
Abstract
Objective:
Examining the joint effect of two functional variants in two dopamine-related genes (DRD3 and COMT) on ADHD-relevant behaviors under three experimental conditions (EC).
Method:
362 children with ADHD were assessed by parents and teachers during a week of baseline evaluation, followed by 1 week of MPH and placebo, administered in a double-blind crossover design.
Results:
Statistically significant 3-way (DRD3-by-COMT-by-EC; p = .004) and 2-way interactions (COMT by EC; p = .002) were observed on Conners’-Teachers scores. Children with the COMT Met/Met genotype had lower scores at baseline and on placebo compared to the other genotype groups. Furthermore, stratifying the children according to their COMT genotypes helped to detect statistically significant and biologically meaningful effects of DRD3 genotype.
Conclusions:
These findings suggest that COMT and DRD3 genetic variants may together play a role in ADHD symptomatology and response to treatment through gene-gene interaction.
Introduction
Attention-deficit/hyperactivity disorder (ADHD) is one of the most common psychiatric conditions in childhood with a worldwide prevalence of 5.9%–7.1% (Willcutt, 2012). Genetic epidemiological studies strongly suggest that genetic factors play an important role in this disorder. Indeed, the mean heritability estimate of ADHD was found to be 70%–80% (Faraone & Mick, 2010; Freitag et al., 2010), placing ADHD among the most heritable psychiatric disorders.
In spite of this strong heritability, molecular genetic studies have failed to show a consistent association between ADHD and specific genetic variants. Most of the candidate gene association studies conducted over the last few decades have led to inconsistent results. In addition, initial genome-wide association studies (GWAS) of ADHD with both case-control (Hinney et al., 2011; Neale et al., 2010; Stergiakouli et al., 2012; Yang et al., 2013; Zayats et al., 2015) and family-based study designs (Mick et al., 2010; Neale et al., 2008) did not discover any DNA variants that achieved genome-wide significance (p ≤ 5 × 10−8). However, recently the first genome-wide significant ADHD risk loci were published with a meta-analysis of 20,183 individuals diagnosed with ADHD and 35,191 controls (Demontis et al., 2019).
The lack of clear evidence for the involvement of single genetic variations in ADHD could be explained under the multifactorial polygenic nature of ADHD where minor contributions from each susceptibility gene are at play (Stergiakouli et al., 2012; Williams et al., 2012). Under such a scenario, much larger sample sizes would be required to identify these small effects (Button et al., 2013; Hawi et al., 2015)—which is confirmed by the latest GWAS. It is also possible that more substantial genetic effects are at play, but they are undetectable because of gene-gene (G-G) or gene–environment interactions (G×E). These cryptic genetic variants are “dormant” polymorphisms that express their phenotypic effects only under extreme conditions, such as when specific and rare combinations of alleles segregate together leading to epistatic genetic effects, or under specific environmental conditions (Le Rouzic et al., 2008; Le Rouzic & Carlborg, 2008; Paaby & Gibson, 2016). While alternative models of complex genetic architecture, such as the rare de novo mutations in a large number of cases have been proposed (Girard et al., 2015), the model of cryptic genetic variations is compatible with the known robustness of neural systems. This model often embodied as redundancy within and between functional networks, such that negative outcomes, including psychiatric manifestations, become apparent only when several “failures” occur in connected functional systems. Most importantly, a very large meta-analysis including all twin studies investigating a very large number of physical, behavioural and psychiatric traits concluded that the majority of these traits (69%) the ressemblance between twin can be accounted for by a simple additive genetic model with the notable expection of hypekenic disorders, where a non-addictive genetic component might be involved (Polderman et al., 2015).
Dopamine (DA) has a major role in modulating several aspects of human behaviors, including attention and motor behaviors, the main behavioral dimension altered in ADHD. Synaptic DA is tightly and dynamically regulated by a large number of proteins, each one implicated in specific processes regulating DA synthesis, release, turnover and signaling. External “environmental” factors such as pharmacological agents play a role in modulating the synaptic DA. Both, methylphenidate (MPH) - the main molecule used for the treatment of ADHD symptoms - and placebo have shown to act, at least partly, through DA signaling (de la Fuente-Fernandez et al., 2001, 2006; Hall et al., 2012; Rowland et al., 2002; Wilens, 2008).
Given the importance of the brain dopamine systems in controlling behaviors, it is therefore possible that robust regulatory mechanisms are involved such that deviation in phenotypic outcomes will happen only under extreme conditions, such as co-occurrence of several interacting genetic variants or under specific environmental conditions. If this hypothesis is true, attempts to identify single genes involved in ADHD risk may be futile. However, it is likely that joint investigation of functional variants in several dopamine-related genes under different DA tones (as experimentally imposed by pharmacological probes) may allow a fine-grained analysis of genetic variants in the regulation of ADHD relevant behavioral dimensions.
While several genes could be jointly selected for epistasis analysis, this approach is limited by the requirement of very large sample size, as genotype strata increase rapidly with each additional gene (e.g., 9 or 27 strata in the case of 2 or 3 bi-allelic genes respectively). However, two genes selected on the basis of strong a priori hypothesis, could serve as a proof of concept if applied in a reasonably large sample.
In a previous study, we used a comprehensive approach to investigate the role of DRD3 in ADHD (Fageera et al., 2018). The results provided some evidence on the implication of the DRD3 (Ser-9-Gly) polymorphism in the etiology of ADHD and the modulation of its various behavioral dimensions. In this study, we propose to further study the effect of this gene by studying its interaction with the Catechol-O-Methyltransferase (COMT) gene as an illustration of the gene by gene (G-G) interaction approach. Several arguments support the selection of these candidate genes. First, both genes encode proteins that regulate synaptic DA concentration and signaling consecutively. Because of these sequential functional effects, it is possible that lower DA concentration, as imposed by high enzymatic activity of the COMT, will have phenotypic effects only in case of the presence of hypoactive DRD3 receptors. Second, COMT is an essential enzyme for catecholamine metabolism regulating the concentration of DA in the prefrontal cortex (Tunbridge et al., 2004), a brain area highly implicated in ADHD (Arnsten, 2009). A common polymorphism in this gene (Val108/158Met) results in important variation of enzymatic activity (Chen et al., 2004), with the methionine isoform of the enzyme having only 25% of the activity of the wild-type valine enzyme (Lotta et al., 1995). Third, DRD3 is predominantly localized in the limbic areas of the brain. These areas are critical for cognitive and emotional functions, as well as expression of reward (Wise & Bozarth, 1984), all of which have been reported to be dysregulated in ADHD (Barkley, 2010; Martinez et al., 2016; Willcutt et al., 2005). A Ser9Gly isoform (encoded by SNP rs6280) has been associated with differences in affinity for DA, with the Gly/Gly (or C/C) being associated with a protein isoform having a higher rate of DA binding compared to the other two genotypes (Lundstrom & Turpin, 1996; Lundstrom et al., 1998). It has also been suggested that the C (Gly) allele could influence intracellular signaling pathways; yet the exact molecular basis of this effect is unclear (Hellstrand et al., 2004). Thus, it is possible that behavioral manifestations of ADHD will become apparent only when specific combinations of genetic variants in the COMT and DRD3 genes occur, particularly under some critical environmental or pharmacological conditions.
In this study, we propose to investigate the joint role of DRD3 and COMT variants in modulating ADHD behaviors. We approached this question by evaluating behavior under three experimental conditions (EC) that are theoretically associated with different synaptic levels of DA: baseline (lowest DA levels), placebo (partial increase in DA levels) and MPH (highest increase in DA levels). The target behaviors were assessed by parents and teachers using the Conners’ Global Index, a well-validated scale to assess the child’s behavior (Conners’-P and Conners’-T respectively). All behavioral assessments were conducted while being blinded to the EC.
Methods
Participants
Children with ADHD, ages between 6 and 12 years (mean = 9.03, SD = 1.79), who completed both Conners’-P/T and were successfully genotyped for both genes, were included in this study (n = 619). Children and their parents were interviewed by a psychiatrist and all children met DSM-IV criteria for ADHD. Children having an IQ < 70 or a history of Tourette’s syndrome, autism, or psychosis were excluded from the study. The study was approved by the Research Ethics Board of the Douglas Mental Health University Institute, Montreal, Canada. All parents provided written consent and children provided verbal assent after explaining them the details of the study.
Study Design
This study is part of an ongoing randomized pharmacogenetic behavioral study using placebo and MPH as pharmacological probes to modulate synaptic levels of DA. The treatments were administered according to a double-blind crossover trial and the research protocol was registered in clinicaltrials.gov, number NCT00483106. The data were collected from November 1999 to 2018 and the study was conducted at the DMHUI in Montreal, Canada. The flow of participants through the study is summarized in Supplemental Figure S1 (available online).
Following 1 week of medication wash out (if necessary), all participants received 1 week of MPH 0.5 mg/kg/day and 1 week of placebo (Supplemental Figure S2), in a divided bid schedule.
Procedures and Study Outcomes
Parents and teachers were asked to assess the behavior of the child using the Conners’-P and Conners’-T, respectively, at baseline, at the end of placebo and MPH weeks. Both parents and teachers were informed of the study design in the same manner; however, all participants were blind to the order of treatment. Both MPH and placebo were encapsulated in opaque gelatin capsules in weekly blisters, packed by a pharmacist who was not affiliated with the study. Furthermore, a statistician who was also not affiliated with the study prepared a computer-generated randomization list to determine the order of the treatment.
Genotyping
Children were invited to give blood samples or saliva, if the child is only amenable to the latter. Genomic DNA was extracted from the lymphocytes with a commercial method (QiAmp DNA Blood Mini Kit, Qiagen, Hilden, Germany) according to the manufacturer’s instructions. DRD3 rs6280 and COMT rs4680 were amplified using Sequenom (now Agena Bioscience) iPLEX Gold Assay technology at McGill University and Genome Quebec Innovation Center (Quebec, Canada) according to the standard protocol.
Statistical Analysis
Statistical analyses were conducted using SPSS version 20. Chi-square and analysis of variance (ANOVA) were applied to test for differences between categorical (e.g., sex, income, and ethnicity) and continuous variables (age, medication, full-scale IQ, Conner’s, CBCL and DISC) between the groups respectively (Table 1). To test for the effect of genotype and EC, we used repeated measure ANOVA, where genotypes in each of the two genes (DRD3 and COMT) were the two between-subject factors (three levels each) and the behaviors under the three ECs (baseline, placebo, and MPH weeks), were the within-subject repeated measure factor; ethnicity was used as a covariate.
Demographic and Baseline Characteristics of Children with ADHD Separated According to Their DRD3 rs6280 and COMT rs4680 Genotypes.
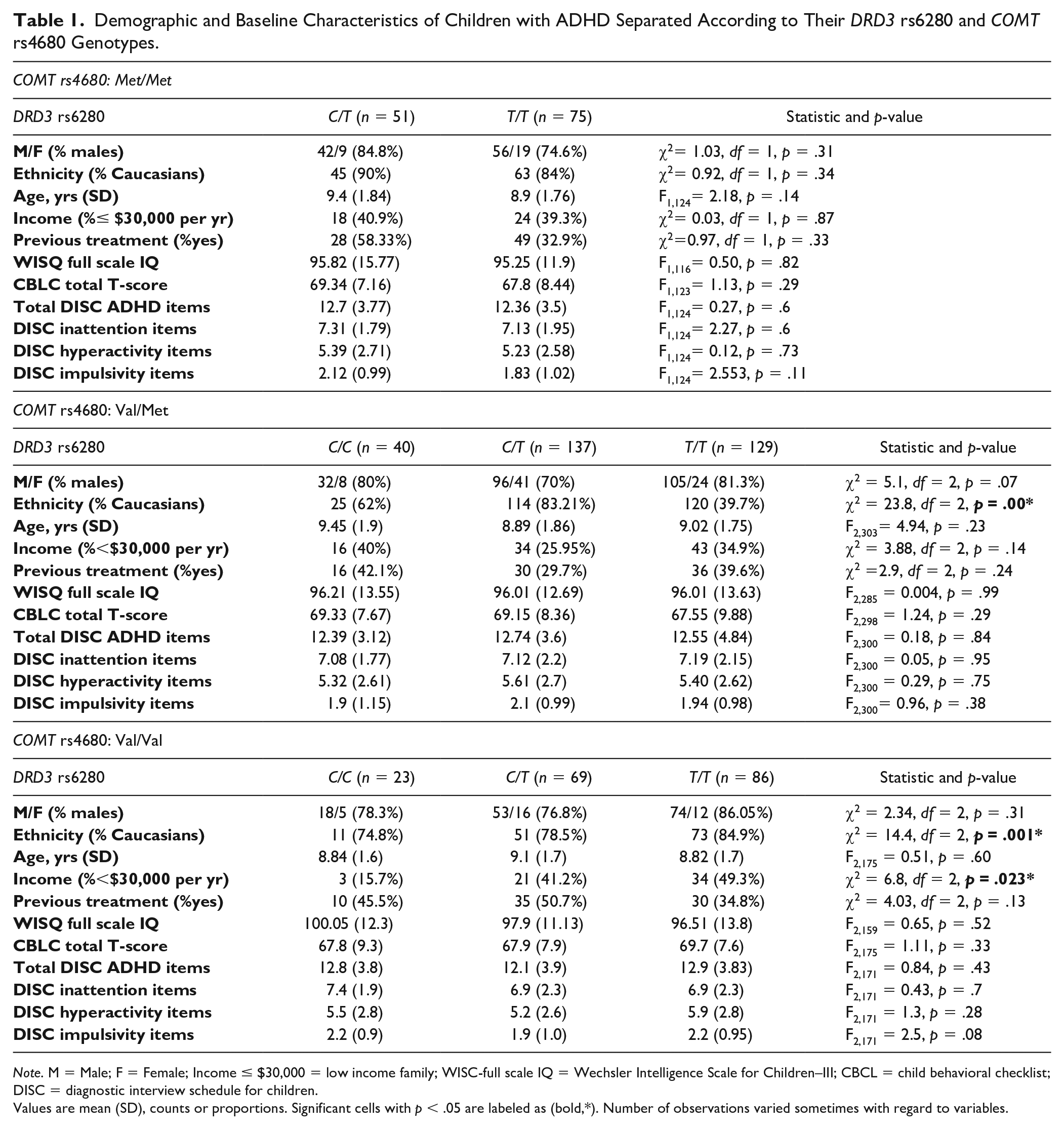
Note. M = Male; F = Female; Income
Values are mean (SD), counts or proportions. Significant cells with p < .05 are labeled as (bold,*). Number of observations varied sometimes with regard to variables.
Results
The genotype in the two SNPs tested in this study do not deviate significantly from the Hardy–Weinberg proportions (all ps > 0.05). All nine possible genotype combinations of DRD3 Ser9Gly and COMT Val158Met are presented (Supplemental Table S1). Clinical characteristics of the six-genotype groups (three groups for each gene) of children are presented in Table 2. Gender, age, and ADHD subtype were similar among all genotype groups of children. A significant difference was observed between ethnicity and genotype distribution in both genes (p < .001). Thus, we controlled for ethnicity in subsequent analyses. The COMT Met/Met and DRD3 C/C genotype combination had only nine subjects and was excluded from the analysis.
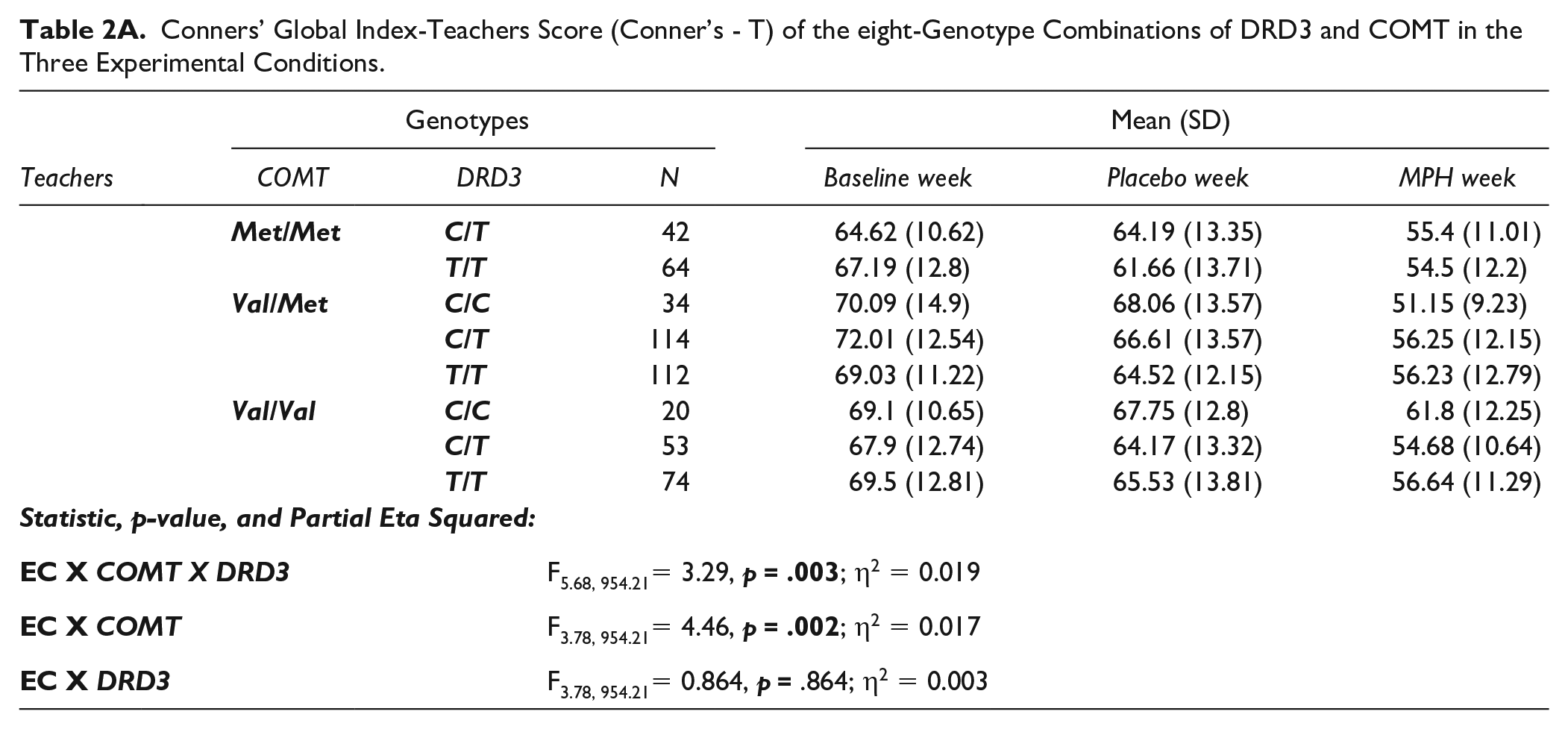
Conners’ Global Index-Teachers Score (Conner’s - T) of the eight-Genotype Combinations of DRD3 and COMT in the Three Experimental Conditions.
Conners’ Global Index-Parents Score (Conner’s - P) of the Eight-Genotype Combinations of DRD3 and COMT in the Three Experimental Conditions.

Effect of DRD3 and COMT Polymorphisms on Conners’ Teacher Scores
Mauchly’s test of sphericity indicated that the assumption of sphericity was not met (χ2 (2) = 29.18, p < .001) and a Greenhouse-Geisser correction was applied. Repeated measure analysis of variance revealed a significant COMT by DRD3 by EC 3-way interaction (F (5.7, 954.23) = 3.29; p = .004). A two-way interaction, COMT by EC, was also significant (F (3.8, 954.23) = 4.46; p = .002). No DRD3 by EC interaction was observed (F (3.7, 954.23) = 0.864; p = .48).
As expected, there was a highly significant effect of the EC (F (1.89, 954.23) = 24.6; p < .001). Post-hoc analysis showed that behavioral scores are highest at baseline (Mean = 68.9; SD = 12.3), intermediate under pla-cebo (Mean = 64.9; SD = 13.3) and lowest under MPH (Mean = 55.8; SD = 11.6) conditions (Figure 1, lower scores indicate better behavior). These observations are compatible with a gradient of dopamine tone; highest under MPH, intermediate under placebo and lowest at baseline, in conformity with our experimental design.

Conners’ global index score (Conners’-T, mean ± SEM) in the three experimental conditions: methylphenidate (MPH), placebo and during baseline. Error bars represent standard error of the mean. The horizontal dotted line represents the threshold for clinical significance on the Conners’-T (≥65).
As mentioned above, there was a statistically significant interaction between COMT and EC (F (3.8, 954.23) = 4.46; p = .002). Power calculation indicated that at n = 513, there was a 70% chance of rejecting the null hypothesis. Figure 2 shows that this effect stems from the fact that children homozygous for the Met allele display significantly lower scores compared to the Val heterozygous under baseline (Mean Met/Met = 65.96, SD = 12.56 vs. Mean Val/Met = 70.35, SD = 14.38; p = 0.004) and marginally under placebo (Mean Met/Met = 63.1, SD = 13.55 vs. Mean Val/Met = 66.18, SD = 15.46; p = 0.056) conditions.
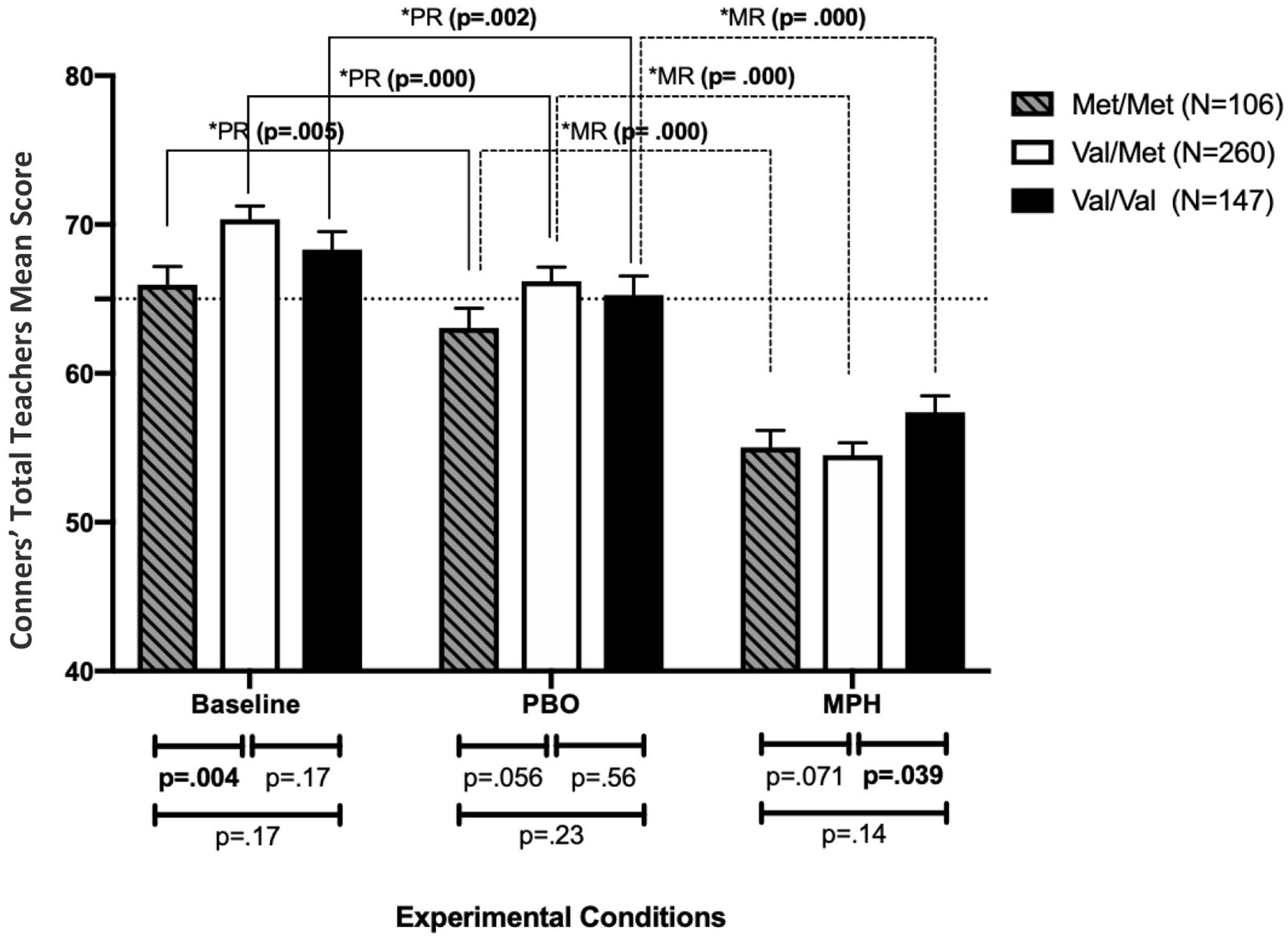
Conners’ Global Index Teachers (Conner’s-T ± SEM) scores separated according to child’s COMT genotype at methylphenidate (MPH), placebo, and baseline weeks. Error bars represent standard error of the mean. The horizontal dotted line represents the threshold for clinical significance on the Conner’s-T (≥65).
Although children with the Val/Val genotype showed higher scores under baseline and placebo (Mean baseline = 68.32, SD = 14.54; Mean placebo = 65.38, SD = 15.63) compared to Met/Met genotype group, these differences were not statistically significant (p > .05). However, a marginal significant difference between the Val/Val group and the Val/Met was detected under MPH, where the Val/Met group has a greater improvement on MPH (Mean Val/Met = 54.5, SD = 13.36 vs. Mean Val/Val = 57.3, SD = 13.5; p = .039).
COMT X DRD3 X EC Interaction
When children were stratified according to their DRD3 genotypes within each of the three COMT genotypes, statistically significant and biologically meaningful effects of the DRD3 genotypes were detected. Power calculation indicated that with this sample size, there was a 60% chance of rejecting the null hypothesis.
Within the COMT Met/Met genotype, there were only two DRD3 genotypes (T/T and T/C; the C/C genotype was excluded because of low numbers; n = 9). As seen in Figure 3A, the two-genotype groups were statistically indistinguishable under the three EC (indicating similar behavioral scores under all conditions) (all p values > .05). However, patients with the T/T genotype have significantly higher scores under the baseline condition compared to the placebo condition (Mean Baseline = 67.19, SD = 12.8 vs. Mean Placebo = 61.66, SD = 13.71; p = .000). In contrast, patients with the C/T genotype had similar scores under both baseline and placebo conditions (Mean Baseline = 64.62, SD = 10.62 vs. Mean Placebo = 64.19, SD = 13.35; p = .86). In other words, carriers of T/T genotype have a better response to placebo compare to those with the C/T genotype. Both genotype groups had lower scores under the MPH condition (Mean MPH (C/T) = 55.4, SD = 11.01 vs. Mean MPH (T/T) = 54.5, SD = 12.2) compared to their scores under placebo (p = 0.000); indicating a significant response to active medication.
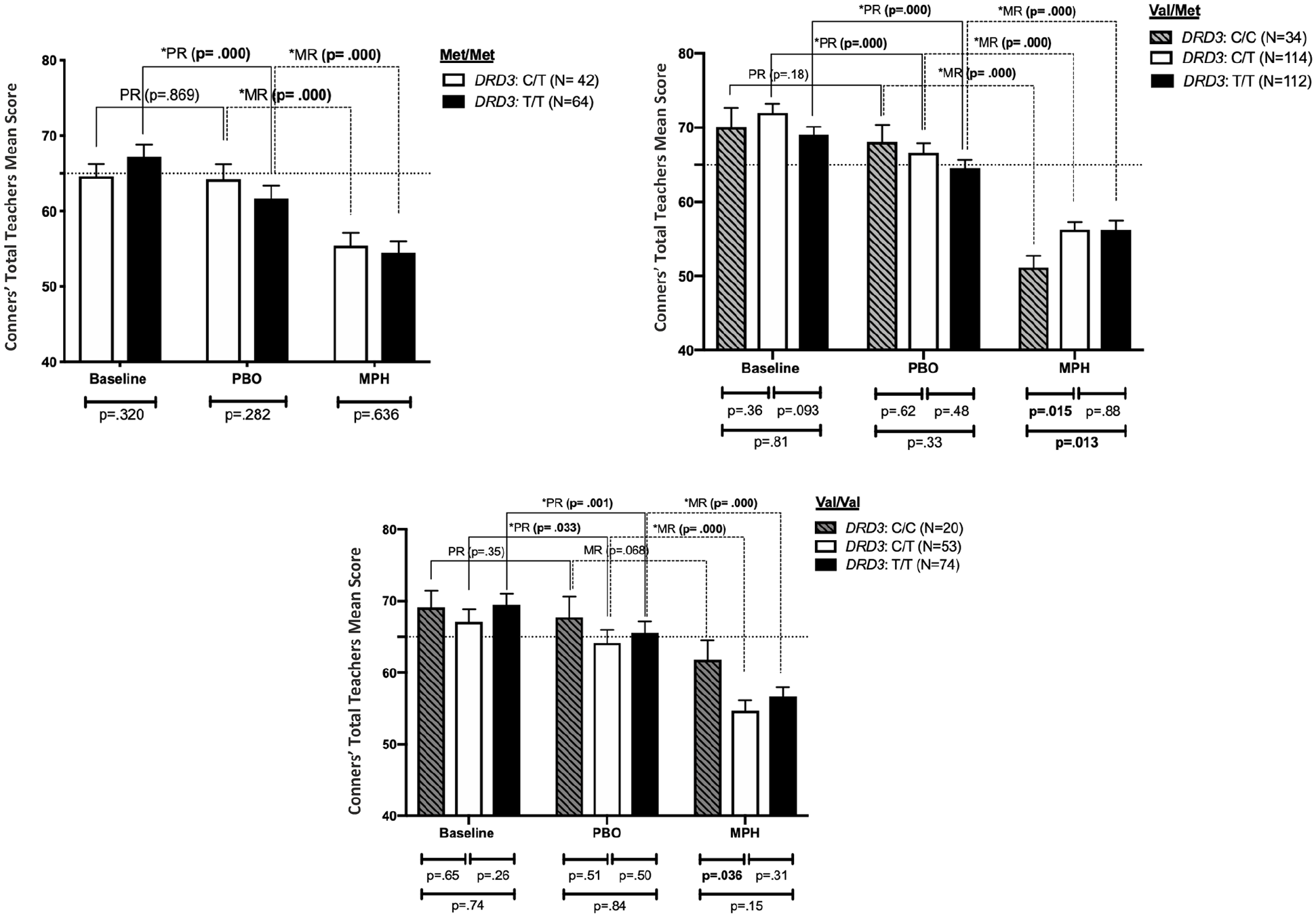
Conners’ Global Index Teachers (Conner’s-T, mean ± SEM) scores separated according to child’s DRD3 genotypes within each of the three COMT genotypes at methylphenidate (MPH), placebo, and baseline weeks. Error bars represent standard error of the mean. The horizontal dotted line represents the threshold for clinical significance on the Conner’s-T (≥65).
Within the COMT Val/Met genotype (Figure 3B), the three genotype groups of DRD3 (i.e., C/C, C/T, T/T) were statistically indistinguishable at baseline and placebo evaluations (all p values > .05); indicating similar behavioral scores under these two EC. However, statistically different behavioral scores were observed under MPH. The C/C genotype has significantly lower scores (Mean = 51.15, SD = 9.23) compared to the C/T (Mean = 56.25, SD = 10.74; p = .015) and the T/T (Mean = 56.23, SD = 12.79; p = .013) genotype groups. By looking at the level of improvement under each EC for each genotype group, we observed that children with T/T and C/T genotypes displayed significantly higher scores under the baseline condition (Mean Baseline (T/T) = 69.05, SD = 11.22; Mean Baseline (T/T) = 72.01, SD = 12.54) compared to the placebo condition (Mean Placebo (T/T) = 64.52, SD = 12.15; Mean Placebo (C/T) = 66.61, SD = 13.57; both p-values = .000). In contrast, patients with the C/C genotype had similar scores under both baseline and placebo conditions (Mean Baseline = 70.09, SD = 14.9 vs. Mean Placebo = 68.06, SD = 13.57; p = 0.18) (i.e., they are not showing any improvement after administering placebo). All genotype groups had lower scores under the MPH condition compared to both baseline and placebo conditions (p-values = .000).
Within the COMT Val/Val genotype group (Figure 3C), we observed that patients with the three genotyping groups of DRD3 were statistically indistinguishable in the baseline and placebo conditions (all p values > .05). Nonetheless, carriers of C/T tend to marginally have a better score under MPH compared to the carriers of T/T (p = .036). Examining the levels of response to placebo and MPH indicates that carriers of C/C did not show a significant improvement on placebo. In other words, their behavioral scores didn’t differ significantly under placebo compare to the baseline (Mean Baseline = 69.1, SD = 10.65; Mean Placebo = 67.75, SD = 12.8; p = .35). Interestingly, this group didn’t show a significant improvement on MPH compared to placebo (Mean MPH = 61.8, SD = 12.25; p = .068). On the other hand, the other two genotype groups (i.e., C/T and T/T) had significantly lower scores under the placebo condition (Mean Placebo (C/T) = 64.17, SD = 13.32; Mean Placebo (T/T) = 65.53, SD = 13.81) compared to the baseline condition (Mean Baseline (C/T) = 67.09, SD = 12.74; Mean Baseline (T/T) = 69.5, SD = 12.8; both p-values = .000). They also had significantly lower scores under the MPH condition compared to placebo (p-values = .000) (Table 2A).
Effect of DRD3 and COMT Polymorphisms on Conners’ Parents Scores
As with teachers, there was a highly significant effect of the EC (F (2,722) = 255.41; p = .000); but no statistically significant interaction or main effects of the genotypes (all p values > .05, Table 2B).
Discussion
One of the major problems with investigating the genetic factors underlying the etiology or modulators of treatment response in children with ADHD, has been the complexity of the disorder. While the most recent ADHD GWAS identified 12 loci that showed significant association with the disorder, these results “only capture a tiny fraction of common variant risk for ADHD” (Demontis et al., 2019). While the polygenic nature of this disorder could be challenging, testing genetic loci without taking into consideration their possible interaction with other genetic variants or with environmental factors might be unyielding, particularly in disorders manifested by behavioral outputs of complex neural systems where robustness, embodied through several redundant biological and neural systems, is the rule rather than the exception (Girard et al., 2015). While gene-by-gene and gene-by-environment interactions have usually been tested in separate studies, the novelty of this study is testing gene (COMT) by gene (DRD3) by environment (i.e., MPH, placebo, baseline) interactions, where the environment is conceived as controlled induction of synaptic dopamine using pharmacological probes (MPH and placebo). The experimental manipulation of the environment by pharmacological probes allowed us to approximate this synaptic level of dopamine, which contrasts with all previous studies where the environment was measured retrospectively and may have multiple effects on the nervous system, including on dopamine levels. The large sample size used in this study is also an important improvement compared to previous gene by gene and gene by environment interaction studies in ADHD.
As expected, there was a highly significant effect of the EC according to parents’ and teachers’ observation. This effect validates our assumption of studying the effect of DA-related candidate genes on ADHD-relevant behavior under different tones of synaptic DA: highest under MPH, intermediate under placebo, and lowest at baseline. Under the premise that behavioral outputs would be abnormal only when extreme genetic and environmental conditions are met, we predicted that the genetic effects of the two candidate genes will be revealed mostly when the children are under the least favorable conditions “in terms of lowest putative dopamine levels,” that is, at baseline or while they are on placebo.
This study has three main findings, which are partially in line with these expectations. First, a significant interaction between COMT Val/Met functional polymorphism and EC in modulating ADHD behaviors as assessed by teachers. The COMT Val/Val and Val/Met have the highest Conners’-T scores (indicating worst behavior) compared to Met/Met group particularly at baseline and under placebo, the two conditions that are associated with the lowest and second lowest levels of dopamine respectively. Interestingly, the difference between the COMT genotype groups almost disappeared on MPH, indicating that under this condition the genotype differences might not have any effect on behaviors. This result is in line with our expectations and illustrates the pertinence of this research approach.
Second, after stratifying subjects based on their COMT genotypes, an effect of the DRD3 functional Ser9Gly polymorphism on Conners’ teachers scores was revealed, although the results were complex and partially in line with our initial expectations. First, under the Met/Met and Val/Val COMT genotypes and after correcting for multiple testing, there were no statistically significant differences between the DRD3 Ser9Gly genotypes with regard to Conners’ Teachers evaluations, suggesting that when dopamine is too low or too high as imposed by the COMT Val/Val and Met/Met variants respectively, the effect of the DRD3 genotypes is obliterated. However, under intermediate synaptic dopamine levels, as imposed by the COMT Val/Met genotype, the effects of the DRD3 genotype are revealed. Although several contrast between DRD3 genotypes were significant under various EC, the most significant contrast was observed between patients with the C/C genotype and patients with the T/T and C/T genotypes under the MPH condition, with the C/C carriers having significantly lower scores than the other two groups. The higher binding affinity for dopamine, as suggested by cellular models expressing the C allele (Lundstrom & Turpin, 1996), could explain these findings.
The third important result is the contrast between the finding of genetic/behavioral associations when the behaviors are assessed by teachers, and the complete absence of any association when the behaviors are assessed by parents even though the scales used by parents and teachers assess the same behavioral dimensions. While this might be surprising at a first glance, there is important literature suggesting that the determinants of a child’s behaviors in different environments could be very different. Indeed, it has been shown, for example, that the heritability of ADHD as assessed by parents and by teachers is different (Martin et al., 2002; Sherman et al., 1997; Thapar et al., 2006). Further, parents evaluate the behavior of the child usually at the end of the day and during weekends when the expectations of parents and the work load demands on the child are very different from school settings. Thus, it is not surprising to see such differences.
Strengths and Limitations of the Study
Taken together, the present study suggests that, while it is possible to uncover association between ADHD relevant behaviors and genetic variants, these associations may be very complex and need stringent experimental conditions and large sample sizes to be reliably pinned down. The major strength of this study is its use of pharmacological probes to experimentally “control” the level of synaptic DA in order to reveal the effect of genetic variations in DA-related genes on ADHD relevant behavior. It is also to be noted that although this study can be informative as a pharmacogenetic study (i.e., understanding the effect of genes on therapeutic variability to MPH), it has not been designed primarily for this purpose. Indeed, in this study, MPH and placebo have been used as pharmacological probes to study dynamically ADHD relevant behavior under different conditions. In this context, we used relatively low levels of MPH (0.5 mg/kg/day) and short period of treatment (1 week). The other main strength of this study is its double-blind design, where evaluators and researchers did not know what treatment the child is receiving. This might result in more reliable evaluation of behaviors.
Some limitations of this study need to be kept in mind. While, to our knowledge, this is the largest study in ADHD investigating gene-by-gene interaction and included 619 children evaluated prospectively with a very stringent evaluation protocol, some of the gene-by-gene stratum contained relatively few subjects (one stratum with nine subjects was excluded from the analysis and the smallest stratum retained in the analysis contained 23 subjects). Early evidence from our group suggests a stronger effect of DRD3 in boys with ADHD (Fageera et al., 2018). However, due to the sample size, we may have insufficient statistical power to stratify the sample by gender and study the gene-gene interaction. Also, while the results with regard the COMT interaction with the EC seem robust, the three-way interaction between the COMT, DRD3, and the EC need to be considered with caution and needs further confirmation in a larger sample size. In addition, Ser9Gly is the only polymorphism that was shown to affect the protein structure of DRD3 (Lannfelt, 1992); yet, other unexplored polymorphisms could play a role in modulating the behaviors of children with ADHD that need further exploration. Further, the fact that these two markers didn’t reach the significance threshold in the latest ADHD GWAS study, could be a reflection of the extreme heterogeneity of the samples and the measures used in these very large international studies.
Conclusion
This study uses a combination of methodological tools (pharmacological probes, large sample size, and selecting genes consecutively implicated in synaptic transmission pathway of DA activity), that might be essential to better understand the role of candidate genes in complex behaviors. In fact, studying candidate genes with a priori knowledge, could make them amenable to clinical translation. Nevertheless, to further confirm these findings and because of the complexity of behavioural genetics, additional independent studies, with different ethnic populations, more genes, multiple other polymorphisms in these loci, and haplotype-tagging strategies are warranted. These studies will be informative only if large sample sizes and rigorous phenotype assessments are implemented.
Supplemental Material
Figure_S1 – Supplemental material for COMT by DRD3 Epistatic Interaction in Modulating Behaviors in Children with ADHD: A Pharmaco-Dynamic Behavioral Approach
Supplemental material, Figure_S1 for COMT by DRD3 Epistatic Interaction in Modulating Behaviors in Children with ADHD: A Pharmaco-Dynamic Behavioral Approach by Weam Fageera, Natalie Grizenko, Sarojini M. Sengupta, Norbert Schmitz and Ridha Joober in Journal of Attention Disorders
Supplemental Material
Figure_S2 – Supplemental material for COMT by DRD3 Epistatic Interaction in Modulating Behaviors in Children with ADHD: A Pharmaco-Dynamic Behavioral Approach
Supplemental material, Figure_S2 for COMT by DRD3 Epistatic Interaction in Modulating Behaviors in Children with ADHD: A Pharmaco-Dynamic Behavioral Approach by Weam Fageera, Natalie Grizenko, Sarojini M. Sengupta, Norbert Schmitz and Ridha Joober in Journal of Attention Disorders
Supplemental Material
Supplementary_material – Supplemental material for COMT by DRD3 Epistatic Interaction in Modulating Behaviors in Children with ADHD: A Pharmaco-Dynamic Behavioral Approach
Supplemental material, Supplementary_material for COMT by DRD3 Epistatic Interaction in Modulating Behaviors in Children with ADHD: A Pharmaco-Dynamic Behavioral Approach by Weam Fageera, Natalie Grizenko, Sarojini M. Sengupta, Norbert Schmitz and Ridha Joober in Journal of Attention Disorders
Footnotes
Acknowledgements
We thank present members of the ADHD group (Nellie Fotopoulos, Sana Miled, Phuong-Thao Nguyen, Jacqueline Richard, Mira Thakur, Adam Torkamanzehi), and past members (Johanne Bellingham, Rosherrie DeGuzman, Zia Choudhry, Parvin Jinnah, Matthew Lebaron, Nissa Lebaron, Nicole Pawliuk, Anna Polotskaia, Sandra Robinson, Marina Ter Stepanian, Geeta A. Thakur, Nadia Zeramdini, Darya Naumova), for technical and clinical assistance.
Author Contributions
Conceived and designed the experiments: RJ and NG. Analyzed the data: WF. Performed the literature search: WF and SS. Wrote the paper: WF and RJ. Provided critical revision of the manuscript: NS, NG, and SS.
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: Dr. S.M. Sengupta, and W. Fageera report no biomedical financial interests or potential conflicts of interest. Dr. Natalie Grizenko reports receiving research funding from CIHR and is a member of the advisory board for Purdue and Shire. Dr. Ridha Joober reports having received research funding from CIHR. He is on the advisory boards and speakers’ bureau of Pfizer, Janssen Ortho, BMS, Sunovion, Otsuka, Lundbeck, Purdue and Myelin. He has received grant funding from them and from AstraZeneca and HLS. He has received honoraria from Janssen Canada, Shire, Lundbeck, Otsuka, Pfizer and from Purdue for CME presentations and royalties for Henry Stewart talks.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: WF holds a PhD scholarship from the Ministry of Education of Saudi Arabia. This work was supported in part by grants from the Fonds de la recherche en santé du Québec (FRSQ#6690) and the Canadian Institutes of Health Research (MOP#53309, MOP# 84537, MOP# 126023) to RJ and NG.
Supplemental Material
Supplemental material for this article is available online.
Author Biographies
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.