7 plus inhibitor (5 µM BAY11-7082 as NFkB inhibitor, 5 µM LY294002 as PI3K inhibitor, 10 µM SB202190 as p38K inhibitor, 10 µM AG490 as JAK inhibitor, 5 µM U0126 as MEK inhibitor, or 5 µM AZD5363 as an Akt inhibitor) as test group. The conditioned medium from each group was collected and analyzed by ELISA for the quantitative detection of TNF-a. The data were analyzed and graphed. CXCL1, chemokine ligand-1; JAK, Janus kinase.
Although JAK inhibition reduced CXCL1-induced cytokine production, it remained unclear whether JAK and CXCL1 directly regulate cytokines in response to LPS, as JAK activation can also affect other factors such as STAT (Hu et al., 2021). To achieve more specific inhibition, several CXCL1-targeting shRNAs were designed and synthesized (see Materials and Methods). These shRNAs were tested for their ability to suppress CXCL1 expression (data not shown). shRNA #2 was identified as the most effective, significantly reducing CXCL1 production in response to either CXCL1 overexpression (Fig. 4, No. 4) or LPS treatment (Fig. 4, No. 7) compared to controls (Fig. 4, Nos. 2 and 5). Importantly, the ability of shRNA#2 to suppress CXCL1 expression under both endogenous (LPS-induced) and overexpression (pcCL7-mediated) conditions supports the specificity of the CXCL1-targeting shRNA construct.
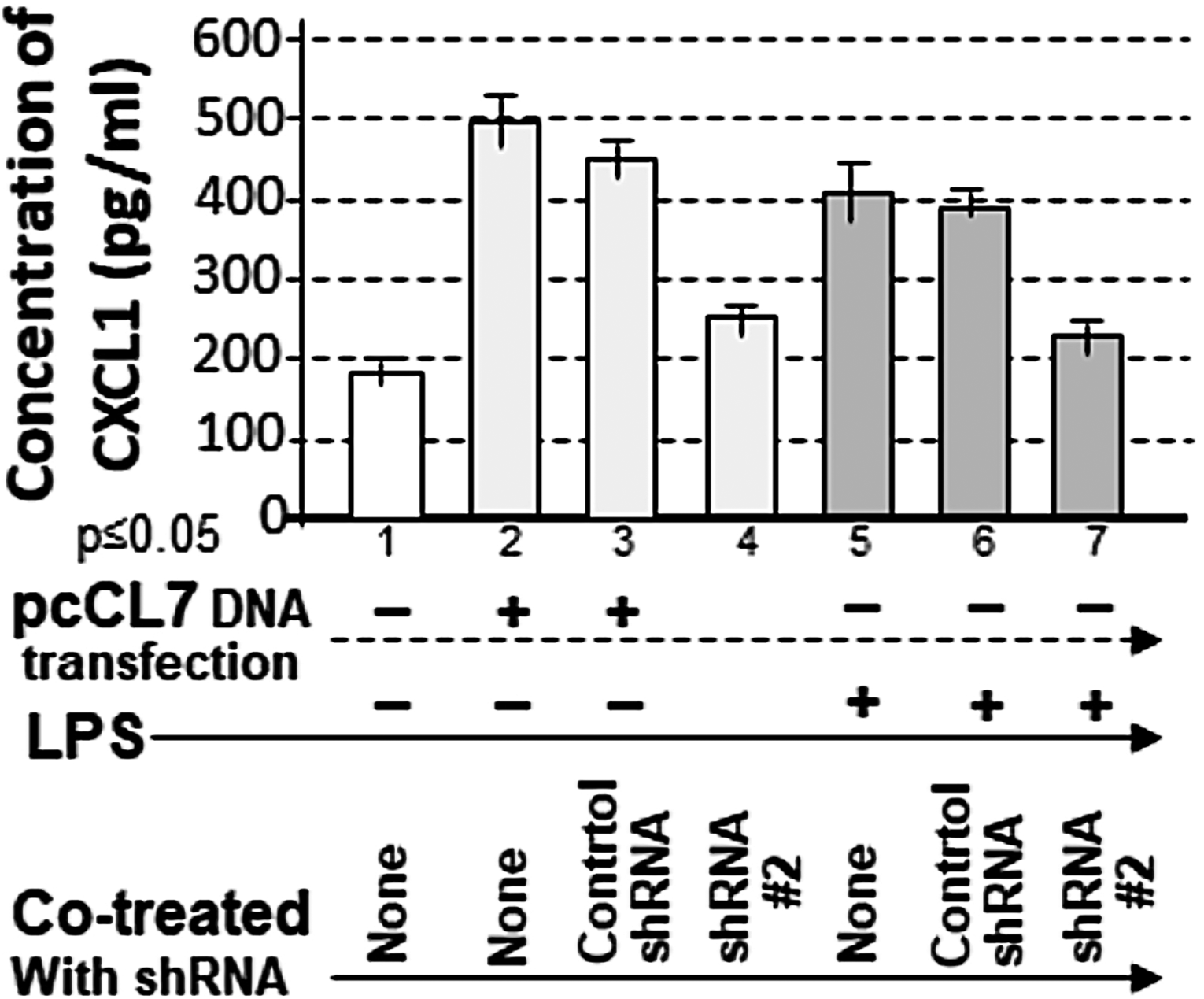
Analysis of the specificity of CXCL1 shRNA#2 for inhibition of CXCL1 gene expression in response to LPS. 1 × 105 THP-1 cells were either untreated as the negative control (No. 1) or cotreated with 1 µg/mL pcCL7 (Nos. 2–4) and 0.1 µg/mL E. coli LPS (Nos. 5–7) plus the control shRNA (Nos. 3 and 6) or shRNA#2 (Nos. 4 and 7) as the test groups. The conditioned medium from each group was collected and analyzed by ELISA for the quantitative detection of CXCL1. The data were analyzed and graphed. shRNA, short hairpin RNA.
We further examined whether shRNA #2 inhibits LPS-induced cytokine production. Cotreatment of cells with LPS and shRNA #2 significantly suppressed LPS-induced TNF-α (Fig. 5A, No. 6), IL-1β (Fig. 5B, No. 6), and caspase 3 (Fig. 5B, No. 7) compared to controls (Fig. 5A, No. 2; Fig. 5B, No. 3).
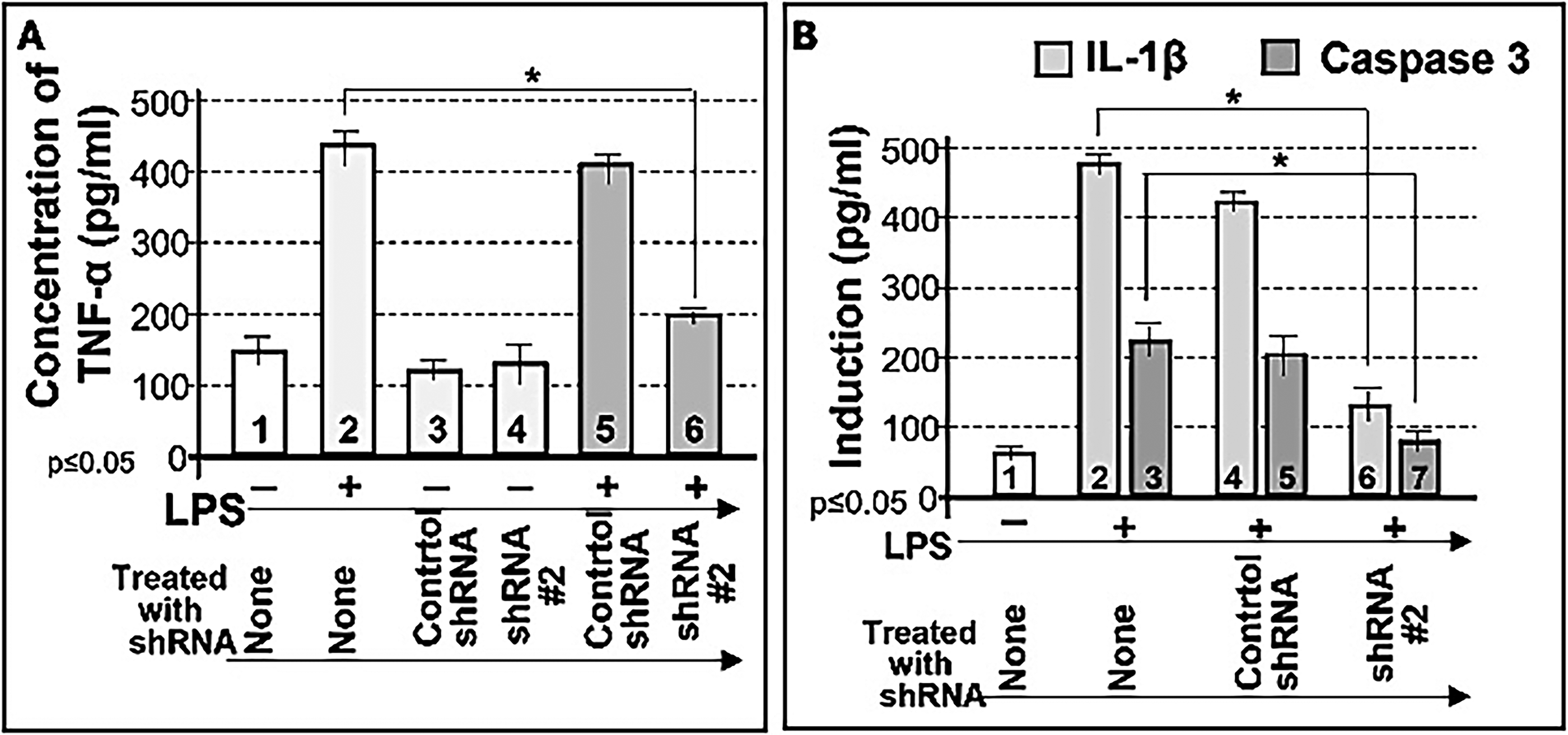
Inhibition of LPS-induced TNF-α by shRNA#2. 1 × 105 THP-1 cells were untreated (No. 1) or treated with either the control shRNA alone (No. 3) or shRNA#2 alone (No. 4) as the negative control group, treated with 0.1 µg/mL E. coli LPS as the positive control group (No. 2) or cotreated with 0.1 µg/mL E. coli LPS (Nos. 5 and 6) plus control shRNA (No. 5) or shRNA#2 (No. 6) as the test group. The conditioned medium from each group was collected and analyzed by ELISA for the quantitative detection of TNF-α
To assess whether shRNA #2 affects other LPS-responsive proteins, we examined multiple signaling molecules. Cotreatment with LPS and shRNA #2 significantly suppressed LPS-induced TNF-α (Fig. 6, No. 4), IL-1α (Fig. 6, No. 6), IL-10 (Fig. 6, No. 8), phosphorylated FAK (Fig. 6, No. 10), phosphorylated JNK (Fig. 6, No. 12), phosphorylated extracellular signal-regulated kinase 1 (ERK1) (Fig. 6, No. 14), Akt1/2/3 (Fig. 6, No. 16), caspase 3 (Fig. 7, No. 4), BAX (Fig. 7, No. 6), p53 (Fig. 7, No. 8), caspase 9 (Fig. 7, No. 10), p21 (Fig. 7, No. 12), and NF-κB (Fig. 7, No. 14), while actin served as the loading control (Figs. 6 and 7, Nos. 1 and 2). These results indicate that CXCL1 suppression is associated with reduced expression of multiple LPS-induced cytokines and signaling proteins, suggesting a potential upstream regulatory role for CXCL1. However, indirect or off-target effects cannot be excluded.
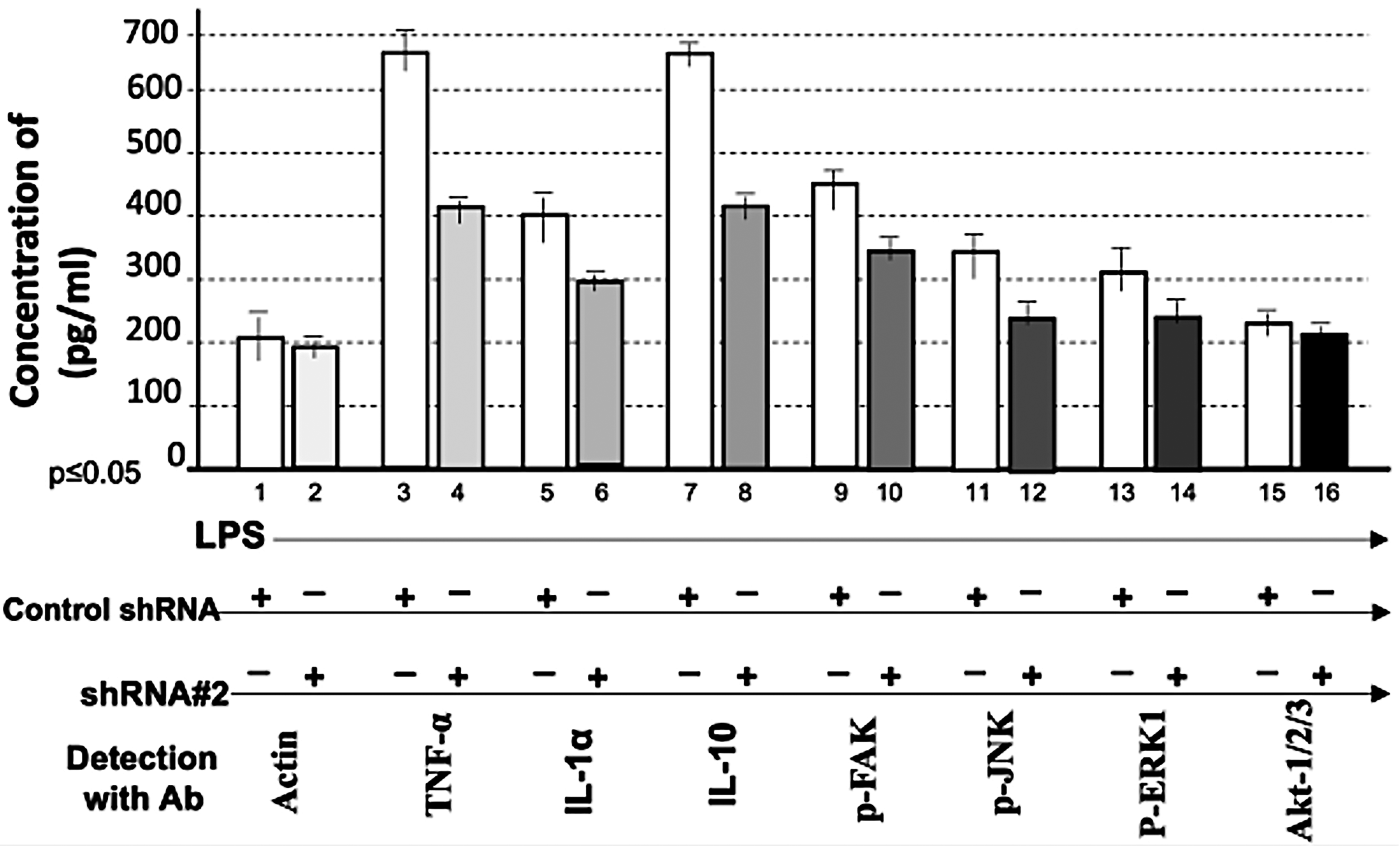
Effect of shRNA#2 on LPS-induced proteins. 1 × 105 THP-1 cells were cotreated with 0.1 µg/mL E. coli LPS plus the control shRNA or shRNA#2. The intracellular protein from each group was collected and detected by hybrid method for the quantitative detection of TNF-α (Nos. 3 and 4), IL-1α (Nos. 5 and 6), IL-10 (Nos. 7 and 8), phosphorylated focal adhesion kinase (Nos. 9 and 10), p-JNK (Nos. 11 and 12), phosphorylated extracellular signal-regulated kinase 1 (Nos. 13 and 14), Akt-1/2/3 (Nos. 15 and 16), or actin as the control (Nos. 1 and 2). The data were analyzed and graphed.
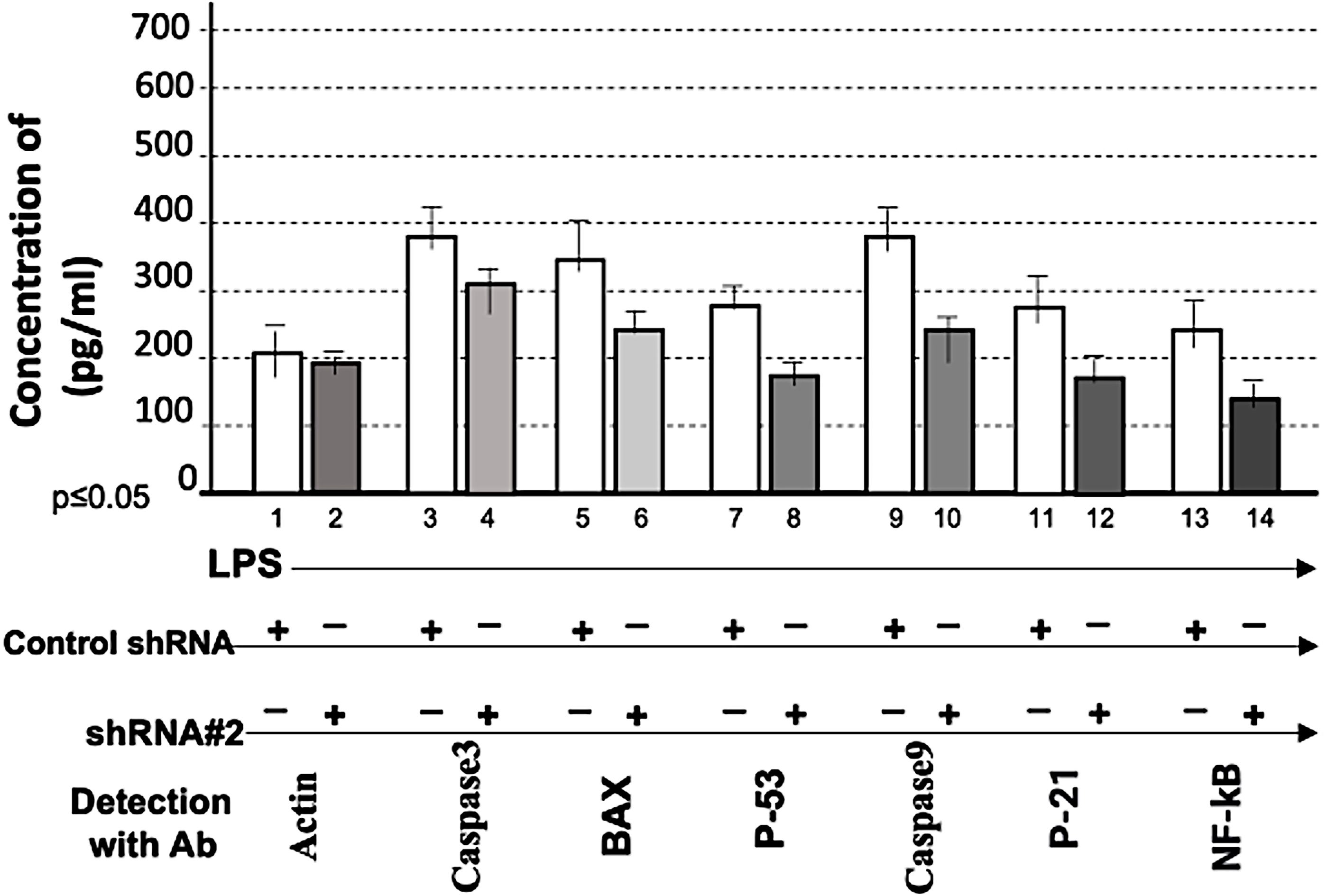
Effect of shRNA#2 on LPS-induced proteins. 1 × 105 THP-1 cells were cotreated with 0.1 µg/mL E. coli LPS plus the control shRNA or shRNA#2. The intracellular protein from each group was collected and detected by hybrid method for the quantitative detection of caspase 3 (Nos. 3 and 4), BAX (Nos. 5 and 6), P-53 (Nos. 7 and 8), caspase 9 (Nos. 9 and 10), p-21 (Nos. 11 and 12), NF-kB (Nos. 13 and 14) or actin as the control (Nos. 1 and 2). The data were analyzed and graphed.
Discussion
The reason that many inflammatory conditions and diseases are associated with high CXCL1 expression has remained unclear. While numerous studies have shown that abnormal CXCL1 production is part of the immune response to inflammation (Kunisada et al., 2017), they have not determined whether CXCL1 acts merely as a marker or as an active stimulator of disease progression. We sought to clarify whether CXCL1 functions solely as a disease-response marker or as a biologically important protein contributing to inflammation.
We hypothesized that if CXCL1 is only a response marker, inhibiting its expression would not reduce inflammation or disease progression, since it would not play a causative role. Conversely, if CXCL1 is an active participant in disease processes, its inhibition could alleviate related inflammation. In this study, treatment of THP-1 cells with either a JAK inhibitor or shRNA #2 significantly reduced LPS-induced CXCL1 and cytokines such as TNF-α. These findings suggest that CXCL1 may function as a regulatory mediator in the LPS-dependent inflammatory pathway, rather than merely serving as a marker of disease progression. CXCL1 knockdown reduced multiple signaling pathways, suggesting a potential upstream regulatory role. However, these broad effects may also reflect indirect or off-target actions and should be interpreted cautiously. The use of CXCL1-specific shRNA, along with control shRNA and validation under both endogenous and overexpression conditions, supports the specificity of CXCL1 targeting.
We acknowledge that only a single CXCL1-targeting shRNA construct (shRNA#2) is presented in this study. Although multiple shRNA candidates were initially designed and screened, shRNA#2 was selected based on its superior knockdown efficiency. Nevertheless, the use of additional independent shRNA sequences would further strengthen the reproducibility and specificity of the observed effects and should be considered in future studies.
Although formal cell viability or toxicity assays were not performed, no obvious changes in cell morphology or growth were observed during the experiments, suggesting that the observed effects are unlikely to be solely due to cytotoxicity. Nevertheless, given the broad suppression of multiple signaling pathways, future studies incorporating quantitative viability assays would further strengthen these findings.
Measuring low concentrations of intracellular cytokines by Western blot proved challenging. Although ELISA can address this limitation, analyzing multiple proteins requires several kits, which can be costly and time-consuming. To overcome this limitation, we employed a hybrid protein detection approach combining features of ELISA and protein array methodologies (Edabashi et al., 2023). This approach enabled the analysis of multiple targets with improved efficiency. While this method shows potential as a cost-effective and scalable technique, it should be considered a proof-of-concept that requires further validation and optimization.
Cloning CXCL1 cDNA in this study provides a powerful tool to examine CXCL1’s biological function in human or mammalian cells. Our data indicate that LPS induces not only CXCL1 production but also TNF-α, IL-1β, IL-1α, IL-10, and caspase 3 in THP-1 cells. To further investigate the regulation of gene expression, we cloned promoter regions of cytokines such as TNF-α and IL-1β, as well as transcription factors such as NF-κB and LITAF, into the pGL3-basic plasmid as reporter constructs (data not shown). These systems will enable future studies to delineate the biological function and regulatory mechanisms of CXCL1.
While our findings support a regulatory role for CXCL1 in LPS-induced cytokine production, additional mechanistic studies are required to establish direct causality. In particular, rescue experiments in which CXCL1 expression is reintroduced following shRNA-mediated knockdown would help confirm the specificity of the observed effects. Furthermore, the involvement of CXCL1 receptor (CXCR2) signaling and downstream signal transducer and activator of transcription (STAT) activation was not directly assessed in this study. Previous studies have demonstrated that STAT-dependent signaling pathways regulate chemokine expression (Tang et al., 2011). In addition, CXCL1 has been reported to signal through CXCR2 and may engage downstream JAK/STAT-related pathways, providing a plausible mechanistic framework for the observed effects in this study (Hu et al., 2021). Future studies will be necessary to validate these pathways.
This study was conducted using THP-1 monocyte-like cells, which represent a widely used in vitro model for investigating inflammatory responses. However, as an immortalized cell line, THP-1 cells may not fully recapitulate the behavior of primary human monocytes or macrophages. Therefore, further studies using primary human cells and in vivo models will be important to validate the physiological relevance and translational potential of these findings.
Conclusion
CXCL1 gene expression appears to contribute to the regulation of inflammatory responses. In this study, suppression of CXCL1 production using inhibitors such as a JAK inhibitor or CXCL1 shRNA #2 significantly reduced LPS-induced TNF-α, IL-1α, and IL-10 levels. Compared to controls, treatment with shRNA #2 decreased LPS-induced pro-inflammatory cytokines, chemokines, kinases, transcription factors, and apoptotic genes. These findings provide insight into the mechanism of CXCL1 in the LPS-dependent pathway and may inform the development of future therapeutic strategies for inflammatory diseases; however, additional mechanistic studies are needed to fully elucidate the underlying signaling pathways.
Authors’ Contributions
X.T. and S.D. designed the study. T.E. and X.T. drafted the article. T.E. and X.T. performed the experiments and analyzed the data. All authors provided the final approval of the article and agreed to be accountable for all aspects of the work.
Footnotes
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
No funding was received for this article.