
Abstract
Dietary modification in early life can either increase susceptibility or improve resistance to metabolic diseases in adulthood. This study evaluated the potential of lycopene to mitigate metabolic derangements caused by a high-fructose diet in rats mimicking adolescents fed a high-fructose diet. Ninety-six weanling Wistar rats (male and female, aged 21 days) were randomly allocated to six treatment groups: (1) standard rat chow (SRC), plain drinking water (PDW), and plain gelatin cubes (PG); (2) SRC + PDW + 20% fructose solution (FS); (3) SRC + FS + 100 mg/kg fenofibrate (FENO) + PG; and (4–6) SRC + FS with lycopene at 30, 60, and 100 mg/kg/day, respectively, for 12 weeks. Body mass, feed, and fluid intake were measured twice weekly. At termination, rats were fasted overnight, weighed, euthanized, and assessed for gastrointestinal viscera, femora, and tibiae parameters, including mass and length, with bone mass-to-length ratios computed. The rats grew significantly (P < .05) during the trial. Control males had higher mean weekly body mass than medium-dose lycopene-supplemented rats (weeks 3–4) and fenofibrate-treated rats (weeks 10–12), and higher terminal body mass than fructose-fed and fenofibrate groups (P < .05). In both sexes, controls had greater total feed intake (TFI) than all groups (P < .05). Lycopene reduced TFI but increased fluid intake compared with control and fenofibrate-treated groups (P < .05). Fenofibrate increased total calorie intake in females compared with all groups and in males compared with fructose-fed and lycopene-supplemented groups (P < .05). Lycopene-supplemented males had heavier femora and longer tibiae than fenofibrate-treated counterparts (P < .05). Medium- and high-dose lycopene increased stomach mass in males compared with controls, while low-dose lycopene reduced small intestine mass compared with fenofibrate (P < .05). Lycopene may support bone health, enhance gastric function, and prevent fenofibrate-induced bone loss in male Wistar rats.
INTRODUCTION
Dietary changes during early life can significantly impact the risk of developing metabolic disorders or contribute to improved health outcomes in adulthood (Reynolds et al., 2015). Metabolic syndrome affects individuals across all age groups worldwide (Bitew et al., 2021), with an estimated global prevalence of approximately 3.3% among children and adolescents (Al-Hamad and Raman, 2017), ranging from 0.2% to 38.9% (Al-Hamad and Raman, 2017). Among other factors, the increased dietary intake of fructose has been and is associated with the rising incidence of metabolic disorders (Aoun et al., 2022). Following its absorption from the small intestine, fructose is primarily metabolized through fructolysis, glycolysis, gluconeogenesis, and lipogenesis in the liver (Hannou et al., 2018).
The gastrointestinal tract (GIT) is essential for nutrient digestion and absorption (Basile et al., 2023). Excessive fructose intake has been shown to disrupt energy balance, body mass regulation, and growth, as well as induce morphological changes in the GIT system (Guney et al., 2023). Furthermore, it alters the composition of gut microbiota, mediating increased lipopolysaccharide by Gram-negative bacteria (Beisner et al., 2020). This shift can lead to alterations in the morphometry of visceral and accessory organs within the GIT (Zhang et al., 2017). Consequently, these lipopolysaccharide-induced alterations in GIT morphology could increase the risk of developing metabolic disorders (Taskinen et al., 2019). In addition to its effects on the GIT, high dietary fructose negatively impacts bone metabolism by disrupting the absorption, reabsorption, and elimination of essential vitamins and minerals necessary for healthy bone growth (Kuan et al., 2018). In children and adolescents, a balanced diet is essential for proper bone growth and development and helps reduce the risk of osteoporotic fractures later in life (Proia et al., 2021). Osteoporosis, a condition affecting all races, ethnicities, and genders (Askari et al., 2019), is characterized by decreased bone mass, lower bone mineral density, compromised microarchitectural integrity, and reduced bone strength (Barnsley et al., 2021). This leads to increased bone fragility and a higher incidence of fractures, particularly in the spine, hip, and humerus (Morin et al., 2014). Globally, osteoporosis affects over 200 million people (Akkawi and Zmerly, 2018), with vertebral or femoral fractures occurring approximately every 200 sec (De Martinis et al., 2021). Individuals with spinal cord injuries are more prone to femora and tibiae fractures (Zheng et al., 2021). At age over 50 years, about 33% of women and 20% of men will suffer a fragility fracture at some point in their lives (Shen et al., 2022).
Addressing the negative effects of a high-fructose diet on growth, GIT morphology, and bone development and health is crucial. Functional foods rich in secondary plant metabolites offer a promising solution for counteracting these diet-induced changes. These metabolites, which primarily protect plants from herbivores and various pathogens (Al-Khayri et al., 2023), also offer health benefits that can help mitigate diet-induced metabolic disturbances (Divekar et al., 2022). Lycopene, abundantly found in tomatoes and watermelons, is a lipophilic carotenoid with potent antioxidant and anti-inflammatory properties (Rasmus and Kozłowska, 2023). It has been shown to protect the gastrointestinal tract mucosa and enhance nutrient absorption (Rajput et al., 2021). Furthermore, it promotes growth (Wu et al., 2024) and supports bone health (Costa-Rodrigues et al., 2018).Therefore, we aimed to evaluate the effects of supplemental lycopene on feed per calorie and fluid intake, growth performance, GIT viscera macro-morphometry, and the indices of femora and tibiae in growing Wistar rats fed a high-fructose diet.
MATERIALS AND METHODS
Ethical approval and study site
The study was conducted at the Wits Research Animal Facility (WRAF), University of the Witwatersrand, Johannesburg, South Africa. Ethical clearance was obtained from the University’s Animal Ethics Screening Committee (Certificate: AESC 2022/03/02/C). The research strictly adhered to the Animal Research: Reporting of In Vivo Experiments (ARRIVE) guidelines, ensuring compliance with ethical standards throughout all experimental procedures.
Chemicals and reagents
All chemicals and reagents utilized were of analytical grade. Fenofibrate was sourced from Sigma-Aldrich (St. Louis, MO), fructose from Nature’s Choice (Randvaal, South Africa), and lycopene was procured from Changsha Staherb Natural Ingredient Co., Ltd. (Hunan, China).
Animals, housing, and general care
Ninety-six 21-day-old Wistar rat pups (48 male; 48 female) were used for the study. These weanling Wistar rats were sourced from litters of first-time dams, each litter containing 8–12 pups, and were individually housed in standard Perspex cages with wood shavings as bedding at the WRAF, University of the Witwatersrand. Bedding was replaced twice weekly. The rats were exposed to a 12-h light and darkness cycle, with light on from 7 a.m. to 7 p.m. Room temperature was maintained at 24°C ± 2°C. A standard commercial rat feed supplied by Pet Food Industry of Southern Africa, and either plain drinking water (PDW) or 20% (w/v) fructose solution (FS), depending on treatment group, was provided ad libitum. The nutrient composition of the commercial rat feed was as follows: 40 g/kg fiber, 70 g/kg ash, 100 g/kg moisture, 220 g/kg protein, 50 g/kg oils and fats, 12 g/kg linoleic acid, 7.5 g/kg phosphorus, 12 g/kg calcium, 16,000 IU/kg vitamin A, 2000 IU/kg vitamin D, and 100 g/kg vitamin E. The fructose used contained 1680 kJ energy, 0 g protein, 0 g fiber, 0 g fat, 99.8 g/100 g carbohydrates, and 0.2 mg/100 g sodium. The 20% (w/v) FS was prepared by dissolving 20 g of fructose in 100 mL of drinking water.
Experimental design
On postnatal day (PND) 23 following a 2-day habituation period, the rats were randomly allocated to one of the treatment groups:
Control: standard rat chow (SRC) + PDW + plain gelatin cube (PG), Negative control: SRC + 20% w/v FS as drinking fluid + PG, Positive control: SRC + FS + 100 mg/kg body mass per day of fenofibrate in gelatin cube (FENO), Low dose: SRC + FS + lycopene 30 mg/kg body mass per day in gelatin cube (LDLY), Medium dose: SRC + FS + lycopene 60 mg/kg body mass per day in gelatin cube (MDLY), High dose: SRC + FS + lycopene 100 mg/kg body mass per day in gelatin cube (HDLY).
Each group comprised 16 rats: 8 males and 8 females, and treatments were administered for 12 weeks. The fenofibrate dose (Kopf et al., 2014) and lycopene doses were as previously used by Gustin et al. (2004) and Zhang et al. (2020), respectively. The gelatin cube vehicles for fenofibrate and lycopene administration were prepared following the procedure described by Kamerman et al. (2004), with a slight adjustment. Instead of using 16 g of brown sugar, 8 g Selati sugar (RCL Foods and Sugar Milling Ltd., South Africa) were utilized as it was found that a higher sugar concentration led to decreased intervention intake.
The sample size was determined using the formula:
N = DF/(K − 1) + 1,
where N is the number of subjects per group, DF is the degrees of freedom, and K is the number of groups (Arifin and Zahiruddin, 2017).
Substituting the values:
N = 20/(6 − 1) + 1 = 20/5 = 4 + 1 = 5 rats per group.
Thus, the minimum required sample size was five rats per group. However, to increase the reliability of the results and allow for possible losses, eight rats per group were used. With six treatment groups for each sex, this gave a total of 96 rats (48 males and 48 females) used in the study. Figure 1 shows the schematic diagram of the study design.
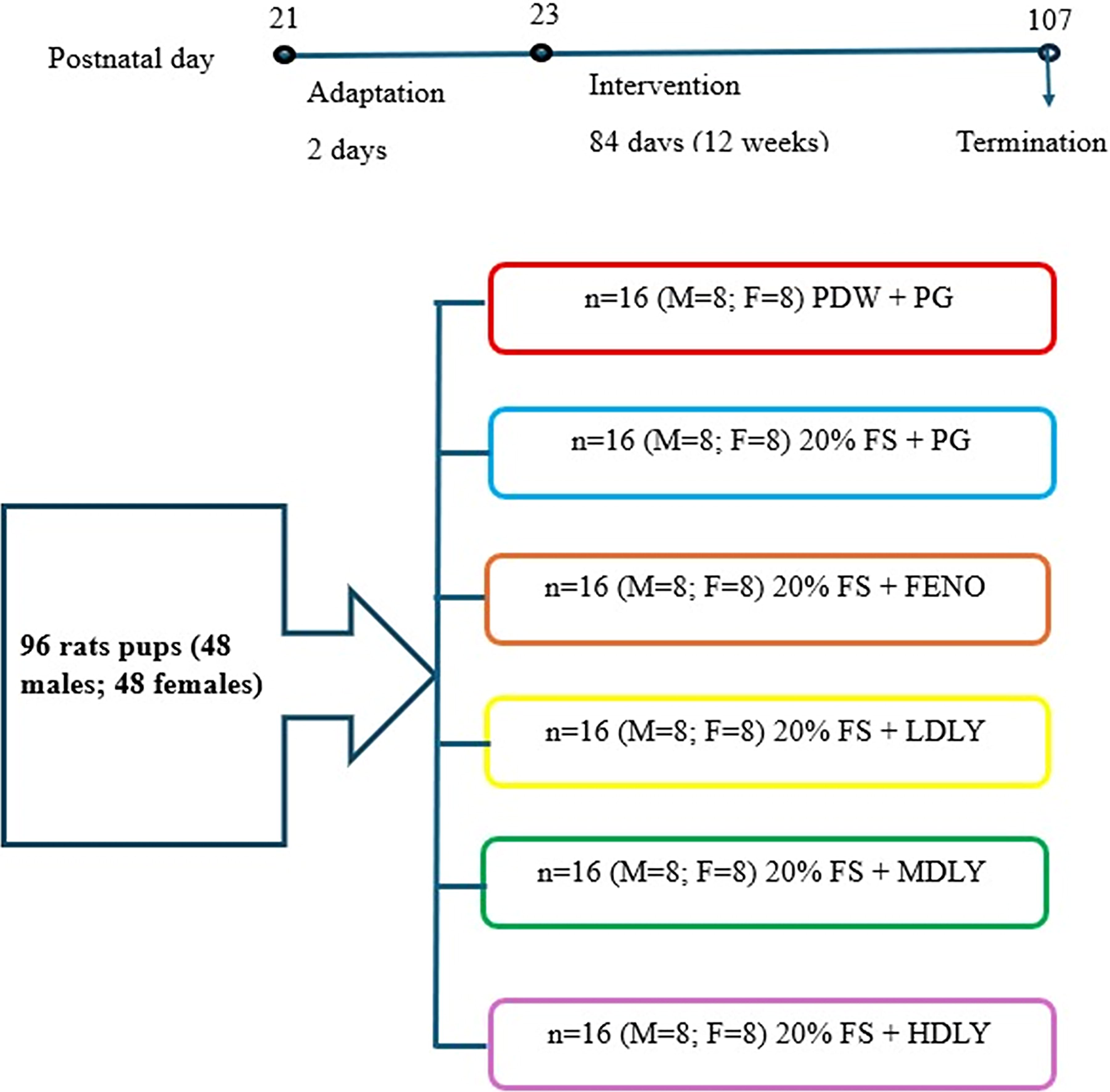
Schematic diagram of study design. FS, fructose solution (20%); FENO, fenofibrate 100 mg/kg body mass (BM) per day; HDLY, high-dose lycopene 100 mg/kg BM per day; LDLY, low-dose lycopene 30 mg/kg BM per day; MDLY, medium-dose lycopene 60 mg/kg BM per day; PDW, plain drinking water; PG, plain gelatin cube.
Measurements and computations
Following measurement of the induction body mass using an electronic balance (Snowrex Electronic Scale from Clover Scale [Pty] Ltd., Johannesburg), the rats were thereafter weighed twice a week. The twice-weekly weighing sought to track the rats’ growth performance and overall health and to maintain a consistent dose of fenofibrate and lycopene throughout the trial. Weekly feed and fluid intake were determined throughout the trial. At the end of each week, each rat’s weekly total feed and fluid intake were computed and expressed, as a percentage of body mass, in g/100 g and mL/100 g, respectively. Total weekly calorie intake was calculated by multiplying the weekly consumption of both feed and fructose by their corresponding reference calorie values and the resulting values were summed. The total intake of feed, fluid, and calories for each experimental group was determined by summing up the respective weekly consumption over the 12-week period.
Terminal procedures and sample collection
On PND 106, the rats were subjected to an 11-h overnight fast but had ad libitum access to PDW to prevent dehydration. The morning after (PND 107), each rat’s body mass was recorded using an electronic balance (Snowrex Electronic Scale from Clover Scale [Pty] Ltd., Johannesburg). Following this, the rats were euthanized using an intraperitoneal injection of sodium pentobarbitone at 200 mg/kg body mass (Euthapent; Kyron Laboratories, Johannesburg, South Africa). A midline incision was made in the abdomen of each rat and the stomach, cecum, small intestine, and large intestine were meticulously extracted and weighed using an electronic balance (Presica 310M, Instruments, Johannesburg, South Africa). The length of the small and large intestines was measured with a ruler fixed on the dissection board. The femoral attachment from the left hind leg to the pelvis was excised from each rat’s carcass, sealed, and preserved in Ziplock bags at −20°C until the long bones were dissected.
Determination of femora and tibiae length, mass, and mass-to-length ratio
The frozen-stored femora and tibiae were thawed at room temperature for about 45 min and thereafter, using a pair of scissors and scalpels, were defleshed and then detached from each other. The femora and tibiae were then dried in an oven (LABOTEC [Pty] Ltd., South Africa) at 50°C for 5 days to constant masses. Each dried bone was then weighed on an electronic balance (Presica 310M, Instruments, Johannesburg, South Africa) and each bone’s length was measured using a digital Vernier caliper (Major Tech [Pty] Ltd., KTV 150 digital caliper, Elandsfontein, South Africa). Tibia length was assessed from the tibia head to the medial malleolus, and femur length was measured from the distal femoral articular surface to the greater trochanter.
The bone mass-to-length ratio was determined using the equation by (Seedor et al., 1991) as: bone mass-to-length ratio = mass of bone (mg)/length of bone (mm).
Statistical analysis
The data were analyzed using GraphPad Prism version 9.0 (GraphPad Software Inc., San Diego, CA) and presented as mean ± standard deviation. Parametric data from multiple groups were assessed using a one-way analysis of variance (ANOVA), and weekly body mass, feed, fluid, and calorie intake were analyzed using a repeated-measures ANOVA. Tukey’s post hoc test was used for the mean comparison. Statistical significance was defined as P < .05.
RESULTS
Growth performance
Figures 2 and 3 show the mean weekly, induction, and terminal body masses of the female and male rats, respectively. There were no significant differences (P > .05) in the mean induction, weekly body mass, and terminal body masses across treatment groups of the female rats (Fig. 2), and there was no difference (P > .05) in the males’ induction body mass (Fig. 3). Both female and male rats grew significantly during the course of the trial (P < .00001, Figs. 2 and 3). Control males’ mean weekly body mass was higher (P < .05) compared with that of counterparts administered medium lycopene dose at weeks 3 and 4 and fenofibrate at weeks 10, 11, and 12 (Fig. 3), and they had heavier (P < .05) terminal body masses compared with fructose-fed counterparts but with fenofibrate as an intervention. However, the control males’ terminal body mass was similar (P > .05) to that of the fructose-fed and lycopene-administered counterparts (Fig. 3).
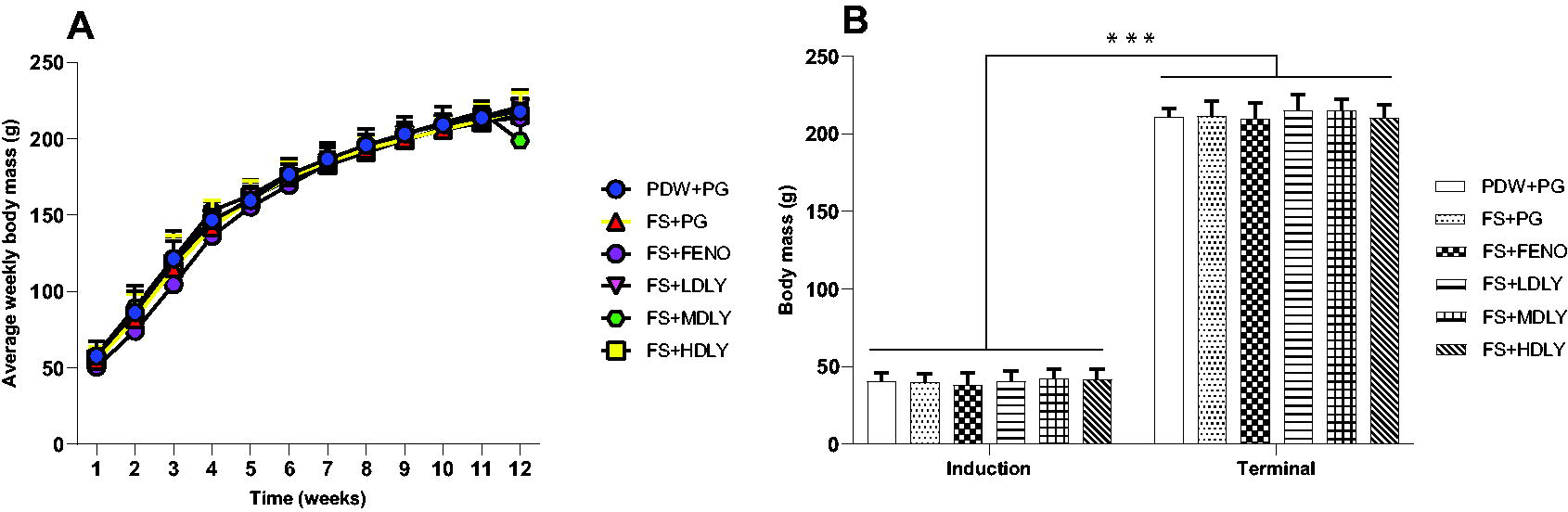
Weekly body masses
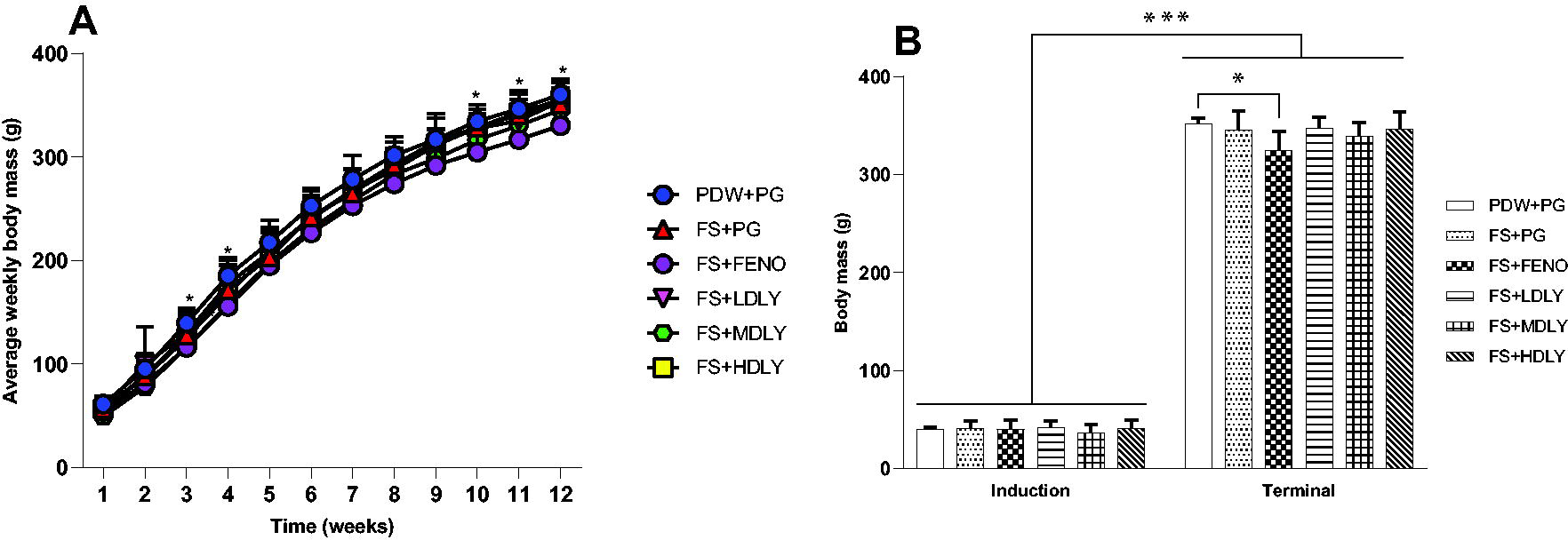
Weekly body masses
Feed, fluid, and calorie intake
Feed, fluid, and calorie intake of the female and male rats is shown in Figures 4 and 5, respectively. In females, week 1 mean weekly feed intake (MWFI) was similar (P > .05) across treatments, but it (MWFI) differed (P < .05) from weeks 2 to 12. In weeks 2–9 and week 11, fenofibrate-administered females had lower MWFI compared with controls. The fructose-fed and lycopene-administered counterparts had lower (P < .05) MWFI compared with controls from weeks 3 to 12, but fenofibrate-administered females had significantly higher (P < .05; Fig. 4A) MWFI from weeks 5 to 11 compared with the fructose-fed and lycopene-administered counterparts. The total feed intake (TFI) of fructose-fed female rats and their counterparts fed the high-fructose diet supplemented with the low and medium lycopene doses was similar (P > .05), and the TFI of females administered the low and high lycopene doses was also similar (P > .05; Fig. 4B). The females’ mean weekly fluid intake (MWFLI) from weeks 1 to 5 and in week 11 was similar (P > .05) across treatment groups, but at weeks 6 and 10, the MWFLI of the fenofibrate-administered females was lower (P < .05) compared with controls and low-dose lycopene-administered counterparts. However, the MWFLI of the low-dose lycopene-administered females was higher (P > .05; Fig. 4C) compared with controls at week 12. The total fluid intake (TFLI) of control females and their fructose-fed and medium-dose lycopene-supplemented counterparts was similar (P > .05), as was that of the fructose-fed and the medium- and high-dose lycopene-administered groups (Fig. 4D). The female rats’ mean weekly calorie intake (MWCI) was similar (P > .05) from weeks 2 to 6, weeks 8 and 11, and week 12 across treatment groups. Fructose-fed rats and their fructose-fed but medium-dose lycopene-supplemented counterparts had higher MWCI compared with controls in week 1 (P < .05). Fenofibrate-administered females had higher (P < .05) MWCI in week 7 compared with the controls, fructose-fed, and medium dose of lycopene-administered counterparts, and in weeks 9 and 10, they also had higher (P < .05, Fig. 4E) MWCI compared with the fructose-fed and medium- and high-dose lycopene-supplemented counterparts. Total calorie intake (TCI) of control, fructose-fed, and high-dose lycopene-supplemented females was similar, as also was the TCI of the fructose-fed females and their lycopene (low, medium, and high dose) counterparts (P > .05; Fig. 4F). Lycopene-supplemented females had similar TCI.
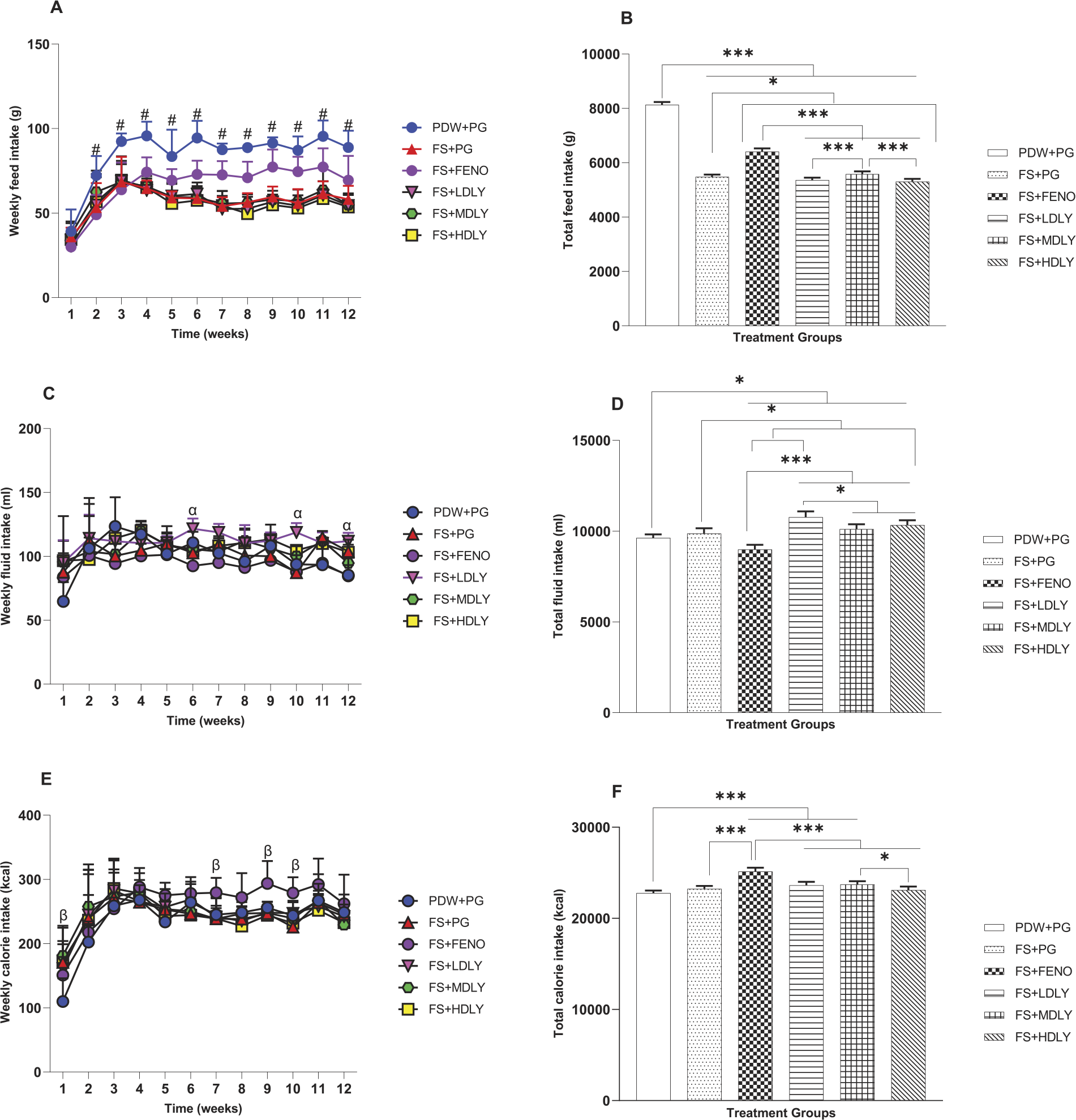
Effect of lycopene on weekly feed
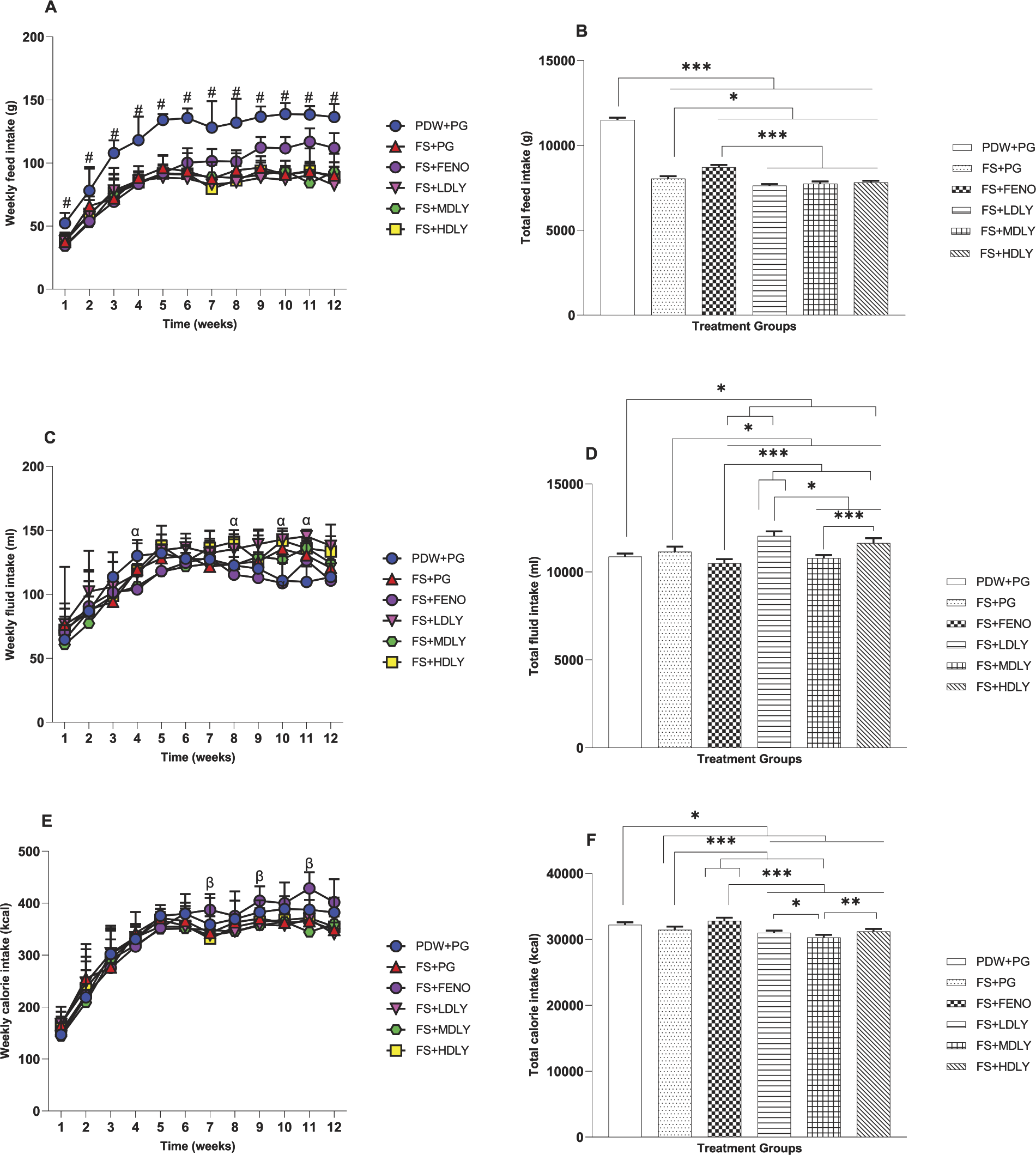
Effect of lycopene on weekly feed
Compared with control counterparts, the fructose-fed, fenofibrate, and lycopene-administered male rats had lower MWFI from weeks 1 to 12. In week 2, the MWFI of the fructose-fed but medium-dose lycopene-supplemented males was lower compared with that of controls (P < .05), and the MWFI of males administered fenofibrate was higher (P < .05) compared with their low- and high-dose lycopene-supplemented counterparts from weeks 7 to 12. However, week 10 MWFI of the male fenofibrate group was higher (P < .05) compared with that of their lycopene-supplemented counterparts. Similarly, the MWFI of the fenofibrate-administered males was higher compared with the fructose-fed group in weeks 9–12 (P < .05; Fig. 5A). The TFI of the lycopene-supplemented males was similar, as also was that of the fructose-fed group and their medium- and high-dose lycopene-supplemented counterparts (P > .05; Fig. 5B). Male rats’ MWFLI was similar (P > .05) from weeks 1 to 3, 5 to 7, week 9, and week 12 across treatments. Week 4 MWFLI of the fructose-fed but medium-dose lycopene-supplemented males was lower compared with controls, but the MWFLI of fructose-fed and lycopene (low, medium, and high dose) supplemented males was higher than their control counterparts in weeks 10–11. Fenofibrate-administered males had lower MWFLI (P < .05) compared with high- and low-dose lycopene-supplemented counterparts in weeks 8 and 10, respectively (Fig. 5C). The TFLI of controls, fructose-fed, and low-dose lycopene-supplemented males was similar, as was that of fructose-fed, fenofibrate-administered, and medium-dose lycopene-supplemented males (P > .05; Fig. 5D). Male rats’ MWCI across treatment groups was similar (P > .05) from weeks 1 to 6 and in weeks 8, 10, and 12, but the MWCI of the fenofibrate-administered males was higher (P < .05; Fig. 5E) compared with that of the low-, medium-, and high-dose lycopene-supplemented counterparts in weeks 7, 9, and 11, respectively. The TCI of fructose-fed males and their low- and high-dose lycopene-supplemented counterparts was similar, as also was that of the fenofibrate group and the controls (P > .05; Fig. 5F).
Long bone indices
The effect of lycopene on femora and tibiae indices of female and male rats fed a high-fructose diet was assessed. In females, there were no differences in the bones’ lengths, masses, and mass-to-length ratio across treatment groups (P > .05; Table 1). The femora masses of males administered fenofibrate were lower than those from low-dose lycopene-supplemented counterparts, but femora masses of the controls, fructose-fed, and lycopene (low, medium, and high dose) supplemented males were similar (P > .05; Table 2). Tibiae of males administered fenofibrate were shorter (P < .05) compared with those from controls and the lycopene-supplemented counterparts, but mean tibiae lengths of the controls, fructose-fed, and lycopene (low, medium, and high dose) supplemented males were similar (P > .05).
Effect of Lycopene on the Lengths, Masses, and Mass-to-Length Ratio of Tibiae and Femora in Growing Female Wistar Rats Fed a High-Fructose Diet
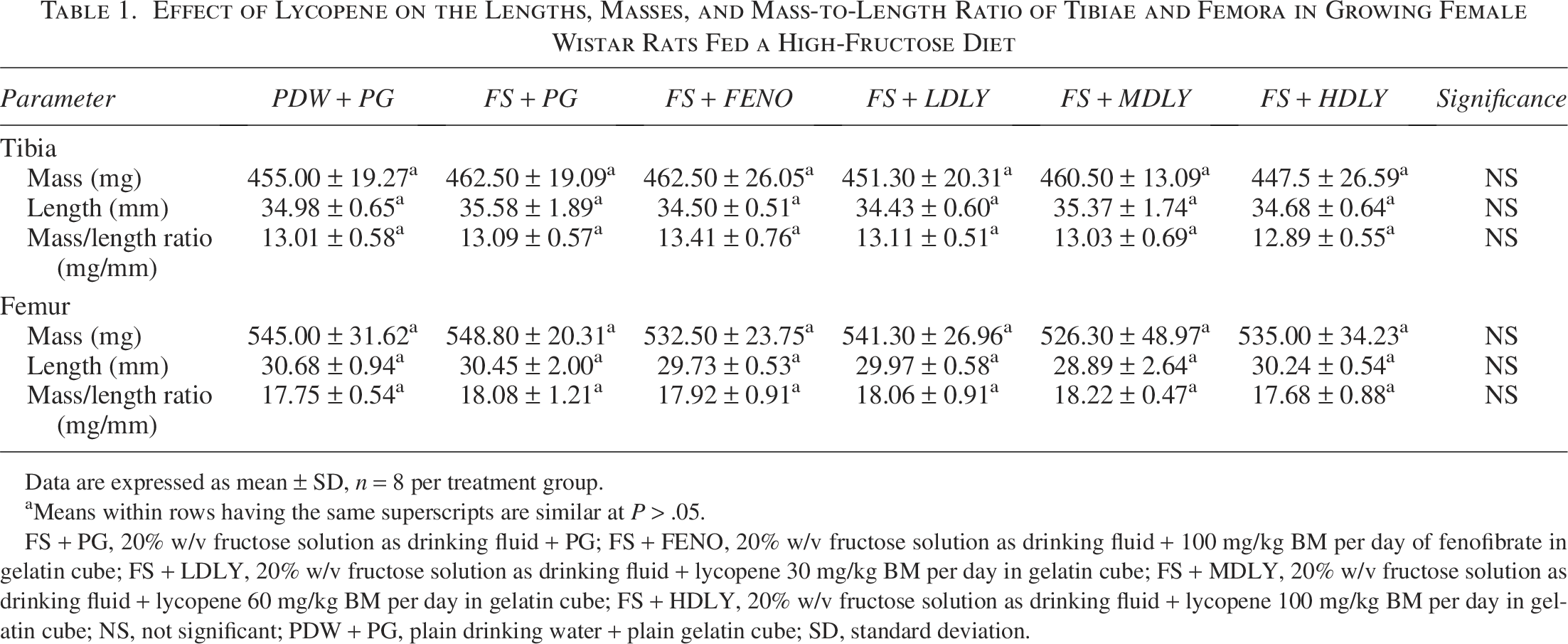
Data are expressed as mean ± SD, n = 8 per treatment group.
aMeans within rows having the same superscripts are similar at P > .05.
FS + PG, 20% w/v fructose solution as drinking fluid + PG; FS + FENO, 20% w/v fructose solution as drinking fluid + 100 mg/kg BM per day of fenofibrate in gelatin cube; FS + LDLY, 20% w/v fructose solution as drinking fluid + lycopene 30 mg/kg BM per day in gelatin cube; FS + MDLY, 20% w/v fructose solution as drinking fluid + lycopene 60 mg/kg BM per day in gelatin cube; FS + HDLY, 20% w/v fructose solution as drinking fluid + lycopene 100 mg/kg BM per day in gelatin cube; NS, not significant; PDW + PG, plain drinking water + plain gelatin cube; SD, standard deviation.
Effect of Lycopene on the Lengths, Masses, and Mass-to-Length Ratio of Tibiae and Femora in Growing Male Wistar Rats Fed a High-Fructose Diet
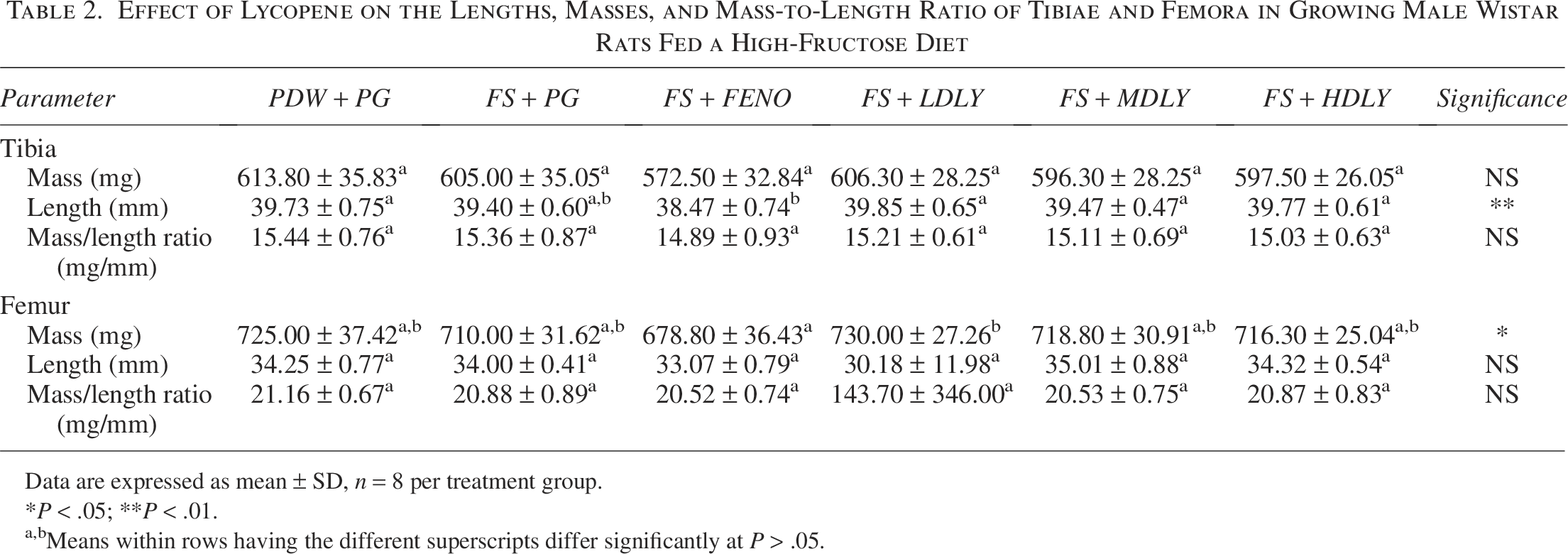
Data are expressed as mean ± SD, n = 8 per treatment group.
*P < .05; **P < .001.
a,bMeans within rows with different superscripts differ significantly at P < .05.
Gastrointestinal tract viscera macro-morphometry
The effect of lycopene on the absolute and relative masses and length (where applicable) of the GIT viscera of the female and male rats fed a high-fructose diet is shown in Tables 3 and 4, respectively. Treatments had no effect (P > .05) on the female rats’ GIT viscera masses and lengths (Table 3). However, the mean absolute and relative stomach mass of control males was lower (P < .05, Table 4) compared with those from the fenofibrate-administered and medium- and high-dose lycopene-supplemented counterparts, but the mean stomach masses of the controls, fructose-fed, and low-dose lycopene-supplemented males were similar (P > .05).
Effect of Lycopene on Masses and Lengths of Gastrointestinal Tract Viscera of Growing Female Rats Fed a High-Fructose Diet
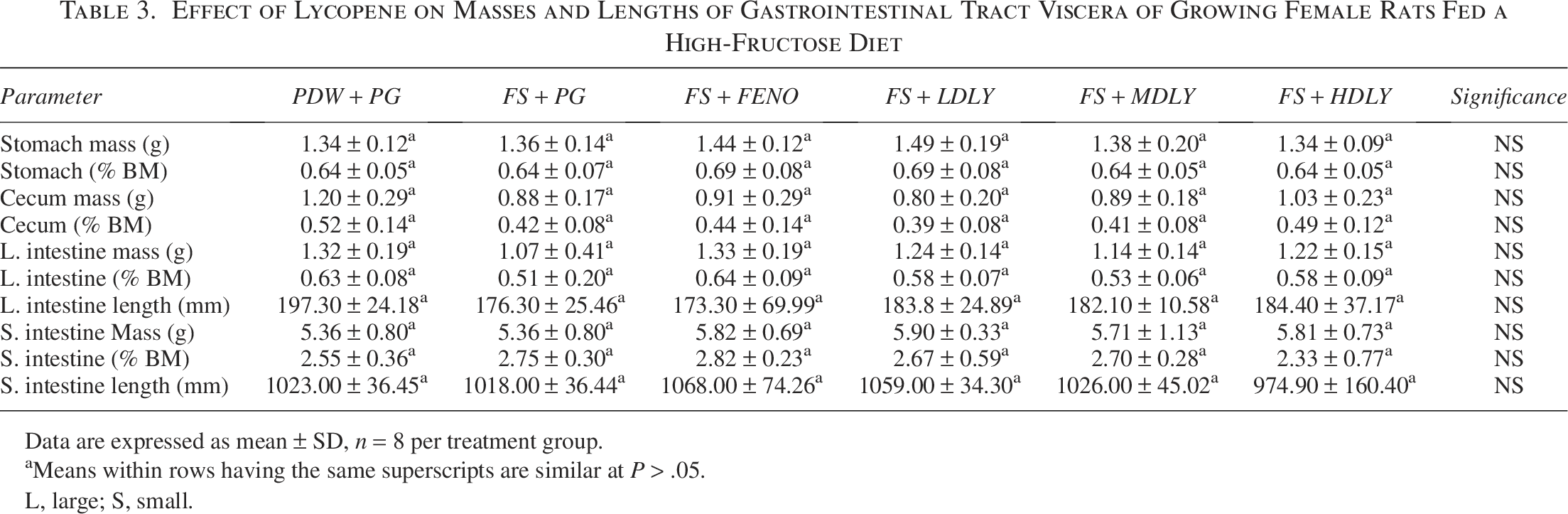
Data are expressed as mean ± SD, n = 8 per treatment group.
aMeans within rows having the same superscripts are similar at P > .05.
L, large; S, small.
Effect of Lycopene on Masses and Lengths of Gastrointestinal Tract Viscera of Growing Male Rats Fed a High-Fructose Diet
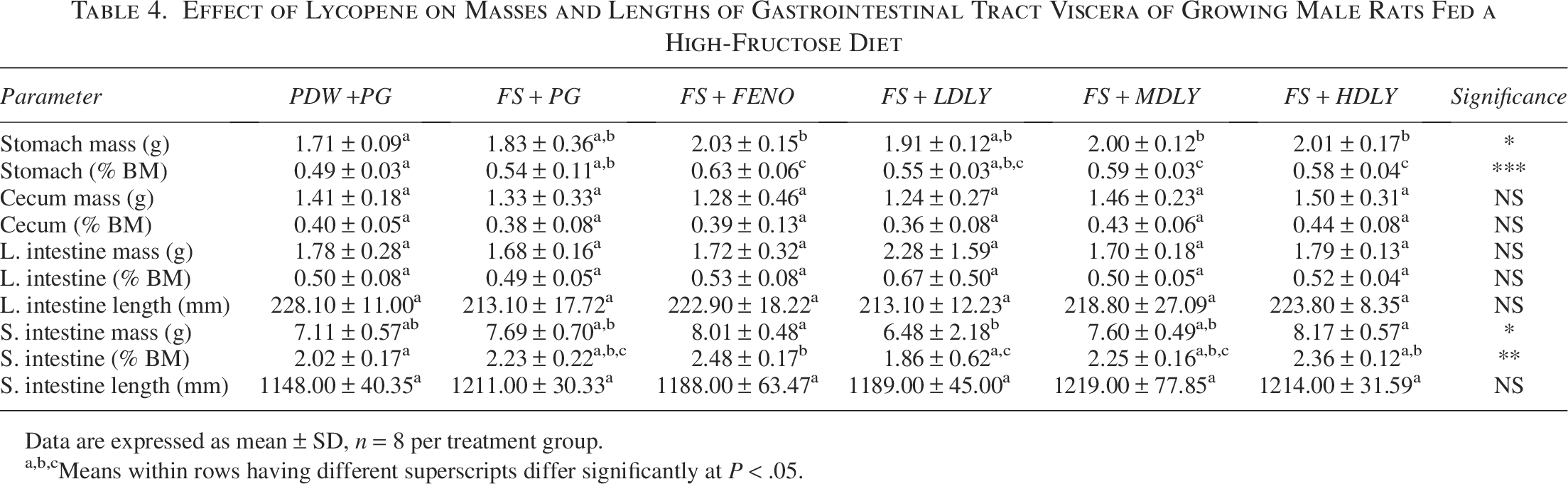
Data are expressed as mean ± SD, n = 8 per treatment group.
Means within rows having different superscripts differ significantly at P < .05.
DISCUSSION
The study evaluated the effect of orally administered lycopene, a phytochemical possessing health-beneficial biological activities, on growth performance, feed, fluid, and calorie intake, and GIT viscera of growing Wistar rats fed a high-fructose diet. Female rats’ mean induction, weekly body, and terminal body masses were similar. The males’ induction body masses were also similar across treatments, but the mean weekly body masses of fenofibrate-treated, as well as the medium-dose lycopene-supplemented males, were significantly lower in weeks 10–12 and weeks 3 and 4, respectively, compared with controls. These findings demonstrate sexually dimorphic effects of fenofibrate and lycopene on the growth of Wistar rats. Importantly, these findings suggest that the medium dose of lycopene (60 mg/kg body mass) was as effective in managing body mass gain in growing male Wistar rats as fenofibrate, a conventional pharmacological agent. Similar to the current study findings, Wang et al. (2019) observed that supplemental lycopene mediated a reduction in body mass gain in high-fat diet-fed male mice. Despite similarities in the mean induction body masses of female and male rats, both rat sexes grew significantly during the trial period, demonstrating that the dietary fructose insult, fenofibrate, and/or lycopene prevented body mass gain. The observed decrease in the mean terminal body mass of males treated with fenofibrate compared with control counterparts mirrors the report by (Lee et al., 2023), who reported a fenofibrate-mediated decreased body mass and adipose tissue mass and size in ovariectomized (estrogen-deficient) mice fed a high-fat diet. Fenofibrate regulates body mass by activating nuclear peroxisome proliferator–activated receptor α (PPARα), which controls the key genes involved in lipid and lipoprotein metabolism by enhancing triglyceride lipolysis, β-oxidation of fatty acids, and reducing triglyceride synthesis and secretion (Shin et al., 2021). The similarity in the terminal body masses of the control female and male rats to that of fructose-fed counterparts, as well as the fructose-fed but lycopene (low, medium, and high dose) counterparts, suggests that dietary fructose did not mediate increased body mass gain, and neither dietary fructose nor supplemental lycopene compromised the rats’ growth during the 12-week intervention. Current study findings align with Ramos et al. (2017), who observed that dietary supplementation with 20% FS as drinking fluid did not alter Wistar rats’ body mass but augmented their visceral adiposity. The elevated adipogenic potential and relatively low energy content of fructose are thought to be accountable for the observed increased visceral fat without a concurrent increase in body mass (Ramos et al., 2017). It is essential to emphasize that while body mass is a crucial marker of an animal’s health, its accuracy as a gauge of growth may be compromised by viscera organ sizes, gut fill, and hydration status, thus decreasing its reliability as an objective measure of growth performance (Borga et al., 2018; Müller et al., 2018; Ekingen et al., 2022).
This study noted differences in feed, fluid, and calorie intake in the rats across treatment groups. In summary, control diet-reared rats consumed more feed, drank less plain water, and had lower TCI. Conversely, rats provided with 20% FS as the drinking fluid drank more fluid, consumed less feed but had higher TCI. Notably, orally administered fenofibrate decreased fluid intake. In both sexes, control diet-fed rats had higher MWFI and TFI compared with the fructose-fed, fenofibrate-treated, and lycopene-supplemented counterparts. Long-term dietary fructose intake has been reported to decrease feed and energy intake (Smajis et al., 2020), impacting the secretion and function of leptin and insulin, hormones that regulate appetite (Capucho and Conde, 2022); a disruption likely to mediate irregular hunger signals and increase the risk of obesity and type 2 diabetes (Cui et al., 2022). Fructose-fed rats but treated with fenofibrate had higher MWFI and TFI compared with plain fructose-fed and lycopene-supplemented counterparts. This suggests that in adolescent fructose-fed rats, orally administered fenofibrate (100 mg/kg body mass daily) mediates increased feed intake, which contradicts the observation that it (fenofibrate) decreases the intake of a SRC by mediating increased ketogenesis from mobilized body fat (Park et al., 2012). Based on our observation wherein fenofibrate decreased the body mass of fructose-fed rats compared with controls, we hypothesize that the increase in feed intake might have been in a bid to mitigate the fenofibrate-induced decrease in body mass. We contend that the dichotomy in our findings could have been due to altered metabolic pathways or hormonal responses leading to increased feed intake. Furthermore, there are possibilities that fenofibrate might have independent effects on appetite or metabolism that contribute(s) to increased feed intake when combined with fructose. These effects could be mediated through various pathways, possibly changes in gut hormone secretion or alterations in central nervous system signaling related to appetite control. The fructose-fed male rats had significantly higher TFI compared with their lycopene (low, medium, and high dose) supplemented counterparts, but in females this difference in feed intake was particularly evident when compared with the high-dose lycopene-supplemented group. Additionally, the TFI of medium-dose lycopene-supplemented females was higher compared with that of their low- and high-dose lycopene-supplemented counterparts. Wang et al. (2019), reported that dietary fortification with lycopene inhibited, in mice, the high-fat diet-mediated increase in body mass, and similar findings were reported by Zhu et al. (2020). Our findings show that supplemental lycopene in fructose-fed rats did not negatively affect body mass, as also was with the controls and the fructose-fed groups. However, we did notice a decrease in feed consumption; thus, we hypothesize that this reduction in feed intake could be linked to the physiobiochemical effects of lycopene on metabolic processes. It is plausible that higher metabolic rates induced by lycopene (Wang, 2012) could lead to decreased feelings of hunger and, subsequently, lower feed consumption. Another explanation could be lycopene’s potential to influence appetite regulation by enhancing leptin sensitivity or reducing ghrelin secretion. Furthermore, lycopene might alter feed taste perception and hence its palatability, affecting rats’ feed preferences and resulting in reduced consumption.
Our findings show similarities in females’ MWFI in 58.3% (weeks 1–6 and 11) of the 12-week intervention period. Importantly, the TFLI of control females, fructose-fed, and medium-dose lycopene-supplemented counterparts was similar. We report no differences in the females’ MWCI across treatment groups in 66.7% (weeks 2–6, 8, 11, and 12) of the 12-week intervention period, but control females (week 1) had lower MWCI compared with the fructose-fed and low-dose lycopene-supplemented groups. In weeks 9 and 10, the fenofibrate-treated group had higher MWCI compared with the fructose-fed and medium- and high-dose lycopene-supplemented groups. Furthermore, we observed similarities in the TCI of control, fructose-fed, and high-dose lycopene-supplemented females, as well as between the fructose-fed females and their lycopene-supplemented counterparts, with no difference in TCI among the lycopene-supplemented females. Our findings show similarities in males’ MWFLI across treatments in 75% (weeks 1–3, 3–7 and weeks 9 and 12) of the 12-week intervention period. The TFLI of control males, fructose-fed, and low-dose lycopene-supplemented males was similar, as was that of fructose-fed, fenofibrate-administered, and medium-dose lycopene-supplemented males. The MWCI of males was similar across treatment groups in 75% (weeks 1–6, 8, 10, and 12) of the 12-week intervention period, but in weeks 7, 9, and 11, fenofibrate-treated males’ MWCI was higher compared with that of lycopene-supplemented counterparts.
Fenofibrate has been shown to alter perception and reduce preference for sweet tastes in adult humans (Kochem and Breslin, 2016; Davis, 2020). This could explain why fenofibrate-treated rats may find the FS less palatable, leading to reduced fluid consumption. Despite this reduction in fluid intake, both female and male rats administered fenofibrate had higher TCI, which could be attributed to fenofibrate’s effects on metabolism, as mentioned earlier. Increased fatty acid oxidation and energy expenditure may necessitate higher calorie intake to meet the body’s energy demands. Interestingly, while supplemental lycopene generally increased TFLI, the low-dose lycopene-supplemented rats consumed more fluid compared with their medium- and high-dose supplemented counterparts. This suggests that different doses of lycopene may affect metabolic processes, such as digestion or nutrient absorption, which in turn might influence the rats’ fluid intake patterns. We speculate that, due to its antioxidant activity, the supplemental lycopene interacts with fructose to modulate thirst and/or fluid intake regulatory mechanisms. This hypothesized interaction could enhance the perceived need for fluid intake, leading to increased consumption in lycopene-supplemented rats compared with their control, fructose-fed, and fenofibrate-treated counterparts. Moreover, it could be posited that, in rats, supplemental lycopene may regulate metabolic processes related to fluid intake regulation and/or thirst mechanisms by altering hormone-mediated signaling pathways that regulate fluid balance, leading to an increase in total fluid consumption. Findings from the current study also showed that in male rats, high-dose supplemental lycopene increased the TFLI and TCI compared with medium-dose supplemented counterparts. Conversely, in female rats, the medium-dose supplemental lycopene resulted in higher TCI compared with high-dose lycopene-supplemented counterparts. Additionally, supplemental lycopene decreased males’ TCI compared with control counterparts, yet in females low- and high-dose supplemental lycopene decreased TCI compared with control counterparts. These findings show dose-dependent lycopene-mediated decreased energy intake in growing Wistar rats, suggesting that supplemental lycopene could be used to manage body mass. In both sexes, fructose-fed rats had both higher TFLI and TCI compared with fenofibrate-treated counterparts, which suggests that fenofibrate suppressed fluid and energy intake. The similarities in control and fructose-fed females’ TFLI and TCI suggest dietary fructose does not alter fluid and energy intake of growing female Wistar rats. Additionally, the similarity in the TFLI of control, fructose-fed, and medium-dose lycopene-supplemented males is interesting as it suggests that neither dietary fructose nor medium-dose supplemental lycopene impacts growing male Wistar rats’ fluid intake. Although the current study made no comparisons between female and male rats, generally the male rats consumed more feed and fluid and hence had greater calorie intake than female counterparts. This dimorphic response in feeding has been reported, with males typically having higher intake than females (Maric et al., 2022). Males and females have varying metabolic pathways that lead to sex-specific diet-induced modifications (Hyer et al., 2019). Generally, male rodents are larger and heavier than females and thus require more energy to maintain their body mass (Maric et al., 2022). This larger size may contribute to their increased feed and fluid intake. Hormonal fluctuations, for example, estrous cycle hormonal changes, have been shown to decrease the appetite and feed intake of female rats (Asarian and Geary, 2013; Alonso-Caraballo and Ferrario, 2019). In males, testosterone has been linked to the regulation of appetite that causes increased feed intake (Alrabadi et al., 2020).
Femora and tibiae longitudinal growth provides a more precise measure of animal growth performance than body mass (Cho et al., 2020), since these antigravity bones exhibit a dose-dependent response to growth hormone (Ranke, 2021), a feature that makes them a more reliable indicator of growth performance. Lycopene has been shown to prevent femora bone loss and increase the number of osteocytes and osteoblasts in ovariectomized rats (Semeghini et al., 2022). In the current study, no significant differences were recorded in femora and tibiae lengths, masses, and mass-to-length ratios across treatment groups in female rats. The findings align with Muhammad et al. (2020), who observed neither dietary fructose nor fenofibrate effects in growing female Sprague Dawley rats fed a high-fructose diet for 10 weeks. Based on the current study findings, it can be inferred that dietary fructose, supplemental lycopene, and treatment with fenofibrate did not negatively affect femora and tibial indices of growing female rats. However, in males supplemental lycopene resulted in longer tibiae compared with fenofibrate-treated counterparts, and the low-dose supplemental lycopene increased their femora masses compared with fenofibrate-treated counterparts. Taken together, current study findings demonstrate sexually dimorphic responses: while females’ bones may not be compromised by dietary fructose and/or fenofibrate, males’ tibiae growth may benefit from supplemental lycopene, and their femora can be potentially protected against fenofibrate-induced loss of mass. The observed enhancements in male rats’ tibiae lengths and femora masses could be attributed to lycopene’s antioxidant properties, which protect bone cells from oxidative damage, as well as enhanced osteoblast activity, thereby promoting bone growth and density, thus contributing to overall bone health. The sex-specific femora mass and tibiae length responses to supplemental lycopene observed in this study could be due to differences in plasma testosterone and estrogen concentrations in females and males, since estrogen and testosterone have different effects on bone growth and metabolism between the sexes (Emmanuelle et al., 2021). During the adolescent-to-adulthood transition, testosterone has been shown to promote linear growth (Breehl and Caban, 2023), increase bone mineral density, and enhance bone microarchitecture (Shigehara et al., 2021). Interestingly, supplemental lycopene has been reported to increase testosterone levels in varicocele-induced Sprague Dawley rats (Antonuccio et al., 2020). These combined effects may explain the observed enhancements in femora mass and tibia length in male rats in this study.
Plant-derived chemical compounds have been demonstrated to affect viscera organ masses (Mir et al., 2019). It is from this observation that underscores the importance of assessing the effect of phytochemicals used as dietary supplements on viscera organs’ macro- and micro-morphometry as well as on overall health and metabolic function (Shaik Mohamed Sayed et al., 2023). Current study findings showed that in female rats, treatment regimens did not affect GIT viscera macro-morphometry; a finding which suggests that the dietary insult (fructose drinking fluid), the conventional pharmacological agent (fenofibrate), and the proposed intervention (lycopene) neither compromised nor caused atrophy and/or hypertrophy of GIT viscera. Similarly, in males, the treatment regimens neither compromised nor caused atrophy and/or hypertrophy of the rats’ ceca and large intestines, as evidenced by similarities across treatment regimens. However, fenofibrate-treated males and lycopene (medium and high dose) supplemented counterparts had heavier stomachs compared with controls, and the fructose-fed males’ relative stomach masses were lower than their fenofibrate-treated counterparts. These findings suggest that treatment with fenofibrate and dietary fortification with lycopene at the medium and high dose could potentially enhance gastric function (increased mass), and importantly both fenofibrate and supplemental lycopene could be used to mitigate dietary fructose-mediated decrease in gastric mass. Treatment with fenofibrate and high-dose supplemental lycopene resulted in heavier small intestinal masses in males when compared with low-dose lycopene-supplemented counterparts. Furthermore fenofibrate-treated males had heavier relative small intestinal masses compared with controls. Taken together, these observations suggest that high-dose supplemental lycopene could be beneficial to the growth and development of males rats’ small intestines and that in fructose-fed males fenofibrate could also increase small intestinal mass, which is likely to increase the digestive and nutrient absorption function. The observed fenofibrate-mediated and supplemental lycopene-induced increased gastric and small intestine mass is an interesting finding. Fenofibrate increases plasma HDL-cholesterol concentration and decreases plasma triglyceride concentration, and together with other fibrates, it stimulates cellular fatty acid uptake, conversion to acyl-CoA derivatives, and nonesterified fatty acid catabolism via β-oxidation, which when combined with a reduction in fatty acid and triglyceride synthesis overall decreases VLDL production (Staels et al., 1998). These actions would decrease fat deposition in viscera including the stomach and small intestines and likely reduce their mass; thus, its observed mediation of increased gastric and intestinal mass requires further investigation. However, fenofibrate has been shown to reduce inflammation in various tissues (Jin et al., 2023), and similarly lycopene has been shown to mitigate ulcerative colitis and Crohn’s disease-induced inflammation (Głąbska et al., 2016; Chen et al., 2023). It can be speculated that through the anti-inflammatory activities both lycopene and fenofibrate may prevent tissue damage and promote gastric and small intestinal tissue integrity leading to an increase in mass. Lycopene has been touted to enhance nutrient absorption by modulating gut microbiota composition and activity, which translates to improved intestinal barrier function and upregulation of nutrient transporters (Meng et al., 2022; Tu et al., 2023). This improved nutrient absorption and gut microbiota can potentially contribute to improved nutrient by GIT that could translate to increased gastric and small intestine mass. We hypothesize that lycopene may promote the growth and regeneration of gastric and small intestinal mucosal cells, leading to the observed increased stomach and small intestine masses in the male rats. Hormones play a crucial role in GIT function regulation and tissue homeostasis (Norman and Henry, 2015; Farhadipour and Depoortere, 2021). Therefore, we speculate that lycopene may alter hormone signaling pathways involved in gastrointestinal health in a manner that promotes tissue growth and development. However, further research is needed to fully understand the specific mechanisms underlying this phenomenon.
CONCLUSIONS
Dietary lycopene supplementation reduced TFI in both male and female rats, but increased stomach and femora masses, as well as tibia length in males. These findings suggest that lycopene, when used as a dietary supplement, may enhance gastric function and support bone health in males, potentially protecting them from fenofibrate-induced bone loss. The observed improvements in male bone growth, specifically the increase in femora mass and tibia length, highlight the potential of lycopene as a natural intervention for managing diet-induced metabolic disturbances and promoting bone health, particularly in males.
Future studies should explore the mechanisms by which lycopene influences bone growth and metabolism, particularly its interaction with sex-specific hormones such as testosterone and estrogen. These findings carry biological significance, as lycopene, a naturally occurring antioxidant, could provide a dietary strategy for improving bone health and metabolic function. Lycopene supplementation may offer promise as a preventive approach for managing metabolic disorders, especially in populations at risk of osteoporosis and other bone-related conditions.
AUTHORS’ CONTRIBUTIONS
All authors contributed to the study’s conception and design. M.O.S. conducted the experiments, data collection, formal analysis, interpretation of results, and wrote the original drafts, with review and editing. N.M.G. and T.T.N. assisted with data collection and contributed to the review and editing. E.C. provided supervision, fund acquisition, and was involved in data collection, interpretation of results, validation, writing, and review and editing. All authors read and approved the final version.
Footnotes
ACKNOWLEDGMENTS
The authors thank the Wits School of Physiology and Bingham University in Nasarawa State, Nigeria, as well as the staff of the WRAF for their dedicated care of the rats throughout the study. The authors specially thank Kennedy Erlwanger, Bernice Asiedu, and Mmahiine Mosana for their invaluable technical assistance, and to all the students who contributed to the terminal procedures.
AUTHOR DISCLOSURE STATEMENT
The authors declare no competing interests relevant to this publication.
FUNDING INFORMATION
This research was supported by Research Incentive funding from the Faculty of Health Sciences, University of the Witwatersrand, Johannesburg, South Africa.
DATA AVAILABILITY STATEMENT
Data are provided within the article.