
Abstract
Introduction
ADHD is the most common neurodevelopmental disorder, with a worldwide prevalence rate of 3.4% (Polanczyk, de Lima, Horta, Biederman, & Rohde, 2007). Although the core symptoms of the disorder are inattention, hyperactivity, and impulsivity (American Psychiatric Association [APA], 2000), social and emotional deficits have recently been included as inherent features of ADHD. In previous major reviews, children with ADHD appear to display inadequate social behavior (Nijmeijer et al., 2008) and to have impaired social cognition (Uekermann et al., 2010). Socioemotional problems in ADHD are associated with poorer prognoses (Greene, Biederman, Faraone, Sienna, & Garcia-Jetton, 1997), and present as a risk factor for psychiatric comorbidities (Mary et al., 2016). That such socioemotional problems can be if at least partially explained by Theory of Mind (ToM) deficits stresses the importance of understanding the role of ToM as a pathophysiology of ADHD (Greene et al., 1997).
Another disease that frequently displays difficulties in the ability of interpreting the emotional states of others, such as facial expressions (Fairchild, Van Goozen, Calder, Stollery, & Goodyer, 2009) and exhibiting anger, frustrations, and poor self-regulation of attention and behavior, is conduct disorder (CD; Campbell, Gonzalez, & Silva, 1992; Caspi, Moffitt, Newman, & Silva, 1996). Children with ADHD have long been associated with a higher risk of CD comorbidity (Angold, Costello, & Erkanli, 1999; Costello, Mustillo, Erkanli, Keeler, & Angold, 2003; Yoshimasu et al., 2012). The co-occurrence of CD in children with ADHD confers risk of more severe and frequent symptoms and overall impairment, increased destructive behaviors (Steinberg & Drabick, 2015). Several common single nucleotide polymorphisms (SNPs) of oxytocin receptor gene (OXTR) are associated with phenotypic variations in callous-unemotional traits, which specify a distinct subgroup of CDs (Beitchman et al., 2012; Dadds et al., 2014). However, some recent research reported that there were no associations between CD and OXTR genotypes (Sakai et al., 2012), as well as oxytocin gene (OXT) genotypes (Malik, Zai, Abu, Nowrouzi, & Beitchman, 2012).
Although there are clear environmental influences on the developmental process of ToM ability, there is some genetic basis behind the development of ToM ability. Recent studies finding genetic associations with ToM functioning (Xia, Wu, & Su, 2012) strengthen the evidence for a genetic component to ToM functioning. Such genetic link to ToM functioning is also shown in studies on individuals with autism spectrum disorder (ASD; Nydén, Hagberg, Goussé, & Rastam, 2011). A twin study incorporating 1,116 60-month-old twin pairs has shown that environmental influences are not sufficient to completely account for the variance in ToM, even though precise heritability has not been established (Hughes, Jaffee, Taylor, Caspi, & Moffit, 2005). Because a number of common and rare genetic variants are associated with more than one psychiatric disorder (Cross-Disorder Group of the Psychiatric Genomics Consortium, 2013), it is reasonable to hypothesize that ToM deficits are also shared. One genetic variant that has been associated with ToM functioning in patients concerns the OXTR. The well-documented role of oxytocin in mammalian social behavior (Huber, Veinante, & Stoop, 2005), and the previously demonstrated effects of oxytocin administration on social abilities of humans (Radua & Shergill, 2015), predicts the OXTR SNPs to be associated with a range of general social phenotypes in the healthy population, including emotional processing, empathy, reward dependence, prosocial behavior, positive affect, stress reactivity, moral judgment, ToM, and global social cognition (Meyer-Lindenberg, Domes, Kirsch, & Heinrichs, 2011).
Although less thoroughly studied, the variants of OXT also seem to be associated with mental disorders, in particular, ASD. The OXT SNP rs2740204 has been found to be significantly associated with stereotyped behaviors in ASD (Yrigollen et al., 2008), and the OXT SNP rs2770378 is shown to be significantly associated with language impairment and restricted behavior in girls (Hovey et al., 2014) and ASD diagnosis (Chakrabarti et al., 2009).
Numerous studies that have investigated the role of the oxytocin system in social processes in healthy individuals focused on genetic variation of the OXTR (Ebstein, Knafo, Mankuta, Chew, & San Lai, 2012). To date, most studies investigating the relationship between OXTR and social phenotypes have focused primarily on rs53576 (Chen et al., 2011), rs2254298 (Tost et al., 2010), and rs1042778 (Israel et al., 2009).
Single SNPs and haplotypes within the OXTR were found to be associated with ASD (Jacob et al., 2007; Lerer et al., 2008; S. Wu et al., 2005). There is increasing evidence that ADHD and ASD are neurodevelopmental disorders, which show overlapping symptoms and share some underlying genetic liability (Ronald, Simonoff, Kuntsi, Asherson, & Plomin, 2008). Park et al. (2010) have shown particular SNPs to be associated with better social ability as rated on the Social and Communications Disorders Checklist (SCDC). In a recent study, whereas no significant difference in genotype distribution was observed between ADHD and control groups at rs2268493, rs13316193, and rs53576, the control group had significantly higher T allele frequency at rs2268493 (Ayaz et al., 2015). In the same study, within the ADHD group, no significant difference was observed between the Social Reciprocity Scale (SRS) scores of children with the TT genotype and the CT/CC genotypes. Our hypothesis is that SNP frequencies differ in OXT and OXTR genes between ADHD and control groups, and particular frequency differences are correlated to ToM ability in particular ADHD subgroups.
Method
The present study was conducted in the Child and Adolescent Psychiatry Clinic at Ege University Hospital in Izmir, Turkey. One hundred fifty-one patients with ADHD were recruited through the outpatient clinic from November 2013 to January 2015. An additional 100 typically developing children were recruited with an advertisement through the children of hospital staff. The parents signed and informed consent prior to their children’s participation in the study, and children were verbally asked assent prior to participation. All participants were of Turkish origin and from the Aegean region. The study was approved by the local ethics committee. The study was conducted according to the principles expressed in the Declaration of Helsinki.
Participants
For this study, a Child Behavior Checklist (CBCL; Achenbach, 1991a) and the Diagnostic and Statistical Manual of Mental Disorders (4th ed.; DSM-IV; APA, 1994) Disruptive Behavior Disorders Rating Scale–parent forms were given to the parents of 232 children aged 11 to 18 years. Participants with an inattentive score more than 1 SD greater than the age norms for CBCL and ADHD-Rating Scale-IV (ADHD-RS-IV) were invited to be evaluated to determine their respective groups. The parents and the participants were interviewed according to the Kiddie-Schedule for Affective Disorders and Schizophrenia, Present and Lifetime Version (K-SADS-PL; Kaufman et al., 1997) by three experienced child and adolescent psychiatrists. For a diagnosis of ADHD, participants had to meet six (or more) of the each of the Diagnostic and Statistical Manual of Mental Disorders (4th ed., text rev.; DSM-IV-TR; APA, 2000) inattention criteria and hyperactivity/impulsivity criteria endorsed by parents. The teacher’s report forms (TRF) were given to confirm that symptoms met the criterion of pervasiveness (Achenbach, 1991b). All participants and healthy comparison participants were drug naive. Five children who were treated for ADHD within the last year and two children who have a sibling diagnosed with ASD were excluded from the study. The same psychiatrists also performed a mental status examination of each child. When they did not achieve a consensus about participitants’ mental status, Wechsler intelligence scale for children-revised (WISC-R) was administrated by a clinical psychologist (Wechsler, 1974). Three participants who had an IQ performance below 80 were excluded. Children with significant medical conditions, such as epilepsy and children with diagnosed autism, atypical autism or Asperger syndrome, learning disability, and specific reading disorder; and children who were deemed to require further assessment for ASD as a result of the interview were excluded. In total, 69 participants were excluded from the study because of comorbidities. Specifically, reasons for exclusion included current any depressive disorder (including major depression and any other syndrome involving prominent depressive symptoms and resulting in marked distress or social impairment, n = 22), current any anxiety disorder (including separation anxiety disorder, specific phobia, social phobia, agoraphobia, panic disorder, posttraumatic stress disorder, obsessive compulsive disorder, generalized anxiety disorder, and other syndromes involving prominent anxiety symptoms and resulting in marked distress or social impairment, n = 34), or substance abuse at least once (n = 13). Participants who fulfilled the criteria for both ADHD and CD were diagnosed accordingly. All ADHD plus CD children fulfilled the criteria for early-onset CD (starting before age 10 years). To recruit clinical groups with rather homogeneous severity of psychopathology, children who only fulfilled the criteria for oppositional defiant disorder (ODD) without additionally meeting the criteria for CD were excluded. Two children were excluded from the study for not giving blood sample. The Turkish edition of ADHD-RS-IV was used to evaluate the severity of ADHD and CD symptoms (Turgay, 1994). Children with ADHD-combined subtype (n = 51, 41 boys and 10 girls; Mage ± SD = 14.21 ± 1.87 years), ADHD-inattentive subtype (n = 50, 33 boys and 17 girls; Mage ± SD = 13.86 ± 1.8 years), ADHD-combined subtype plus CD (n = 50, 42 boys and eight girls; Mage ± SD = 14.68 ± 2.05 years) were included in the study.
The children in the control group were recruited from the same community. We aimed to recruit groups that were as balanced as possible in terms of socioeconomic status. A healthy control group (41 boys and 59 girls; Mage ± SD = 14.68 ± 2.77 years) was established consisting of only one participant from each family of hospital staff and their relatives, who were informed about the study and agreed to participate. Inclusion criteria for children in the healthy control group were having no children diagnosed with ADHD in the family. The same diagnostic procedure was applied for the assessment of control participants. For inclusion in the control group, children could not have more than three symptoms of DSM-IV ADHD according to parent report. Because not all TRFs were available on all healthy controls, the control sample included children who scored in the normal range on the CBCL and Youth Self Report (YSR) and using clinician assessment, parent report of school performance and difficulties, incorporating school records when available. For evaluation of adolescence-associated emergence of depression or any externalizing problems, Global Assessment Scale (GAS; Spitzer, Gibbon, & Endicott, 1978) was applied by the psychiatrists, and those with a GAS lower than 70 were excluded.
Measures and Materials
DSM-IV Diagnostic Interviews
In the clinic sample, DSM-IV diagnoses were based on the K-SADS-PL (Kaufman et al., 1997). The Turkish version of the scale was reported to be valid and reliable for use in Turkey (Gökler et al., 2004). The interrater reliability of the K-SADS-PL Turkish version was observed to be good for ADHD (κ = .681, p < .005; Gökler et al., 2004). The K-SADS-PL was used to determine the presence of psychiatric disorders in the participants. At least one parent who could provide information about their child participated in the assessment process.
The CBCL
Data for this study were collected using the CBCL 4-18 (Achenbach, 1991a), a parent-rated form that includes 118 items that describe the child’s behavioral, emotional, and social problems over the past 6 months. These items are rated on a 3-point scale (0 = not true (as far as you know), 1 = sometimes or somewhat true, or 2 = very true or often true).
DSM-IV Disruptive Behavior Disorders Rating Scale–Parents Form
ADHD-RS-IV form was developed by Turgay (1994), based on the DSM-IV diagnostic criteria. This includes nine items regarding attention deficit, six items regarding hyperactivity, three items regarding impulsivity, eight items regarding ODD, and 15 items regarding CD. The symptoms were scored by assigning a severity estimate for each symptom on a 4-point Likert-type scale (specifically, 0 = not at all, 1 = just a little, 2 = much, and 3 = very much).
The GAS is a global measure of impairment associated with psychiatric diagnoses. It is a single rating scale of severity of impairment ranging from 1 (most impaired) to 100 (healthiest). The General Assessment of Functioning Scale was applied to determine interviewers rated each child’s highest level of functioning within the past 6 months (Spitzer et al., 1978).
ToM Battery
Reading the Mind in the Eyes Test (RMET)
In the children’s version of the RMET (Baron-Cohen, Wheelwright, Hill, Raste, & Plumb, 2001), participants have to infer the mental state of a person from pictures of the eyes only. For each picture, the children had to select a word out of four, which best described the way the person’s mental state. Each correct response earned one point, for a maximal score of 28. An English to Turkish translation of the material was validated by Girli and Tekin (2010).
Facial Emotion Recognition Task (FERT)
Sixty photos of 10 individuals (five male, five female) were chosen from the MacBrain Face Stimulus Set. The task assessed the recognition of six basic emotions (happiness, sadness, surprise, anger, disgust, and fear). The respondents were asked to name the appropriate emotion from these six alternatives. Each correct answer was awarded with one point. A total score was calculated (Ekman, 1992; Ekman et al., 1987; Ekman & Keltner, 1970).
The Unexpected Outcomes Test (UOT) is a 12-item measure of the ability to explain apparent incongruities between the emotion-eliciting context and the actual emotions elicited (Dyck, Ferguson, & Shochet, 2001). The UOT items describe a situation likely to cause an emotional response in a protagonist. It is an advanced task; the respondent should reason about the causes of emotions and is expected to present additional situational information to resolve the incongruity. This task assesses the respondent’s ability to reason about emotions that are important for the cognitive component of empathy. The answers are recorded in a booklet and scored on a 3-point (0-2) scale. A total score on the test is calculated (0-24 points). The interrater reliability for the scoring of the UOT was .91 in a pilot study (n = 40 healthy participants; Bora, Gökçen, & Veznedaroglu, 2008).
Procedures
Informed consent form from the parents and children was collected first. Based on information from the parents, the sociodemographic information form was completed by the researchers. Diagnostic interviews were conducted independently and in private with children and parents. After the diagnostic evaluation, the ToM tasks, RMET, FERT, and UOT were administrated sequentially. The entire testing session lasted approximately 45 min. The blood samples were collected after the ToM tasks were performed.
Genotyping
Genomic DNA was extracted from blood samples (2 mL) using standard protocols. Coding exons and the exon–intron boundaries of the OXT and OXTR genes were amplified with polymerase chain reaction (PCR) using the primers listed in Table 1. All PCR products were sequenced by the dye termination method using a DNA sequencing kit (Perkin-Elmer, Foster, California, USA) and analyzed using the ABI Prism 3100 sequence analyzer (Applied Biosystems, Foster, California, USA).
Primers Used for Amplifying the Coding Exons and the Exon–Intron Boundaries of OXT and OXTR Genes.

Note. OXT = oxytocin; OXTR = oxytocin receptor.
Statistical Analyses
Statistical tests were performed with SPSS Statistics version 20 (IBM). Assumptions of normality were checked for each data set and validated. The allelic and genotypic distributions between the patients and controls were examined using the Pearson chi-square test on 2 × 3 contingency tables. First, independent t tests and chi-square (cross tabulation) and ANOVAs were used to examine differences in ToM tasks performance, and OXTR genotypes. Genotype associations with social cognitive and symptom measures were evaluated using a series of ANOVAs with allele subgroups for each SNP entered as independent variables. Age and gender were included as covariates; p values <.05 were considered significant and two-tailed tests were used. In situations in which there were less than three participants in an allele group, we collapsed across categories by combining the low-frequency homozygotic individuals with the heterozygotic participants. This occurred for three SNPs: rs2228485 C>T, rs4686302 C>T, and rs237902 G>A (Table 2).
Description of OXTR Gene Polymorphisms.

Note. OXTR = oxytocin receptor; SNP = single nucleotide polymorphisms.
SNP alleles correspond to The Single Nucleotide Polymorphism database (dbSNP) (http://www.ncbi.nlm.nih.gov/SNP/).
All SNPs were tested for Hardy–Weinberg equilibrium (p > .005).
Results
ADHD group and healthy controls differed significantly in mean years of age and gender (t = −2.669, p < .05; χ2 = 32,954, p < .001). No significant difference was found between the two groups in terms of education level of mother and father, and also working status of parents (see Supplementary Table 1).
Sequencing of the OXT and OXTR genes revealed no causal mutations, but three common SNPs (rs2228485, rs4686302, and rs237902) were detected in OXTR gene. No SNPs have been detected in the OXT gene. All genotype frequencies of rs2228485, rs4686302, and rs237902 were in concordance with the Hardy–Weinberg equilibrium in the ADHD and healthy samples (p > .05). The genotype distributions for the three SNPs within the OXTR gene are summarized in Table 3. A difference was observed in the rs4686302 genotype distribution between the children with ADHD and the healthy controls, in which the TT genotype tended to be more frequent in the children with ADHD (χ2 = 4.163, p = .042). When the children with CT and TT genotype were combined (CT/TT) and compared with the children with CC genotypes, a significant association was shown between the children with ADHD and CT/TT genotypes of rs4686302 (χ2 = 3.695, p = .037). No significant association was observed between the genotype distributions or allele frequencies for the other OXTRs and ADHD. Chi-square tests revealed no significant differences for the studied SNPs between groups, and frequencies were also unrelated to gender in the overall sample (χ2 test, p > .05).
Genotype Distribution for Three SNPs Within OXTR Gene in Children With ADHD and Healthy Controls.

Note. Significant results are indicated in bold type. Count (and percentage) of healthy controls and children with ADHD in each genotype are presented. Chi-square and p values are provided alongside counts. SNP = single nucleotide polymorphism; OXTR = oxytocin receptor; OR = odds ratio; CI = confidence interval; HC = healthy controls; HW = hardy weinberg.
Pearson chi-square test between genotype and group (ADHD/HC).
C-carriers: CC/CT.
G-carriers: GG/GA.
The comparison computed on the percentage of correct responses for RMET and FERT revealed that the children with ADHD-combined plus CD performed more poorly than the other suptypes and healthy controls, FRMET(3, 247) = 55.5, p < .001; FFERT(3, 247) = 80.6, p < .001. There was a significant main effect of diagnose on UOT, FUOT(3, 247) = 54.1, p < .001. The post hoc test showed that ADHD-combined plus CD group scored lower than the ADHD-inattentive and ADHD-combined subtypes (p = .045 and p = .000, respectively).
OXTR Polymorphisms and ToM Scores
ADHD children with CT/TT genotype for the OXTR rs4686302 performed significantly worse on the face emotion recognition task than those with CC genotype (t = 2.124, p < .05; Table 4). Performances of the ToM tasks were analyzed using an ANOVA to investigate genotype effects between ADHD subtypes and healthy controls.
ToM Scores (M, SD) by OXTR rs2228485, rs237902, and rs4686302 Genotypes in Children With ADHD (n = 151) and Healthy Controls (HC, n = 100).
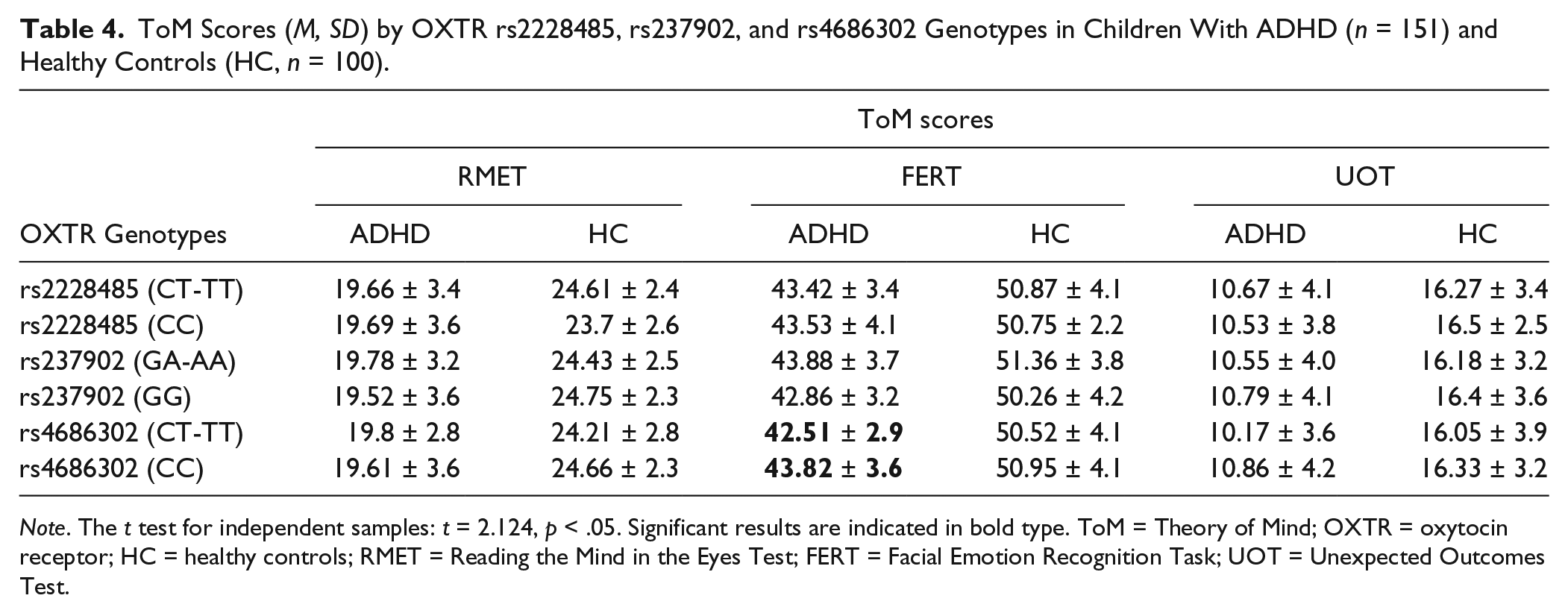
Note. The t test for independent samples: t = 2.124, p < .05. Significant results are indicated in bold type. ToM = Theory of Mind; OXTR = oxytocin receptor; HC = healthy controls; RMET = Reading the Mind in the Eyes Test; FERT = Facial Emotion Recognition Task; UOT = Unexpected Outcomes Test.
ANCOVA was computed to determine main effects of each studied rs2228485, rs4686302, and rs237902 genotypes separately as factors, as well as the covariate mean years of age and gender on the three tasks as dependent variables between ADHD groups and healthy control. Homoscedasticity of samples was confirmed by Box-M and Levene’s tests (p > .05). In ADHD-inattentive subgroup, a significant main effect of OXTR rs4686302 genotype on social cognitive tasks (RMET, FERT, and UOT) was observed after controlling for age and gender, F(1, 49) = 0.42, p = .838, η2 = .001; F(1, 49) = 10.379, p = .002, η2 = .178; F(1, 49) = 4.498, p = .039, η2 = .086. Tukey test indicated that children with the CC genotype were higher performance on FERT and UOT than were those with the CT/TT genotypes (see Figure 1).
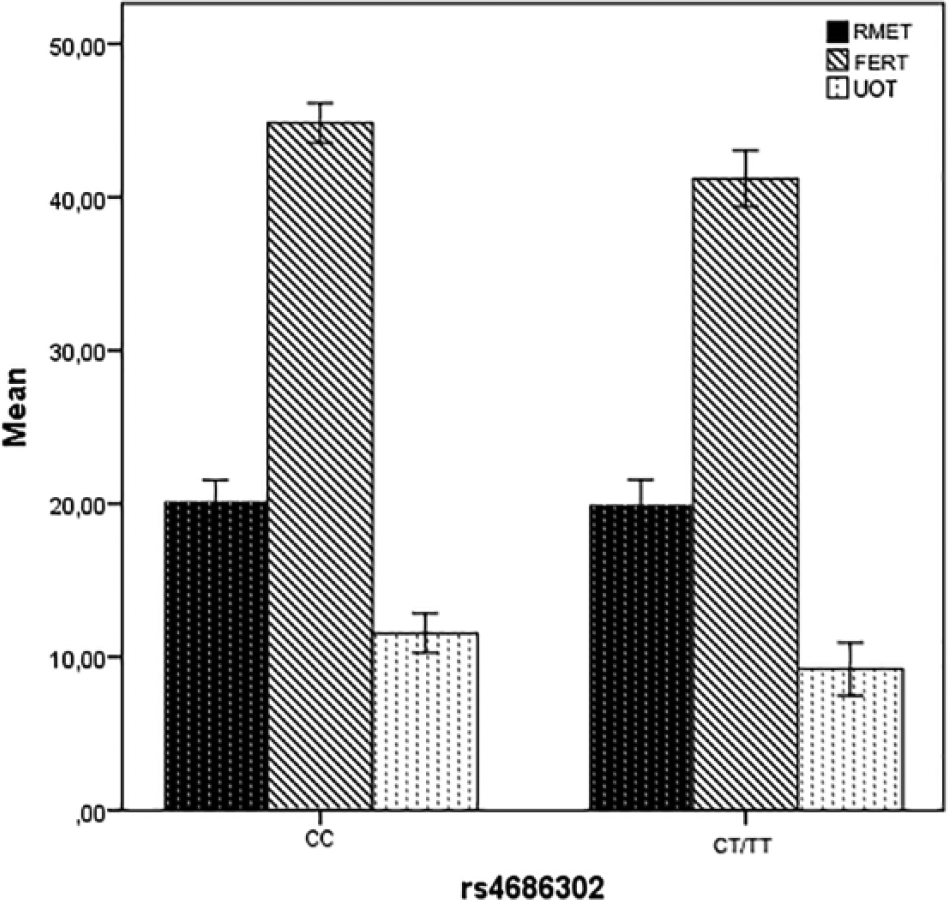
A significant main effect of OXTR rs4686302 genotype on social cognitive tasks (RMET, FERT, and UOT) was observed after controlling for age and gender in ADHD-inattentive subtype (n = 50).
As for the impact of OXTR SNPs rs2228485, rs4686302, and rs237902 on ADHD symptom severity, t tests for independent samples indicated significantly higher values on the hyperactive/impulsive (H/I) symptom severity and inattention + H/I symptom severity scores in participants with AA/AG genotypes of rs237902 (t = −2.202, p = .029; t = −2.090, p = .038; see Figure 2). No significant group differences were detected for the other OXTR genotypes.
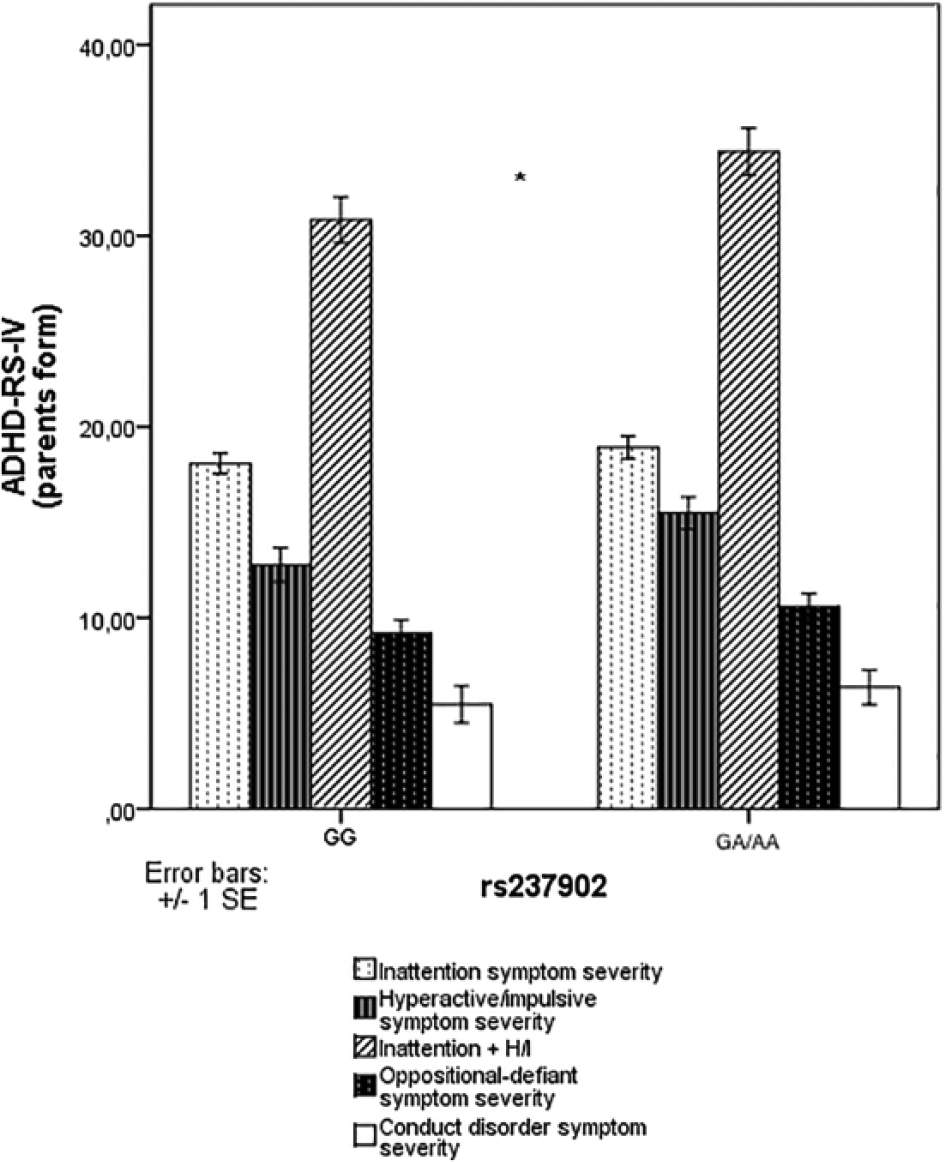
OXTR rs2228485, rs237902, and rs4686302 polymorphisms and ADHD-RS-IV (parents form) scores in children with ADHD (n = 151).
Discussion
In this study, the entire human OXT and OXTR genes, including coding sequence, exons, and exon–intron boundaries, have been sequenced for the presence of mutations and SNPs in children with ADHD and healthy samples. Results of this case control study are suggestive for an association of oxytocinergic system genes and ADHD. In particular, we found a significant association of OXTR SNP rs4686302 with the diagnosis of ADHD, the overall less frequent T allele being more frequent in the ADHD group compared with controls. In contrast to the previously referenced studies (Chakrabarti et al., 2009; Hovey et al., 2014; Yrigollen et al., 2008), we have found no OXT SNPs in the sample populations studied. To the best of our knowledge, our study is the first in literature comparing the relation of OXTR polymorphisms to social cognition tests in children with ADHD and healthy participants.
Social cognition is the ability to understand the mind of other people and is essential for successful social interaction (Uekermann et al., 2010). Recent studies have suggested that the administration of OXT could increase social cognition in humans (Domes, Heinrichs, Michel, Berger, & Herpertz, 2007) and even restore impaired social cognition in neuropsychiatric disorders (Guastella et al., 2010; Meyer-Lindenberg et al., 2011; Striepens, Kendrick, Maier, & Hurlemann, 2011). Multiple studies have identified the A allele of rs53576, situated in the third intron of OXTR, as a potential risk allele for social cognition impairment. This allele has been associated with autistic traits in Chinese participants (S. Wu et al., 2005) and Japanese participants (Liu et al., 2010). Children with ADHD have significant difficulties in social interactions and communication (Reiersen, Constantino, Volk, & Todd, 2007; Santosh & Mijovic, 2004), which are similar to those seen in ASD and exceed that which are found in control participants (Hattori et al., 2006). Current research indicates that this overlap is due partly to shared genetic factors. Further research in a cohort of children with ADHD found that the AA genotype at SNP rs53576 or TT genotype at SNP rs13316193 within the OXTR gene was associated with better social ability as rated on the SCDC (Park et al., 2010). In a recent study, whereas no significant difference in genotype distribution was observed between ADHD and control groups at rs2268493, rs13316193, and rs53576, the control group had significantly higher T allele frequency at rs2268493 (Ayaz et al., 2015). In the same study, within the ADHD group, no significant difference was observed between the SRS scores of children with the TT genotype and the CT/CC genotypes. Therefore, OXTR intron 3 SNPs are implicated in social dysfunction phenotypes. Unlike these reports, we investigated the exon 3 coding polymorphisms in our study, because of their potential functional relevance. We most frequently found three SNPs of the OXTR exon 3, rs2228485, rs237902, and rs4686302, in our sample population. Our findings related to rs2228485 are to some extent consistent with earlier studies. Chakrabarti and coworkers could not find an influence of rs2228485 variants on the RMET (Chakrabarti et al., 2009). In two other studies, rs2228485 did not show significant associations in studies with ASD (Chakrabarti et al., 2009; Kelemenova et al., 2010). Contrary to these results, another recent study showed that participants with the rs2228485 CC genotype performed significantly worse on pictures with male faces compared with the participants with CT/TT genotypes (Lucht et al., 2013). However, for rs2228485, no functional role has been clearly described yet.
Malik et al. (2012) reported that the OXTR SNP rs237902 C allele and the CC genotype were overrepresented in male aggressive child cases as compared with the controls (Malik et al., 2012). Evidence also suggests rs237902 has been associated with the severity of negative symptoms in schizophrenia (Montag et al., 2013). To date, the association between rs237902 genotype and ADHD has not been studied. We found rs237902 to be the singular SNP significantly associated with the severity of ADHD symptoms. In particular, significant association was found for the hyperactivity/impulsivity and inattention and hyperactivity/impulsivity symptom severity. To our knowledge, the present study is the first to provide evidence of this interaction, and this finding needs further exploration on larger samples.
Previous researches have demonstrated that oxytocin promotes emotional and cognitive aspects of empathy, by exogenous administration as well as on gene level. Wu et al. first demostrated the assocation of rs53576 and autism. Later studies examined the association of rs53576 in the OXTR gene and empathy. For trait empathy, marked by Interpersonal Reactivity Index (IRI), four of the SNPs—rs2254298, rs2268491, rs13316193, and rs4686302—were significant at the 5% level, where C carrier displayed more empathy than T allele carrier, and individuals with CT genotype (rs2254298) showed more cognitive empathy than those with TT genotype (N. Wu, Li, & Su, 2012). These findings could account for the previous results suggesting that TT genotype was significantly associated with poorer social and communication ability and higher risk of having a diagnosis of ASD (Costa et al., 2009; Park et al., 2010; Rodrigues, Saslow, Garcia, John, & Keltner, 2009). Another recent study reported that individuals with T allele at rs4686302 had better perspective taking than those with C allele (N. Wu et al., 2012). In the same study, on rs4686302, females with CT genotype had a higher empathy level than males with the same genotype. We found rs4686302 CT genotype to be the singular SNP with significant difference between ADHD and healthy control groups. Compared with the control group, individuals with T allele at OXTR rs4686302 had significantly impaired FERT performance. In addition, in the inattentive subtype, those with CT/TT genotypes had lower scores on both FERT and UOT. No studies to date investigated social cognition tests in ADHD subtypes with OXTR variants. The fact that an SNP with high frequency in the ADHD population is significantly associated with particular subtypes confirms ADHD to be a heterogeneous disorder. Our research provided preliminary evidence and a basis for further research in this area.
In summary, we could not show any role of OXT gene in the formation of ADHD but OXTR gene rs4686302 polymorphism was shown to be an important genetic marker in modifying disease severity. Our results noted the importance of the OXTR gene in modulating individual differences in children’s social behavioral skills, thus indicating that the variations of the OXTR gene were associated with different subtypes of children with ADHD.
Limitations
There are several limitations of this study. The sample size (151 cases and 100 for case control analysis) is small for genetic association studies and restrains the detection of small effect sizes. Furthermore, our statistical tests were not corrected for multiple comparisons so replication in larger samples will be essential. A second limitation, as already outlined, is that false positives are common in candidate gene studies (Hinney, Scherag, & Hebebrand, 2010; Trace, Baker, Peñas-Lledó, & Bulik, 2013). Although the middle childhood age range is ideal to select, as this is the age of peak identification of ADHD, the mean age of our sample was high. Because the ToM tasks would have performed to later childhood, we deliberately selected the study to this age range (Dyck et al., 2001). Another limitation for this study is that the biological mechanisms by which SNPs affect gene transcription were disregarded. Epigenetic modification studies of OXTRs show that the DNA methylation would affect phenotypic variability (Kumsta, Hummel, Chen, & Heinrichs, 2013). Our study assumed a dominant genetic model, whereby carriers of a single risk allele were compared with homozygotes of the other allele. Although an additive genetic model would be preferable, our sample size restricted the possibility of such analysis. Future association studies should attempt to replicate our findings across populations using larger independent samples so that all three genotypes (e.g., AA/AG/GG) can be compared directly.
As we have detected three SNPs and followed the clinical presentation of the individuals, we have faced the multiple comparisons problem, and Type I error accumulated. Similarly, the three social cognition tests employed led to an accumulation of Type I error.
Despite these limitations, this study supports further research of OXTR and related polymorphisms in ADHD, and our results may be valuable for planning future studies.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Author Biographies
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.