
Abstract
What we have attended to in the past, as well as the stimulus context associated with past motor responses, have a strong impact on our current behavior. These influences have been investigated through inter-trial priming effects in visual search and sequence effects in action control, respectively. These two research fields are strongly complementary at the theoretical level and show striking similarities in their experimental-task structure, analyses, and results. Yet, they have developed largely separately. Here, we claim that such fragmentation impedes progress in these two research strands and highlight the potential benefits of intensifying crosstalk between visual search and action control in future research by exploiting the existing structural similarities with regard to sequence effects. We first discuss the main phenomena and theoretical explanations in each field, while emphasizing the similarities and differences between them. Then, we illustrate how the two fields could integrate each other’s insights—namely, how visual-search research could draw on the action-control literature to clarify the role of retrieval in selection and how action-control research could draw on the visual-search literature to explain response-related processes in more complex environments. We argue that combining the two research traditions is necessary for a coherent account of search-for-action behavior.
Research in cognitive psychology is scattered across paradigms. Of course, experimental paradigms are used by researchers as “windows to the mind”—they are tools to unravel the mysteries of cognitive processing. Still, there is an abundance of explanatory frameworks tailored to specific paradigms or phenomena. As a consequence, research traditions have evolved that more or less ignore findings and ideas from related fields. This issue has contributed to the fragmentation in cognitive psychology and impedes scientific progress (Balietti et al., 2015). There is an urgent need to integrate and connect different fields of research (Eronen & Bringmann, 2021) that investigate phenomena that are tightly related and might de facto share the same underlying cognitive mechanisms.
In this paper, we attempt to build a bridge between two hitherto largely independent research strands: inter-trial priming in visual search (e.g., Maljkovic & Nakayama, 1994, 1996) and sequence effects in action control (e.g., Frings et al., 2020). These research fields differ in their focus. Roughly speaking, inter-trial priming in visual-search research is concerned withbrav stimulus-selection processes and how these are influenced by stimulus repetition (mainly repetition of target or distractor locations or of any of their features). Responses are typically seen only as the means to collect a behavioral measure of task performance (RT or accuracy), and response-related processes are mainly regarded as potential confounding factors—the main research focus being on effects influencing the guidance of attention (e.g., Asgeirsson et al., 2013; Kristjánsson & Ásgeirsson, 2019; but see, e.g., Lamy et al., 2010; Geyer et al., 2007; Töllner et al., 2008). By contrast, sequential effects in action-control research are concerned with response-related processes: planning and selecting a response, response execution, and response monitoring. As stimulus-selection processes are not within the scope of this research, displays typically involve one or very few stimuli and the task does not require search (but see Frings & Rothermund, 2011; Moeller & Frings, 2014).
The lack of crosstalk between these two research fields is unfortunate for several reasons. First, it is becoming increasingly recognized that perception and action are closely intertwined (as foreshadowed by discussions by, e.g., Neumann, 1990; Styles & Allport, 1986). In fact, researchers have argued that perception and action are at least partially represented in the same neural circuits (common coding, Prinz, 1997; Hommel, 2004; Rizzolatti & Craighero, 2004). Thus, they should not be treated as separate cognitive modules. For instance, repeatedly responding to a stimulus changes its perception and selection (Pastötter & Frings, 2018) while conversely, selection demands impact response planning (Laub et al., 2018; Schmalbrock et al., 2022). Second, failures to concomitantly consider both perceptual and action-related processes have led to erronous interpretations in the respective literature (e.g., the claim that stimulus salience has no impact on action control, Schmalbrock et al., 2022, or the overestimated role of attention in target-repetition effects, e.g., Lamy et al., 2010). More generally, as we argue in the second part of this paper, visual-search and action-control research complement each other: we search for objects in order to act upon them, and the objects we act upon typically require search because they appear among irrelevant objects. Therefore, if the ultimate goal is to understand behavior in real world settings, the two perspectives must be combined.
Focusing on sequence effects provides a particularly promising starting point for this endeavor, because—as will become clear—we can exploit existing structural similarities in designs and draw upon a large body of literature from the two research traditions. Indeed, it is striking that despite the differences in focus, in both research fields, the experimental tasks have exactly the same structure (Figure 1): participants are presented with a display on each trial and are required to discriminate a feature of the target, such as its identity or color, typically by making a manual response.
1
In addition, in both, trial sequences (trial n-1 and trial n) are analyzed and the aspects of the trials for which repetitions are examined (e.g., response, target feature, target location, and distractor feature) vary randomly and orthogonally on each trial. Therefore, there are no contingencies between response repetitions and stimulus repetitions. The main difference is that action-control tasks do not require search and focus on the interaction between response repetitions and stimulus or task repetitions, whereas inter-trial priming tasks in the visual-search literature require search and the primary focus is on the main effect of stimulus repetition (e.g., of the target’s location). Performance in that literature is almost always found to be faster on stimulus-repetition than on stimulus-changed trials. The visual-search studies that analyzed the Structural similarity between visual-search and action-control experiments with regard to (1) inter-trial effects, (2) the analysis strategy, and (3) the typical results. (1) In both research fields, participants respond to sequences of trials, and both response- and stimulus-related information can repeat. In the search-task example, participants search for a target with the unique color and respond to the orientation of the T inside it. In the action-control task example (here, a distractor-response-binding task), participants respond to the identity of the central letter, while the flanking letters serve as distractors. In both examples, the response to the target changed between trial n-1 and trial n, while the stimulus-related feature (the target color or the distractor identity) repeated. (2) The crucial analysis is how these repetitions/changes affect performance on trial n. (3) As illustrated in the two specific experiments displayed here (Frings et al., 2007; Lamy et al., 2011), a similar interaction is typically found between response repetition and stimulus repetition. The original data are reproduced with permission.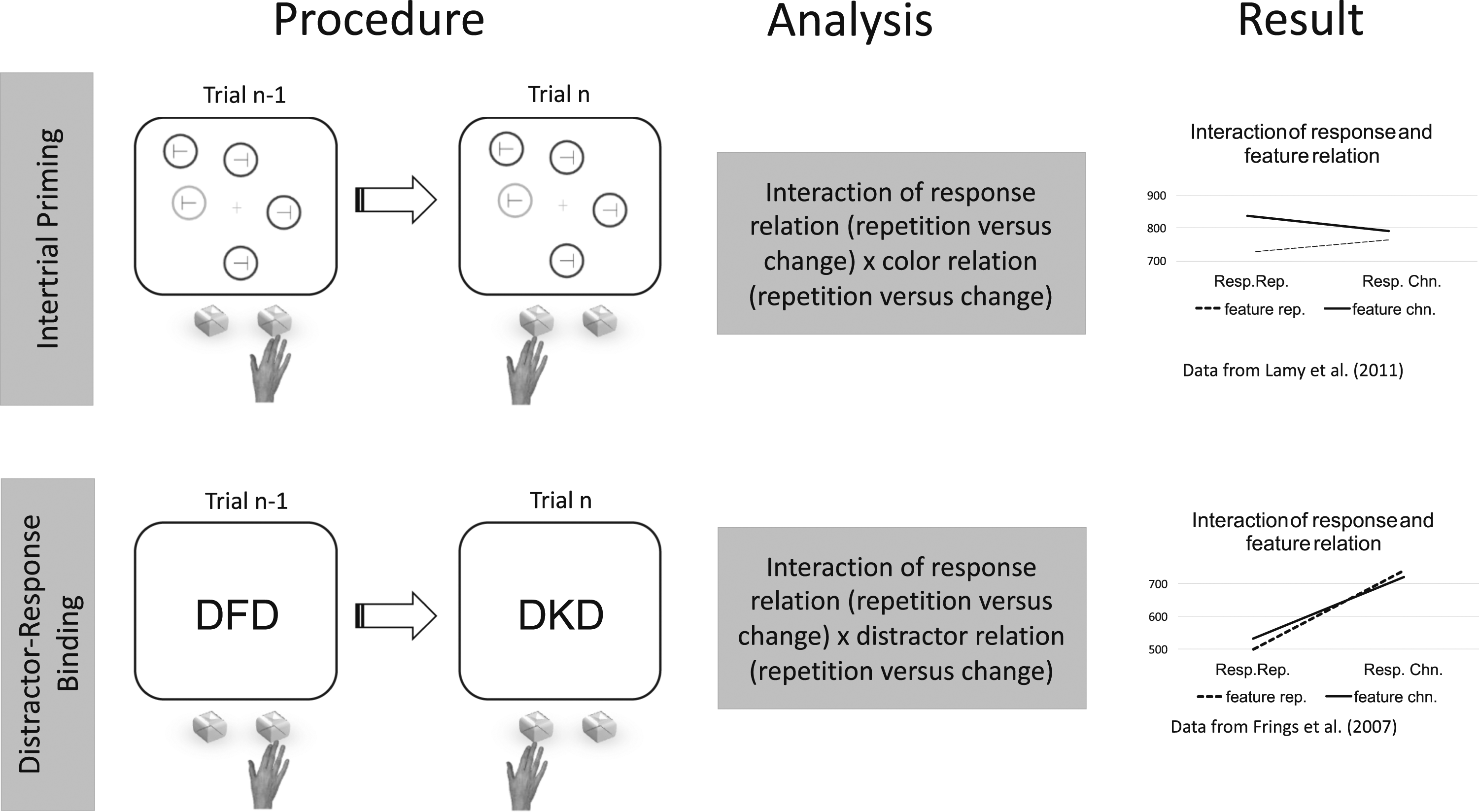
Against the background of these commonalities in experimental procedures, analyses, and results, it is striking how little crosstalk there is between these research fields. In this paper, we first highlight the common structure of the tasks, while also discussing the differences in the procedures used (Part 1). Then, we relate the theoretical explanations proposed in each field and point to urgent questions to be investigated in future research (Part 2). This endeavor holds the potential to advance both fields. On the one hand, applying insights from the action-control literature to visual-search data will allow researchers to better disentangle attention-related and response-related sources of inter-trial effects and thereby advance our understanding of memory influences on search guidance—a burgeoning and highly influential subfield in attention research (Anderson et al., 2021; Awh et al., 2012; Luck et al., 2021; Ramgir & Lamy, 2021). On the other hand, integrating aspects of visual-search designs, high display complexity in particular, will move action-control research closer to the conditions prevailing in the real world, where—in contrast to typical action-control laboratory studies—a stimulus is hardly ever presented in isolation.
Sequential Structure in Visual-Search and Action-Control Experiments
Inter-trial Priming Effects in Visual Search
Our information processing capacity is limited, and our motor behavior can only be directed to a limited subset of stimuli. Selective attention refers to the collection of mechanisms that allow us to prioritize some stimuli in our environment over others. In the lab, mechanisms of attentional selection are often studied using visual-search tasks.
In a typical experiment, participants are instructed to search for a target and to respond to some aspect of that target, on each trial. For instance, the target might be defined as the uniquely colored object (or singleton) among homogeneous nontargets (e.g., the only red diamond among green diamonds or vice-versa, Bravo & Nakayama, 1992) and participants are instructed to make some discrimination with regard to this target (e.g., determine whether the left or right corner of the target diamond is chipped). All stimuli, the target and distractors alike, are equally likely to have each of the possible reported features, so that participants cannot find the target based on its reported feature. In addition, the values of the reported feature (chipped corner on the left or right) are chosen so that it is necessary to focus attention on the target in order to identify the reported feature and select the correct response. The measure of interest is how long it takes for participants to respond (reaction times) and how often they are correct (accuracy) as a function of a variety of independent variables thought to affect search difficulty.
Many effects on search time have traditionally been interpreted in terms of how efficient attention is allocated toward the target, either in terms of how fast distractors are rejected (e.g., Duncan & Humphreys, 1989) or, from another theoretical perspective, in terms of how strongly attention is guided toward the target (Wolfe, 2021). Specifically, much research has focused on the interplay between aspects of the current stimulus display (stimulus-driven guidance) and the intentions of the observer (goal-directed guidance). However, other studies showed that past experience (also referred to as selection history), which cannot be readily assimilated to either stimulus-driven or goal-directed guidance, also determines search performance. In particular, unpredictable repetitions from previous trials were shown to speed search considerably.
Feature, Location and Dimension Inter-trial Priming
In a seminal series of studies, Maljkovic and Nakayama (1994; 1996; 2000) demonstrated that in a task similar to the example described in Figure 1, search was much faster when the target’s color or its location happened to repeat on successive trials (inter-trial priming of feature and location, respectively), with independent contributions of the two repetition types. In addition, when they manipulated repetition of the distractors’ color separately, they found it to also speed search (distractor-repetition priming). Later research confirmed and extended this finding by showing that when roles are reversed (i.e., when the target takes on the feature of a previous distractor or vice-versa, Kristjansson & Driver, 2008; Lamy et al., 2008a), performance is impaired. Remarkably, the target location, its color, and the distractors’ color on any given trial influenced performance on the next 5 to 8 trials. By contrast, repetition of the reported feature (i.e., repetition of which corner of the uniquely colored diamond was chipped) had no impact on performance.
Around the same time, Müller and colleagues (Found & Müller, 1996; see also Müller et al., 1995; Müller et al., 2003) observed a structurally similar effect: participants searched for a singleton target that could stand out from the surrounding nontargets by either its unique color or its unique orientation; the specific feature (e.g., red vs. blue) as well as the feature dimension (color vs. orientation) unpredictably changed across trials, while the surrounding nontargets remained constant. Results indicated a strong performance advantage when the target dimension repeated relative to when it changed. The effect of feature repetition was much weaker and was reliably present for color, thus replicating Maljkovic and Nakayama’s (1994) findings, but not for orientation (for further evidence that salience computations for color are special, see Bravo & Nakayama, 1992; Nothdurft, 1993; for a demonstration that inter-trial priming of orientation is special, see Lamy & Kristjánsson, 2013).
Other related yet less widely investigated repetition effects were reported in visual search. For instance, search for a target that happens to be a singleton on the current trial was found to be faster when the target on the previous trial had also been a singleton on the same dimension rather than when it had not been a singleton (singleton priming, Lamy et al., 2008a; Lamy et al., 2006; see also Won et al., 2019). Likewise, when target-absent trials are interspersed among target-present trials similar to Maljkovic and Nakayama’s (1994) classical pop-out trials, target discrimination performance is better when distractors’ color repeats than when it changes from a target-absent to a target-present trial (distractor preview effect; Ariga & Kawahara, 2004; Goolsby & Suzuki, 2001; LlerasKawahara et al., 2008).
Weighting Accounts of Inter-trial Priming Effects
The canonical accounts of these inter-trial priming effects in visual search can be collectively referred to as weighting accounts (e.g., Chun & Nakayama, 2000, for feature and location priming; Krummenacher & Müller, 2012 for dimension priming; Lamy et al., 2008b for singleton priming; and Wan & Lleras, 2010, for the distractor preview effect). In a nutshell, these accounts posit that an implicit memory trace encoding the properties of the target and distractors is laid on every trial and that this memory trace influences the deployment of attention in subsequent searches by increasing the priority weight associated with the properties of the target and decreasing the priority weight associated with the properties of the distractors.
For instance, Maljkovic and Nakayama (1994; 1996) suggested that only the properties that are important for selection (i.e., the target’s and distractors’ features on the target-defining dimension, as well as their locations) are encoded in these memory traces, but reported features, as well as task-irrelevant features, are not (see Levinthal & Lleras, 2008 for a similar claim with regard to the distractor-preview effect). However, later studies showed that repetition of both the reported feature (e.g., Kristjánsson, 2006) and irrelevant features of the target (e.g., Kristjánsson, 2006; McBride et al., 2009) can speed performance, suggesting that all the features of the selected target are encoded in the episodic memory trace and contribute to guide attention on subsequent trials (e.g., Lee et al., 2009; Olivers & Humphreys, 2003).
Along similar lines, Müller and colleagues suggested the Dimension Weighting Account (DWA; for reviews, see Krummenacher & Müller, 2012; Liesefeld et al., 2018; see also Liesefeld & Müller, 2021) to account for dimension-repetition benefits. They proposed that selecting a target that stands out on a given dimension on trial n increases the weight of that dimension, an increase that typically persists until (at least) the next trial (Allenmark et al., 2018). As a result, guidance toward the target on trial n +1 is enhanced if the target again stands out in the same rather than in a different dimension.
Episodic-Retrieval Accounts of Inter-trial Priming in Visual Search
According to weighting accounts, the mechanisms that underlie inter-trial priming effects operate in a forward manner: selection of a target at the expense of competing distractors on a given trial influences the priority weights accruing to similar stimuli on the next trials. Other accounts, however, referred to as episodic-retrieval accounts (e.g., Hillstrom, 2000; Huang et al., 2004; Huang & Pashler, 2005; Lamy et al., 2010; see also Thomson & Milliken, 2011, 2012, 2013) propose that these mechanisms operate backwards: on each trial, different aspects of the search trial are bound in one episode (e.g., Logan, 1988), object file (Kahneman et al., 1992), or event file (e.g., Hommel, 1998); under certain conditions, a search trial can retrieve the memory trace of a previous search trial and the degree of match between these successive search events influences performance, resulting in inter-trial priming effects.
However, these accounts vary widely with regard to the stage of processing at which retrieval is thought to influence search performance. On the one hand, some episodic-retrieval models of inter-trial priming in visual search posit that retrieval affects the speed of attentional selection. For instance, Hillstrom (2000) suggested that on each trial, a crude preattentive map of the features in the search display is created and triggers retrieval of traces for similar trials, which include the prioritization rules associated with these trials. If the relevant trace is retrieved before standard prioritization processing is completed, priorities on the current trial will be assigned to features according to the retrieved prioritization rules. Huang et al. (2004) also suggested that trace retrieval races with the search algorithm, but situated the process that is influenced by retrieved traces at a later stage of attentional selection. They proposed that after the target is located, its features (defining, response, and irrelevant features; see Liesefeld et al., 2018 in press) on the previous and current trial are compared. “When all the feature dimensions cohere … (all favoring the same judgment or all favoring a different judgment), the judgment is easy. On the other hand, inconsistency across dimensions will have a cost. When it arises, the system may need extra time to double check the status of the target” (p. 21).
Finally, other episodic-retrieval models acknowledge that inter-trial priming guides attention, but argue that in addition, episodic retrieval biases an even later stage, during which the motor response is selected. For instance, Lamy and colleagues (e.g., Lamy et al., 2010; Yashar & Lamy, 2011) argued that inter-trial priming of the target-defining feature reflects two separate mechanisms. On the one hand, selection of a target on a given trial is faster when its defining feature repeats from a previous trial, an effect that reflects the attentional component of inter-trial priming. On the other hand, when the current search trial is similar to the previous trial, participants are more likely to reactivate the motor response associated with the previous trial. Likewise, Hilchey et al. (2018; see also Gokce et al., 2014) suggested that after the target is selected, when its location repeats, the response recently associated with that location is rapidly retrieved and reactivated; however, they acknowledged that an additional mechanism, which may be considered attentional, is also at play in target-location priming. A similar dual-stage account of inter-trial priming was suggested with regard to dimension repetitions (e.g., Töllner et al., 2008).
Empirical research and theorizing on the role of episodic retrieval during visual search is rather limited. This scarcity contrasts with the profusion of studies that have examined inter-trial effects in action control, where episodic retrieval plays a prominent role, as described next.
Inter-trial Effects in Action Control
The ability to act according to an instructed action goal and the cognitive consequences thereof are typically analyzed in experimental paradigms that each focus on a specific aspect of an action. This research tradition employs a variety of quite distinct paradigms. We therefore first briefly describe the paradigms for which inter-trial effects were observed and how they relate to the control of action.
Task Switching
The task-switching paradigm was invented to study cognitive control/flexibility (needed to perform goaldirected behavior) in dynamic and changing environments with different action demands. In order to analyze task-switching costs typically two different tasks are presented in alternation (see Kiesel et al., 2010; Koch, Poljac, et al., 2018 for reviews). Task-switching costs refer to the performance decrement on trial n when the task changes from trial n – 1 relative to when it repeats. Some researchers have argued that while performing one task, participants inhibit the other task (inhibition account, Koch et al., 2010; Mayr & Keele, 2000; Schuch & Koch, 2003). Others have suggested that when the task changes, participants have to re-configure the task set to a specific stimulus-response (S-R) mapping, a process that takes time (re-configuration account, Rogers & Monsell, 1995; for a similar idea in visual search, see the Dimension Weighting Account introduced above). Yet others have linked switching costs to task cues that trigger an episodic-retrieval process (Koch & Allport, 2006). The retrieval includes features of the previous task, the previous response, and/or the previous action goal (Arrington et al., 2007; Logan & Bundesen, 2003). In the case of a task switch, the retrieved features do not match the features of the current trial, leading to a performance cost. Further evidence that task switching costs are related to processes of binding and retrieval is provided by the ubiquitous finding that task-switch costs depend on whether the response switches or repeats from trial n – 1 to trial n (e.g., Altmann, 2011; Hübner & Druey, 2006; Koch, Frings, & Schuch, 2018; Mayr & Kliegl, 2003; Rogers & Monsell, 1995).
Negative Priming
The negative-priming paradigm was invented to investigate inhibitory control of distracting stimuli that is exerted while selectively responding to a target (Tipper, 1985; Tipper et al., 1994; for a recent review Frings et al., 2015). In this task, a target and a distractor are presented on each trial. When the prime distractor from trial n – 1 serves as the probe target on trial n, performance costs are observed relative to a neutral condition, where the probe target stimulus was absent on trial n – 1. Two main accounts have been suggested to explain negative-priming effects. Inhibition models (Houghton & Tipper, 1994) propose that the perceptual representation of the prime distractor is inhibited on trial n – 1 and that such inhibition persists into trial n, impairing probe processing in the case of a distractor-to-target repetition. By contrast, episodic-retrieval theories (Neill, 1997) posit that during probe processing, the last episode containing the probe target is automatically retrieved and when the retrieved information is incompatible with the current information (in the case of a distractor-to-target repetition) interference emerges. More recent accounts explain negative priming in terms of event-file binding in the prime trial and event-file retrieval in the probe trial (Rothermund et al., 2005; see also Mayr & Buchner, 2006). So, while the retrieval process seems comparable to what Neill (19997) suggested, the idea here is that the whole prime episode, including the response, is retrieved.
Stimulus-Response-Binding Tasks
The S1R1-S2R2 paradigm (Hommel, 1998) and the distractor-response-binding task (Frings et al., 2007) were invented to investigate the impact of feature/stimulus repetitions on action control. In the S1R1-S2R2 task, an arbitrary response (R1) is executed while a stimulus (S1) is displayed. In the next display, a second stimulus (S2) is presented, which the participant has to process, because it determines the second response (R2). If aspects of the stimulus and/or the response repeat between S1R1 and S2R2, facilitation (in the case of complete repetitions) or interference (in the case of partial repetitions) is observed relative to complete changes. In the distractor-response-binding task, a distractor appears simultaneously with the target on each trial. The distractor either repeats or changes between two consecutive displays, and so does the response to the target. Distractor-repetition effects are modulated by whether or not the response repeats between prime and probe. The prevailing explanation for the pattern of results in both tasks is that features of the stimuli (S1 or the distractor) are integrated with response codes on trial n – 1 and that stimulus repetition on trial n retrieves the previous response, which facilitates responding when it matches the currently required response (on trial n; complete repetition) and interferes with it in case of a mismatch (partial repetition).
Gratton Effect
The Gratton effect (for a review see, e.g., Verguts & Notebaert, 2008) refers to the sequential modulation of congruency effects, that is, the finding that the congruency effect in trial n is typically smaller when trial n – 1 is incongruent than when it is congruent. Research on the Gratton effect is particularly concerned with cognitive control and more specifically with how conflicting information is handled. Ignoring incongruent information on trial n seems to be easier if trial n – 1 was also incongruent. The conflict-monitoring approach (Botvinick et al., 2001) suggests that detecting a conflict (as in an incongruent trial) increases cognitive control, leading to less interference when conflict is encountered again in the subsequent trial. However, others have observed that this pattern is particularly strong when the stimulus repeats from trial n – 1 to trial n, which opens the door to an alternative explanation in terms of binding and retrieval (e.g., Davelaar & Stevens, 2009; Mayr et al., 2003).
Action-Planning Tasks
Humans can prepare themselves for acting. The key question in action-planning studies is how the mental preparation of an action in a prime trial impacts the initiation of a related action in a probe trial. In a typical action-planning task, the prime response (on trial n – 1) is not executed, then the probe response (on trial n) is executed, and only thereafter the prime response is executed. This sequential prime-probe procedure has proven to be an ideal tool to reveal the underlying structure of action plans (Kunde et al., 2002; Rosenbaum & Kornblum, 1982; Stoet & Hommel, 1999). Specifically, feature binding during action planning on trial n – 1 can hinder or facilitate the initiation of feature-overlapping actions on trial n, though the specific conditions and reasons for such costs and benefits have remained elusive so far. Explanations in terms of feature binding and retrieval extend previous theorizing in terms of the code-occupation hypothesis (which assumes that features of the planned response are bound and less accessible for the prime response; Fournier et al., 2015).
The Binding and Retrieval in Action Control (BRAC) Framework
As is clear from the foregoing review, most action-control tasks share a prime-probe structure (or trial n – 1 to trial n structure) 2 and for each, an episodic-retrieval account has been invoked as an alternative to other paradigm-specific accounts. Recently, the Binding and Retrieval in Action Control (BRAC) framework has been suggested (Frings et al., 2020; Frings et al., 2015) as a common account for the vast corpus of findings on paradigm-specific action-control processes.
BRAC assumes that the different aspects of the stimulus configuration (stimulus, context, and cue), of the response (response goal, decision, and effector), and of the effect (sensory and affective) are integrated into a short-term memory entry that is labeled “event-file” in line with the Theory of Event Coding (TEC; Hommel, 2004). In addition, BRAC assumes that upon repetition of any feature of an event file, the event file containing this feature is retrieved and thereby can modulate current actions (i.e., the building of the current event file).
Binding and retrieval are treated as separable processes in the BRAC framework and BRAC describes action control in terms of dynamic event-file management (integration/binding of features into event files and the retrieval of event files). In each of the abovementioned action-control tasks, event-file binding happens during trial n – 1 (i.e., the prime trial) and if a feature of the event repeats on trial n (i.e., the probe trial), retrieval starts. The typical so-called “S-R binding effects” or “action effects” that emerge in these tasks are according to BRAC always a compound effect of a binding proper plus retrieval.
In addition, BRAC assumes that both top-down and bottom-up control exert their influences separately on binding and retrieval. For instance, top-down control mechanisms can modulate the binding process (e.g., features receiving much attention might be more likely to become integrated into event files) and/or the retrieval process (e.g., features that are ignored might be less effective retrieval cues). Likewise, they can influence binding and retrieval by fostering different levels of semantic representation (e.g., task rules, framing, mind sets, speed/accuracy tradeoffs, and instruction-based effects). In addition, some findings suggest “bottom-up control” of binding and retrieval, that is, modulations due to stimulus contingencies (Giesen & Rothermund, 2015), emotional stimuli (Waszak & Pholulamdeth, 2009), and perceptual configurations (as a result of Gestalt mechanisms; Laub et al., 2018; Frings & Rothermund, 2017) reflect influences on binding and retrieval processes by variables not originating in the observer. Location also influences binding and retrieval as repetitions versus changes of the locations of the stimuli to which one responds influence whether features and responses are bound in a binary fashion (when location repeats) or whether all stimulus features are integrated into a single object and this feature compound is bound to the response (when location changes; Singh & Frings, 2020).
Taken together, BRAC can potentially integrate paradigm-specific findings on sequential-action effects in terms of dynamic event-file management (Beste et al., 2023). We now go a step further and link the sequential effects in visual search and BRAC-centered sequential effects in action control in the next part of our article.
Theoretical Commonalities and Future Directions
Visual-search and action-control theorizing is typically based on classic sequential information processing models, in that it assumes a sequential processing stream broadly consisting of a selection, identification and response stages (see Figure 2). Finer-grained subdivisions of the processing stream have been suggested (e.g., Eimer, 2014; Pashler, 1998; Zehetleitner et al., 2012). For instance, Zehetleitner et al. (2012) also posited three sequential stages and divided each into two substages: the search stage includes attentional selection, followed by identification of the target-defining feature; the discrimination stage includes identification of the response-defining feature followed by response selection; finally, the response stage includes response planning followed by response execution. Processing stages and focus of visual-search and action-control studies. These processing stages are compatible with sequential information-processing models suggested by other authors (e.g., Eimer, 2014; Pashler, 1998; Zehetleitner et al., 2012).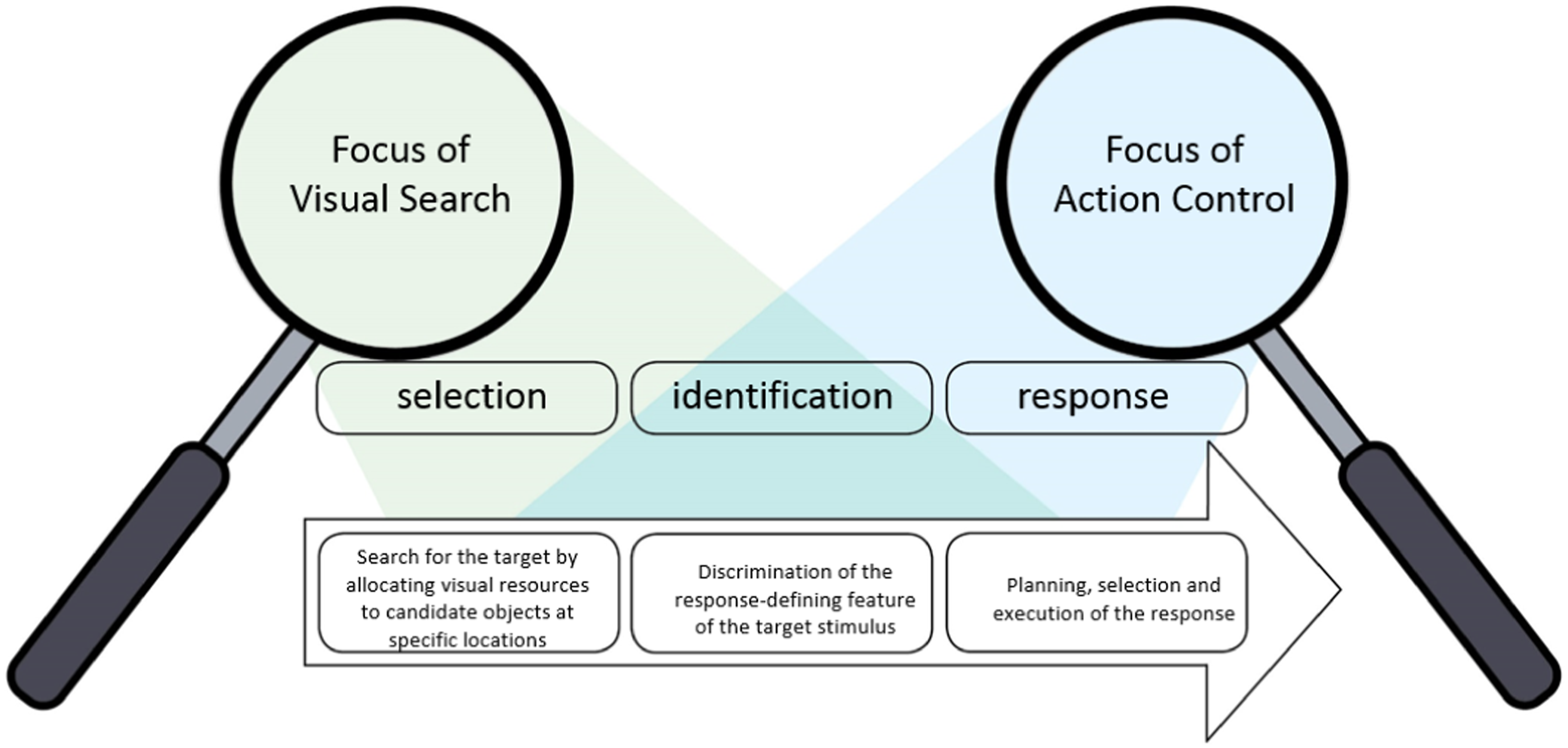
While it is unlikely that these stages proceed in a strictly serial order, the standard tasks used in visual search and action control can be well described by such sequential models. An important observation for the present purposes, however, is that the two research traditions have focused on different stages. Visual-search research is mainly concerned with the selection stage and only marginally with the identification stage, while action-control research is mainly concerned with the identification and response stages. However, as explained in the introduction, since the real-life behavior that each field aims at understanding, ultimately, is search for action, it is clear that all three stages need to be integrated. Such integration becomes even more critical if one acknowledges that not only do early stages influence processing in late stages but also late stages of previous search episodes can influence processing in early stages.
We therefore suggest that visual-search studies should integrate findings on response selection and planning from action-control studies. In particular, insights on the role of retrieval, which is a core explanatory construct in the action-control literature, should motivate further research drawing on the findings and methods of the action-control literature. Indeed, while all three processing stages are typically passed through when a new visual scene is encountered, retrieval from previous event files may provide shortcuts or modulate how each of these processes unfolds, in ways that are often not intended by the researchers (e.g., Henson et al., 2014). Likewise, we suggest that action-control research should integrate findings and theorizing from visual-search studies on selection. In particular, it is noteworthy that this research has typically used displays that are so simple (sometimes consisting of just a single letter) that attentional selection as typically examined via visual-search paradigms plays no significant role for task performance—a state of affairs that has led action-control researchers to neglect the selection stage. It is thus imperative to determine whether the conclusions drawn from the extant action-control literature generalize to more complex displays.
In the next sections, we review the few visual-search studies that have addressed the role of episodic retrieval in inter-trial priming as well as the few action-control studies that have employed more complex displays in sequential effects and point to the issues most in need of future research.
The Role of Episodic Retrieval for Inter-trial Priming Effects in Visual Search
While research on inter-trial priming in visual search has mainly focused on its attentional component—thereby giving the primary impetus for the notion that not only stimulus salience and goals but also selection history, guide attention (e.g., Luck et al., 2020)—it has largely neglected the role of episodic retrieval. Retrieval accounts have typically been lumped together into the camp that challenges weighting accounts of inter-trial priming in visual search, despite substantial dissimilarities between them. As a result, little effort has been expended to test the different episodic-retrieval accounts against each other. In addition, retrieval accounts that emphasize the stage of motor-response selection have been discussed separately for feature, location-, and dimension-repetition effects, with no overarching model accounting for the findings from these three lines of research.
What insights has this research yielded so far? Evidence for the idea that inter-trial priming reflects retrieval of prioritization rules is scarce: the idea entirely relies on the finding that in a conjunction-search task where, on each trial, a cue indicated the target-defining features of the upcoming target, target-feature repetitions sped search but did not reduce search slopes (Hillstrom, 2000)—a finding that is open to many alternative accounts (e.g., Amunts et al., 2014; Ramgir & Lamy, 2021; Wolfe et al., 2022).
Likewise, the claim that inter-trial priming speeds the decision of whether a candidate target is, in fact, the target, also relies on the findings from a single study (Huang et al., 2004). The authors found that the benefit of repeating the target-defining feature was strongly modulated by repetition of its reported feature as well as by repetition of an irrelevant feature of the target. They concluded that selection of the target is slowed when repetition from the previous trial is partial relative to a complete repetition or change (nicely fitting the standard explanation from action-control approaches). However, they did not report whether the three-way interaction between the three types of repetition was significant, as their account would predict: indeed, such an interaction should emerge if any partial repetition slows selection relative to a complete change. In addition, it was not possible to determine whether repetition of the reported feature or of the motor response drove the interaction with the target-defining feature, because these were confounded in that study.
The latter limitation also applies to most of the studies held to support the idea that episodic retrieval affects motor responses in visual search. The main piece of evidence in these studies is that the benefit of repeating the target-defining feature on successive trials (e.g., Lamy et al., 2010; Lamy et al., 2011; Lamy & Kristjánsson, 2013; Yashar et al., 2013), target location (e.g., Gokce et al., 2014; Hilchey et al., 2018), or target-defining dimension (e.g., Töllner et al., 2008) is typically much larger when the reported feature repeats than when it changes. Yet, only two studies addressed the distinction between repetition of the motor response and repetition of the reported feature.
Töllner et al. (2008) used event-related brain potentials (ERPs) and examined whether the interaction between dimension repetition and repetition of the reported feature (or motor response) modulated the lateralized readiness potential (LRP). They found the interactive pattern only in the latency of the stimulus-locked LRP but not in the latency of the response-locked LRP. As stimulus-locked LRP differences may reflect differences pertaining to any processing stage(s) occurring prior to response planning, these findings constrain but do not pin down the exact stage at which the interaction emerged.
Yashar and Lamy (2011) adopted a different approach and associated two rather than just one reported feature to each of two alternative motor responses (i.e., one button press, if either the left or right corner of the target diamond was chipped, and another button press if either the upper or lower corner of the target diamond was chipped). This design allowed them to test the effects of motor-response and reported-feature repetitions independently. The authors found that only repetition of the motor response interacted with repetition of the target-defining feature, thereby supporting the idea that episodic retrieval affects the selection and/or execution of motor responses during visual search.
Current research on the role of episodic retrieval in inter-trial priming effects during visual search suffers from three main limitations. First, unlike in the action-control literature, only a minority of studies examined interactions between different types of repetitions, whereas examining these interactions is the gold-standard tool to study episodic retrieval in the action-control literature. For example, Kristjánsson (2006) conducted a systematic investigation of the effects of repeating target-defining, reported and irrelevant features on three different feature dimensions but did not report the interactions between these effects. In a later study, Kristjánsson (2009) reported the interactions between relevant and irrelevant features but did not examine whether these were modulated by response repetitions—which is crucial to link attentional selection and motor control processes. More generally, in its current state, the visual-search literature does not provide a detailed answer as to whether each feature of the target object retrieves the response associated with this object independently, or whether the conjoined target features have a unique contribution in retrieving this response.
Second, as we showed in the previous section, very few studies disentangled stimulus-related repetitions (i.e., repetition of the target’s reported feature) from action-related repetitions (i.e., motor-response repetition), a distinction that is fundamental for bridging between episodic-retrieval accounts in the visual-search and action-control literatures.
Finally, very few studies have exploited known properties of episodic retrieval processes to investigate the role of these processes in visual search. One notable exception is a series of experiments conducted by Thomson and Milliken (2011, 2012, 2013). These authors reasoned that if inter-trial priming of the target- and distractor-defining features (aka, PoP, Maljkovic & Nakayama, 1994) reflects episodic-retrieval processes, these effects should be sensitive to contextual effects. Accordingly, they explored whether repeating versus changing the task context or increasing the discriminability of a search trial by endowing it with rare target and distractor colors, with a rare location, or with a rare stimulus configuration would influence the effect. These authors reported mixed findings. On the one hand, task changes reduced inter-trial priming and the effect lasted considerably longer for rare than for frequent-color trials. On the other hand, there was no effect of rare versus frequent locations or configurations. Crucially, however, the authors did not examine interactions involving motor responses. Relatedly, Lamy et al. (2011) showed that response retrieval can rely on more global aspects of the search episode: they found that response repetition sped search when both the target and distractor colors repeated and slowed search when these colors swapped but had no effect on performance when either the target’s or the distractors’ color changed to a new color.
Taken together, there is to date little evidence that contextual information unrelated to the target’s or distractors’ features can trigger episodic retrieval of a similar previous trial, in tasks that involve search—unlike in the action-control literature.
The Role of Display Complexity and Salience in Action Control
While the previous section focused on the role of retrieval in visual search, discussed through the lens of action-control studies, we now focus on the selection stage (that has been neglected in action control research) against the background of visual-search theorizing. BRAC (Frings, et al., 2020) emphasizes that integration and retrieval are two independent processes that can be separately modulated by top-down and bottom-up influences. Different modulators can increase or weaken integration/retrieval strength (each separately), and thereby increase/reduce S-R binding effects.
Hommel (2004) had initially suggested that salience can modulate feature integration and that a more salient stimulus should therefore become more strongly integrated with a response. However, this author later developed a different view on integration and binding processes, according to which everything is unconditionally integrated without any effects of bottom-up factors such as salience (Hommel, 2005). Thus, the currently prevailing assumption is that all stimuli during the prime event are equally integrated with the response into an event file. Despite the strong implications of the role of bottom-up influences for action-control research, the empirical evidence on this issue is surprisingly scarce and inconsistent.
On the one hand, indirect support for the claim that bottom-up influences do not affect binding processes comes from recent studies that analyzed the influence of perceptual modulators of binding other than salience, namely, figure-ground segmentation (Schmalbrock & Frings, 2022) and grouping (Schmalbrock et al., 2022). In a nutshell, these studies showed that grouping and figure-ground segmentation modulate retrieval but do not affect binding/integration.
On the other hand, some recent studies explored salience effects on action control and reported findings that are at odds with the idea of unconditional binding. Complementing previous studies on the prioritization of action-control mechanisms by top-down modulators through feature weights (Memelink & Hommel, 2013), Schmalbrock et al. (2021) was the first study to highlight the role of the bottom-up stimulus salience. They explored how salience affects S-R integration in a variant of Hommel’s (1998) S1R1-S2R2-paradigm. They presented the relevant prime letter alongside seven filler letters. The relevant letter had either the same color as the filler letters (non-salient prime letter) or a different color (salient prime letter). S-R binding effects were found only for salient prime letters. These results were interpreted as evidence that salience modulates the integration stage—since the salience manipulation was only applied to the prime, the results could not reflect modulation of the retrieval stage. Relatedly, Qiu et al. (2022) showed that salient contexts (an accompanying tone with varying loudness or emotional content) boosted the integration of distractors and responses in a forced-four-choice auditory negative-priming task (Mayr & Buchner, 2006).
As is clear from the foregoing review, the role of salience in action control is still poorly understood. Because research on action control has focused on the demands on response selection, in most experimental tasks in this field displays consisted of very few stimuli, thereby effectively not taxing mechanisms responsible for stimulus selection (see Figure 2). In contrast, in visual-search studies, displays typically include many stimuli and the processes involved in stimulus selection therefore play a prominent role. Relying on the observation that their earlier finding of a salience effect on integration (Schmalbrock et al., 2021) was observed with prime displays containing an unusual large number of stimuli (eight), Schmalbrock et al. (2023) surmised that the effect of salience on S-R binding effects in action-control might be contingent on display complexity. To test this hypothesis, they contrasted prime displays with few versus many stimuli. They found that salience indeed impacted upon feature binding only in displays with many stimuli. These findings suggest that the effect of salience on feature integration and binding in previous action-control studies has likely been largely underestimated, because these studies used very few stimuli and therefore posed low demands on the selection stage.
For experiments in the laboratory that focus on specific aspects of response selection, the fact that stimulus selection has been neglected does not pose a problem. However, most theoretical frameworks in the action-control literature (such as TEC, Hommel, 1998, 2004 or BRAC, Frings et al., 2020) are meant to describe actions not only in the lab but also in the real world. If action-control theories claim this scope, they must acknowledge the impact of selection mechanisms and their determinants (such as salience) in perceptually more complex scenarios.
Concluding Remarks
In their daily lives, humans select sensory input, coordinate it with current goals, and then select responses to interact with the environment. In cognitive-science research, these different processes have been largely insulated from each other, and specific research traditions, investigating the details of these stages, have developed separately. These research strands, which were originally motivated by the overarching question of how humans interact with their environment, have produced specific paradigms and/or phenomena and specific explanatory frameworks/theories tailored to these. While this approach has produced many intriguing insights into the mechanisms involved in selecting or responding, it has also led to considerable fragmentation in cognitive-science research, thereby impeding scientific progress (Balietti et al., 2015). There is thus an urgent need to increase efforts in theory-building (Eronen & Bringmann, 2021) and to better re-link psychological research fields that examine the very same concepts from different perspectives. We here suggest such a link between visual search and action control via striking structural similarities in task design. We argue that the commonalities between these lines of research should be emphasized and that findings and theories from one field hold a strong—yet hitherto unharnessed—potential to boost progress in the other.
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Deutsche Forschungsgemeinschaft; FOR2790, Israel Science Foundation; 2449/21, Center for Advanced Studies at Ludwig-Maximilians-University Muchen; Handling distraction group.