
Abstract
Keywords
ADHD is a heterogeneous psychiatric disorder with core symptoms of inattention, hyperactivity, and impulsivity. ADHD affects 6% to 8% of school-aged children (Faraone, Sergeant, Gillberg, & Biederman, 2003) with symptoms persisting into adolescence and adulthood in about 50% of cases (Wilens & Dodson, 2004; Wilens, Faraone, & Biederman, 2004). Several neuroimaging studies have demonstrated dysfunctional neural activities within fronto-subcortical networks in patients suffering from ADHD (Arnsten, 2009; Plichta et al., 2009; Wolf et al., 2009). Particularly, the prefrontal cortex was suggested to be critically involved in dysfunctional executive control and regulation of currently ongoing behavior in ADHD (Arnsten, 2006; Wolf & Vasic, 2010).
As a proxy for the clinical symptom of impulsivity (e.g., Enticott, Ogloff, & Bradshaw, 2008; Ruchsow et al., 2008), response inhibition defined as the ability to inhibit inappropriate response tendencies has already been investigated in children (Fallgatter et al., 2004; Pliszka et al., 2006; Pliszka, Liotti, & Woldorff, 2000), adolescents (Schulz et al., 2004), and adults with ADHD (Chamberlain et al., 2007; Fallgatter et al., 2005). In adult ADHD, recent studies consistently demonstrated decreased activation of a fronto-striatal and a parietal, attentional network to be correlated with symptom severity during response inhibition tasks (Cubillo, Halari, Ecker, et al., 2010; Cubillo, Halari, Giampietro, Taylor, & Rubia, 2010; Schneider et al., 2011). Previous studies also showed that behavioral deficits of ADHD patients in response inhibition appeared strongly associated with dysfunctional error processing (van Meel, Heslenfeld, Oosterlaan, & Sergeant, 2007; Wiersema, van der Meere, & Roeyers, 2005), and deficits in error processing may contribute to a decreased ability in patients with ADHD to implement adequate behavioral control (Graf et al., 2011). Indeed, electrophysiological studies investigating inhibition and error processing during stop signal and Go/NoGo-response-inhibition tasks (McLoughlin et al., 2009; O’Connell et al., 2009; Schachar et al., 2004; Wiersema, van der Meere, & Roeyers, 2009) have already indicated limitations in error monitoring in patients with ADHD.
The close relationship between both executive functions has also received support from previous neuroimaging studies demonstrating that the neural network involved in the inhibition of inappropriate response tendencies shares commonalities with the error-processing network, signaling failure of inhibition (Braver, Barch, Gray, Molfese, & Snyder, 2001; Horn, Dolan, Elliott, Deakin, & Woodruff, 2003; Kiehl, Liddle, & Hopfinger, 2000; Menon, Adleman, White, Glover, & Reiss, 2001; Rubia, Smith, Brammer, & Taylor, 2003). The neural system responsible for the identification of errors (i.e., differences between an intended and an actually executed response) has been intensively investigated using electrophysiology (Ehlis, Herrmann, Bernhard, & Fallgatter, 2005; Falkenstein, Hohnsbein, & Hoormann, 1995; Falkenstein, Hoormann, Christ, & Hohnsbein, 2000; Gehring, Coles, Meyer, & Donchin, 1995; Scheffers, Coles, Bernstein, Gehring, & Donchin, 1996) and functional magnetic resonance imaging (fMRI; Ullsperger & von Cramon, 2001, 2003; van Veen & Carter, 2002; van Veen, Cohen, Botvinick, Stenger, & Carter, 2001). These studies identified the anterior cingulate cortex (ACC) together with the adjacent presupplementary motor area (pre-SMA; Herrmann, Rommler, Ehlis, Heidrich, & Fallgatter, 2004; Ridderinkhof, Ullsperger, Crone, & Nieuwenhuis, 2004; Ridderinkhof, van den Wildenberg, Segalowitz, & Carter, 2004; Ullsperger & von Cramon, 2001) as the core neural substrates of error processing. In addition, previous studies on inhibition performance have reported increased error-related activation of the lateral inferior frontal cortex (IFC) encompassing the frontal operculum and parts of the anterior insular cortex (Braver et al., 2001; Debener et al., 2005; Garavan, Ross, Murphy, Roche, & Stein, 2002; Mathalon, Whitfield, & Ford, 2003; Ullsperger, Freude, & Erdmann, 2001).
Error signaling in ADHD has been mostly investigated in children using electrophysiological methods (for a review, see Shiels & Hawk, 2010). Some analyses of different event-related potentials such as error-related negativity (ERN) and error positivity (Pe) revealed that the ERN amplitude (as a marker for early, more automatic error detection) might be unchanged in children with ADHD, whereas the Pe (a later component assumed to reflect more conscious error processing) was found to be attenuated (O’Connell et al., 2009; Wiersema et al., 2005; Zhang, Wang, Cai, & Yan, 2009). Although similar findings were reported in adult ADHD (Wiersema et al., 2009) leading to the assumption that dysfunctional reactions to errors in ADHD patients already derive from dysfunctional error signaling, it is important to mention that also converse electrophysiological findings exist (Albrecht et al., 2008; McLoughlin et al., 2009). In children with ADHD, methylphenidate (MPH) has been shown to normalize fronto-cingulate hypoactivation associated with error signaling (Rubia, Halari, Mohammad, Taylor, & Brammer, 2011).
In the present study, we investigated a group of well-selected adult male patients with ADHD during event-related fMRI while performing a modified version of a combined Eriksen Flanker-Go/NoGo-task. The modified task realized Go- and NoGo-trials under congruent and incongruent stimulus arrays, with incongruent NoGo-trials representing the most demanding condition on inhibition performance and was therefore suitable for response-based analyses of error signaling defined as the difference of neural activation associated with incorrect minus correct inhibition trials. Based on previous electrophysiological and imaging findings in children and recent neuroimaging studies in adult ADHD, we expected to find decreased differential activation on errors particularly in cortical areas being part of the error-processing network, such as the IFC and the ACC.
Materials and Method
Participants
Fourteen right-handed adult men (age = 18 to 32 years) with a diagnosis of ADHD according to Diagnostic and Statistical Manual of Mental Disorders (4th ed.; DSM-IV; American Psychiatric Association, 1994) criteria (combined type: n = 10; predominantly inattentive type: n = 2, predominantly hyperactive-impulsive type: n = 2) were recruited from the outpatients unit of the Department of Psychiatry, University of Würzburg, Germany (see Table 1, for details on demographics and psychopathology). Diagnosis of ADHD in adulthood employed the Structured Clinical Interview for DSM-IV Axis I and II Disorders (SCID-I and SCID-II) and was performed by an experienced clinical psychiatrist specialized in ADHD and reconfirmed by a second psychiatrist. The diagnosis of childhood manifestation of ADHD was retrospectively assessed with the DSM-IV symptom list for ADHD (17 items) and the German version of the Wender Utah Rating Scale (WURS; Retz-Junginger et al., 2007; Ward, Wender, & Reimherr, 1993). To ensure diagnostic validity, the WURS was also used to collect additional information from partners, relatives, friends, and from school report certificates. ADHD patients with concomitant Axis I and Axis II disorders according to DSM-IV (following a SCID-I and SCID-II for DSM-IV) were not included. Other exclusion criteria were history of dependence on illicit drugs and alcohol, current intake of psychotropic medication (other than psychostimulants), and any signs of sensor-motor deficits or other neurological disorders. Further general exclusion criteria were manifest reading disabilities (based on patients’ self-reports), and an IQ level below 80 (as measured by the multiple-choice vocabulary test [MWT]; Lehrl, 2005; Lehrl, Triebig, & Fischer, 1995). All patients had a history of MPH treatment. At the time of the fMRI scanning, seven patients had been off medication for at least 6 weeks, whereas the remaining seven patients had discontinued their treatment with MPH 4 days prior to the scanning procedure, that is, for at least 5 half-lives (Swanson & Volkow, 2002).
Demographic and Psychopathologic Characteristics of the Participants.

Note: MWT-B = Mehrfachwahl-Wortschatz-Intelligenztest.
Group differences were tested by means of two-sample t tests (two-tailed; uncorrected).
Rated by the multiple-choice vocabulary test (MWT-B; Lehrl, Triebig, & Fischer, 1995; Lehrl, 2005).
Rated by the Edinburgh handedness questionnaire.
Group differences tested by means of Fisher’s exact test (χ2 = 0.01; p > .61).
Cutoff score ≥30 (Retz-Junginger et al., 2007).
The healthy control group consisted of 12 men group-matched for age (age = 19-26 years), education, IQ, and handedness (Table 1). Healthy controls had neither any history of Axis I or II pathology nor any signs of a neurological or psychiatric disorder. In both groups, a validated German version of the Adult ADHD Self-Report Scale (ASRS; Kessler et al., 2005; Reuter, Kirsch, & Hennig, 2006) was used to additionally assess current ADHD symptoms.
All participants received a small financial compensation for participation. The project was approved by the local Institutional Review Board. Written informed consent according to the Declaration of Helsinki in its latest version was obtained from all participants following a complete description of the purposes and risks of the study.
fMRI Paradigm
The task (Figure 1) used in this study is a combination of a Go/NoGo task and an Eriksen Flanker paradigm (Eriksen & Eriksen, 1974) established in several electrophysiological studies on error processing (e.g., Ruchsow et al., 2006; Ruchsow, Spitzer, Gron, Grothe, & Kiefer, 2005). Five-letter strings were created from the letters R, U, P, and V with the action relevant targets always midstanding. Participants were instructed to focus on this target letter in the middle of an array. During Go-trials presenting the target letter R, participants had to respond with their right index finger on a two-button box and with the middle finger for the target letter U. In NoGo-trials, participants were instructed to withhold the response on appearance of target letters P and V. Target and flanker stimuli were combined either congruently or incongruently. In congruent trials, all five letters were the same. In incongruent Go-trials, Go-targets were flanked by visually similar NoGo-target letters (e.g., VVUVV). In incongruent NoGo-trials, the central NoGo-target was flanked by visually similar Go-targets. To obtain a sufficient number of errors, the instruction emphasized speed over accuracy on Go-trials. Combination of the factors condition (Go, NoGo) and type (congruent, incongruent) yielded 66 trials of each level resulting in a total of 264 trials. Presentation of each trial began with a centrally presented fixation cross for a period of 500 ms. Afterward, letter strings were centrally shown for a duration of 200 ms followed by a blank screen with a duration of 700 ms. According to the participants’ performance, the German expressions for either “correct,” “wrong,” or “faster” were presented for 500 ms at the end of the trial. Duration of the whole task was 22 min.

The modified Eriksen Flanker-Go/NoGo paradigm for functional challenge during fMRI.
Presentation of the experimental task was run via ERTS software (Experimental Run Time System, ©2009 BeriSoft Cooperation). Visual stimuli were centrally presented on MR compatible video goggles. Reaction times and correctness of participants’ responses per each trial were automatically registered by a standard personal computer that also controlled the sequence of trials. The task was implemented in a rapid event-related fMRI design. Mean intertrial interval was 3.01 s and average stimulus-onset asynchrony for events of the same combination of condition by type was 19.5 s.
Participants got acquainted with this challenging task during a training session on the day of MRI scanning. The training version of the task was different from the test version regarding the trial sequence and the total number of trials, but was identical with regard to the frequency of occurrence of the eight conditions.
Functional Data Acquisition
Imaging data were acquired on a 3T MAGNETOM Allegra (Siemens AG, Erlangen, Germany) head-only MRI system using a standard transmit-receive head coil. T2*-weighted functional images were obtained using gradient echo-planar imaging (EPI) in axial orientation along the AC-PC-line. Image size was 64 × 64 pixels (3.6 × 3.6 mm pixels). The volume consisted of 33 slices (TR = 2,200 ms, TE = 39 ms, BW = 3,906 Hz/Pixel). Slice thickness was 3 mm with a gap of 0.75 mm (25%). For each session, 608 volumes were acquired. The first 10 volumes of each session were discarded to avoid T1 equilibration effects. At the end of an experimental session, high resolution T1-weighted anatomic images were obtained using three dimensional MPRAGE sequences (BW = 130 Hz/Pixel, TR = 2,300 ms, TI = 1.1 s, TE = 3.93 ms, flip angle = 12°) and at least 160 contiguous slices of 1 mm thickness in sagittal direction depending on the left-to-right extension of the participants’ brain.
Data Analysis
Behavioral data analysis
Reaction times in units of milliseconds (ms) and error rates (%) were computed for the different combinations of factors response type (Go/NoGo), condition (congruent/incongruent), and accuracy (incorrect/correct). Mean reaction times and mean error rates were compared between groups using separate analyses of variance with a between-subjects factor group (ADHD, controls). The nominal level of significance was set to p < .05.
Analysis of fMRI data
Image preprocessing and statistical analyses were carried out using Statistical Parametric Mapping (SPM5, Wellcome Department of Cognitive Neurology, London, UK) and MATLAB 7.3 (Math-Works, Natick, Massachusetts). Data from each experimental session were preprocessed, including slice timing, realignment, and normalization into a standard template (Montreal Neurological Institute). Smoothing was applied with a 10-mm full width at half maximum (FWHM) isotropic Gaussian kernel. For normalization purposes, high resolution individual T1-weighted images were used that had been coregistered to the individual mean EPI obtained by spatial realignment.
On the single-participant level, individual event types (see below) were modeled as trains of delta functions at each stimulus onset convolved with a canonical hemodynamic response function (HRF). In the framework of the general linear model, the association between each experimental regressor and the experimental time course was calculated and served as an estimate for the magnitude of neural activity for each event type. A second set of regressors was added to the design matrix using the six motion parameters acquired during the realignment procedures. During calculation of parameter estimates for each regressor, time series were scaled to a grand mean of 100 over all voxels and volumes within a session. Low frequency drifts were removed via a high-pass filter using low-frequency cosine functions with a cutoff of 128 s. Model estimation was corrected for serial correlations using a first-order autoregressive model. Tabulated coordinates are peak maxima in a given cluster according to the standard T1-MNI-template (Montreal Neurological Institute). In the first-level analyses, eight different regressors were involved, which were defined by the combination of the factors condition (Go/NoGo), type (incongruent/congruent), and accuracy (incorrect/correct). Errors of commission were defined as regressors of interest. Because of the rare and inconsistent appearance, missed Go-events (errors of omission) were individually modeled but not further considered. In addition, the factor type of target letter was not further considered during the analysis of behavioral and fMRI data. Condition effects were computed using signed one-tailed t contrasts. These individual contrast images were transferred to a group analysis within the framework of random effect modeling to account for interindividual variances. For fMRI data analysis at the group level, neural error signaling was computed for incongruent NoGo-trials only because not all participants had consistently committed erroneous responses on congruent Go- and NoGo-trials.
To identify differential brain activity related to NoGo-errors, neural activity associated with incorrect minus correct NoGo-trials was computed as an average across both groups. The resulting statistical parametric map was thresholded at a significance level of p < .05, family-wise error (FWE) corrected for multiple comparisons and served as an explicit inclusive mask for the computation of between-group differences, which were analyzed using single-tailed t contrasts for group-by-condition interaction analyses. Again, the significance threshold was set to a level of p < .05, FWE corrected.
In addition, correlation analyses were performed in the ADHD group using the simple regression model in SPM5 between individual differential neural activity on NoGo-errors (incorrect minus correct) and individual global scores of the ASRS, as well as individual scores for either subscale of hyperactivity and inattention. Correlation analyses were locally constrained to the neural circuitry associated with error processing computed as an average across groups (see above). Significance was set to a level of p < .05, uncorrected at the voxel level.
Results
Behavioral Results
Although analysis of fMRI data was restricted to correct and incorrect responses during incongruent NoGo-trials, performance and reaction time data were tested on group differences (ADHD vs. controls) across the entire data set obtained during the fMRI task. Error rates for Go-trials did not significantly differ between groups, F(1, 24) = 0.451; p = .508, or between conditions, F(1, 24) = 2.42; p = .133. In addition, the interaction of both main factors was not significant, F(1, 24) = 0.21; p = .651. Analysis of reaction times for correct Go-trials, however, yielded a significant effect of the factor condition, F(1, 24) = 6.11; p = .021. Averaged over groups mean reaction times on incongruent Go-trials (478 ms; SD = 44.9 ms) were significantly slower than for congruent Go-trials (466; SD = 41.2 ms). Again, there was neither a significant difference between groups, F(1, 24) = 2.40; p = .134, nor a significant condition-by-group interaction, F(1, 24) = 0.063; p = .804.
For the NoGo-condition of the fMRI task, an analysis of variance revealed a significant effect of the factor condition, F(1, 24) = 9.76; p = .005, on error rates. However, neither the main factor group, F(1, 24) = 2.03; p = .167, nor its interaction with condition, F(1, 24) = 1.98; p = .172, was significant. This was confirmed by a reduced and separate test of group differences on erroneous incongruent NoGo-trials that subsequently entered the analysis of fMRI data. No differences between groups were observed (t value 0.93, p = .363). In addition, erroneous reaction times for this condition did not differ between groups (t value 1.36, p = .187) although ADHD patients showed numerically slightly increased reaction times compared with controls (ADHD: M = 479 ms; SD = 107.4 ms; controls: M = 428 ms; SD = 57.5 ms).
Functional Imaging Results
Across groups, a network including bilateral IFC, ACC, and the pre-SMA exhibited significantly greater activation for incorrect minus correct NoGo-trials (Figure 2). In particular, the network comprised the left (x = −30, y = 22, z = −6, Z = 6.32, BA 47) and right (x = 34, y = 18, z = −16, Z = 5.78, BA 47) inferior frontal gyrus, the left (x = −2, y = 20, z = 38, Z = 4.80; BA 32) and right anterior cingulate gyrus (x = 6, y = 30, z = 22, Z = 5.28; BA 24/32), and the superior frontal gyrus (x = 10, y = 12, z = 58, Z = 4.59; BA 6). Bar graphs in Figure 2 depicting mean neural activity for each combination of factors group and condition at the peak voxels of clusters referred above demonstrate that the relationship of mean neural activity on incorrect and correct trials was similar in both groups.
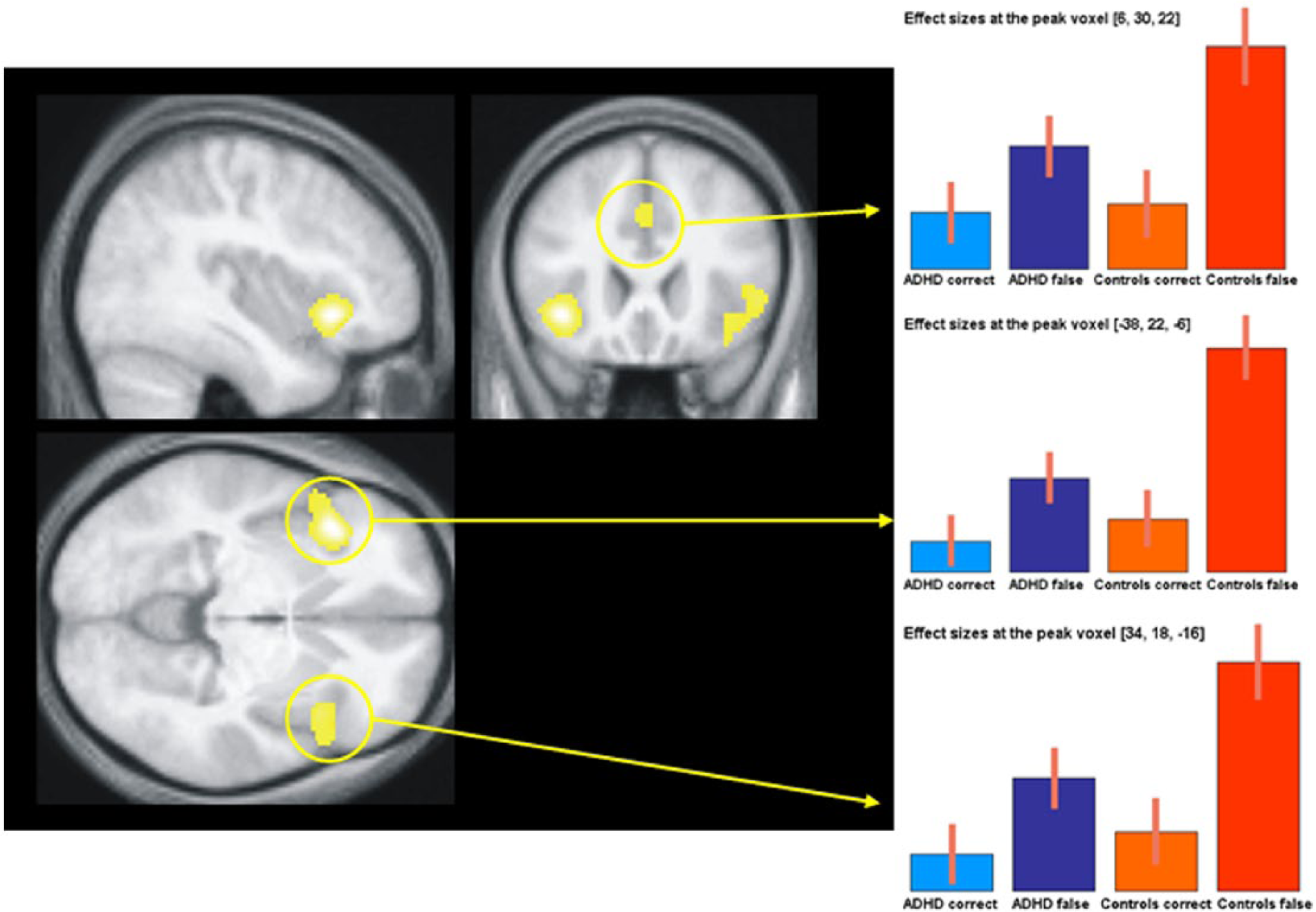
Main effect of error signaling averaged over both groups (ADHD patients and healthy controls).
A test on group differences (controls minus ADHD) of neural error signaling (false–correct) revealed significant (p < .05, FWE corrected) effects exclusively in the left inferior frontal gyrus (x = −44, y = 22, z = −4, Z = 3.76; BA 47; Figure 3). The inverted contrast (ADHD minus controls) did not show significant results even at lowered statistical thresholds. Separate regression analyses in the ADHD group for each of the regressors expressing symptom severity (global score of ASRS as well as subscores for hyperactivity and inattention) did not yield significant results.

Significant group-by-response type interaction in the left inferior frontal gyrus.
Discussion
In the present study, we investigated neural correlates of error signaling in adult patients with ADHD and healthy controls obtained by an Eriksen Flanker-Go/NoGo-task during fMRI. Behaviorally, only condition effects of congruent and incongruent letter arrays were evident for mean reaction time data on correct Go-trials and for error rates during NoGo-trials supporting sufficient overall engagement in performing the task. However, no between-group differences were observed, neither for error rates nor for reaction time data during Go- or NoGo-conditions, supporting that comparing brain activations between both groups is not misguided by differing levels of performance (Price & Friston, 1999).
Comparing failed minus successful response inhibition on incongruent NoGo-trials averaged across groups revealed significant effects in brain regions comprising the bilateral IFC (BA 47), bilateral ACC (BA 24/32), and the superior frontal cortex, including pre-SMA (BA 6). These areas showed similar activation patterns in patients with ADHD and healthy controls with significant neuronal activity during successful response inhibition and an additional increase of activation when response inhibition failed (Figure 2). This observation is in good accordance with previous studies reporting the same regions to be involved in executive function response inhibition and error monitoring (Matthews, Simmons, Arce, & Paulus, 2005; Picton et al., 2007; Ramautar, Slagter, Kok, & Ridderinkhof, 2006; Wager et al., 2005). This also indicates that the task was adequate to trigger the same set of processes in both groups. When compared with the healthy controls, ADHD patients showed significantly decreased neural error signaling exclusively in the left IFC (BA 47). The inverted contrast (ADHD patients minus healthy controls) revealed no statistically reliable differences.
Regarding the functional role of the IFC, the involvement of this structure has been reported under a series of different conditions. In the context of response inhibition, a number of previous imaging studies implementing Go/NoGo or Stop-Signal tasks have suggested that the IFC may represent one of the core parts of the response inhibition network (Chevrier, Noseworthy, & Schachar, 2007; Rubia et al., 2001; Rubia et al., 2003). However, the role of the IFC appears to go beyond mere response inhibition (Aron, Robbins, & Poldrack, 2004), and a more general interpretation relates its activation to the inhibition of interference from a prepotent representation to allow for the selection of task appropriate information or representations involved in maintaining goals/sets (Cardillo, Aydelott, Matthews, & Devlin, 2004). In addition, particularly the right IFC was reported to be implicated in category learning (Freedman, Riesenhuber, Poggio, & Miller, 2001), visuo-motor conditional learning (Passingham, Toni, & Rushworth, 2000), memory retrieval (Fletcher, Shallice, Frith, Frackowiak, & Dolan, 1998), and even memory encoding (Bor, Duncan, Wiseman, & Owen, 2003). The left IFC has also been reported to be involved in successful response inhibition during Go/NoGo-tasks (Konishi et al., 1999; Menon et al., 2001). However, tasks requiring set-switching or set-shifting (Dove, Pollmann, Schubert, Wiggins, & von Cramon, 2000; Monchi, Petrides, Petre, Worsley, & Dagher, 2001) demonstrated its significant activation. Furthermore, the left IFC was implicated in executive processes such as the monitoring of interference (D’Esposito, Postle, Jonides, & Smith, 1999; Jonides, Smith, Marshuetz, Koeppe, & Reuter-Lorenz, 1998) or the selection of relevant information (Cardillo et al., 2004; Hirshorn & Thompson-Schill, 2006; Thompson-Schill, D’Esposito, Aguirre, & Farah, 1997; Thompson-Schill, D’Esposito, & Kan, 1999; Thompson-Schill et al., 2002).
To provide a more integrative account of those various IFC functions, Dosenbach and colleagues (2006) approached this issue empirically with a meta-analysis of 10 different fMRI paradigms permitting the analysis of sustained and transient neural activities. These tasks were analyzed regarding task-set specific processes defined as “task-set initiation activity,” “task-set maintenance activity,” and “error-related task-set activity.” Beyond the dorsal ACC bordering the medial superior frontal cortex, the inferior frontal cortices around the pars opercularis were summarized to form the “core” of a human task-set system. The authors considered this task-set system to perceive bottom-up signals about ongoing performance, indicating discrepancies about the intended and actual outcome. The system either reflects or generates error-related signals that in turn help to adjust ensuing top-down signals. This empirically based theoretical account of inferior frontal involvement during error signaling is further supported by recent research on the neural representations of intentional memories suggesting that the implementation of goals, that is, intended acts to recall or remember a specific task-set, is controlled by the IFC (Badre & Wagner, 2007). In accordance with the suggested interpretation of IFC functioning (Dosenbach et al., 2006), we argue that the attenuated neural error signal in adult ADHD may reflect an attenuated neural representation of a task-set system with error-related violations of this representation resulting in decreased error signals.
In the present study, no significant behavioral differences between patients and healthy controls emerged, which is in line with previous fMRI studies on response inhibition in adult ADHD (e.g., Cubillo, Halari, Ecker, et al., 2010; Cubillo, Halari, Giampietro, et al., 2010; Dibbets, Evers, Hurks, Marchetta, & Jolles, 2009). Possibly, in contrast to the more complex and interfering conditions characterizing cognitive tasks of everyday life, the rather isolated experimental situation of being placed in an MRI scanner may have beneficially contributed to a reduction of impairments in response inhibition. Furthermore, severity of ADHD symptoms observed during childhood appears to decrease with increasing age, which may be a consequence of behavioral adaptation and/or cerebral maturation (Bramham et al., 2012; Herrmann et al., 2010). In this context, it is of note that the present ADHD sample consisted of relative stable patients with moderate impairments who were mostly well integrated in their communities having a regular job and normal social activities. In addition, the actual subtype of ADHD might also have an impact on response inhibition functioning.
In summary, compared with controls, our data showed hypoactivation of the left IFC in adults with ADHD during error signaling of failed inhibition. This might represent a neural marker of dysfunctional monitoring of ongoing behavior due to decreased task-set related memory representations leading to further deficits in error processing, for example, posterror adjustments.
Footnotes
Acknowledgements
The authors would like to thank Andrea Boreatti-Hümmer and Monika Heine for assistance in recruiting patients and Cosimo Izzi and Eva Gräßle for assistance during collection of data.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the Deutsche Forschungsgemeinschaft (KFO 125/1-1).