
Abstract
Bromelain is a mixture of enzymes found in several parts of the pineapple plant, including the stem, fruit, leaves, and peel. Its use was recommended to slow down the progression of neurological disorders such as Parkinson’s disease. Although studies have examined the beneficial effects of bromelain (Br) in neurological disorders, its detailed benefits in schizophrenia are still poorly understood. We, thus, modeled schizophrenia using two developmental methods. In the first experiment, postweaning social isolation (SI) method where male mice were weaned at postnatal day (PND) 21 and isolated in individual cage for 5 weeks was used to induce schizophrenia. The second experiment used a maternal deprivation (MD) model, where mothers were separated from their pups for 24 h at PND 9. In both experiments 1 and 2, Br was administered orally from PND 21 to PND 56. At PND 56, there were behavioral assessments of nest building, anxiety, depression, and locomotion, while oxidative stress, neuroinflammation, brain-derived neurotrophic factor, and astrocyte expression in the prefrontal cortex and hippocampus were assessed at the end of the behavioral studies. Our study found that both SI and MD caused anxiety, depression, and hyperlocomotion. This was accompanied by oxidative stress in the prefrontal cortex and hippocampus, and neuroinflammation was observed only in the hippocampus. While SI rearing caused poor nest-building capacity, MD rearing did not impact nest-building capacity of the mice. The study found that administration of 50 mg/kg of Br reversed oxidative stress and neuroinflammation and significantly reduced anxiety level, hyperlocomotion, and depression in the mice. Mice administered Br also had a better nest-building capacity. Br may, therefore, benefit individuals at ultra-high risk and patients with schizophrenia, alleviating the positive, negative, and cognitive symptoms of the disease, without the side effects reported in the use of psychotic drugs.
INTRODUCTION
Schizophrenia is a chronic mental disorder affecting 0.45% of the world population. 1 The symptoms of schizophrenia are broadly categorized into positive, negative, and cognitive symptoms. 2 Cognitive symptoms include deficits in executive control, working memory, and attention, while delusions and hallucinations form the positive symptoms. Examples of negative symptoms are anhedonia, deficits in social functioning, and avolition. This categorization is critical for proper diagnosis, treatment, and understanding of the disorder’s pathophysiology.
Both heritable and environmental factors are identified as risk factors for developing schizophrenia. Among the environmental factors are early-life adversity, 3 therefore, animal models such as postnatal social isolation (SI) and maternal deprivation (MD) have been successfully used to induce and study schizophrenia-related behaviors in rodents. 4 Currently, the etiology of schizophrenia is not clear, it is, however, argued that imbalance of the central immune system plays a major role in the pathogenesis of the disease. 5 Furthermore, astrocytes were proposed to play role in the pathogenesis of schizophrenia by producing components involved in inflammation and the functioning of interneurons. 6
Although there are pharmacological treatments available to ameliorate the positive symptoms, about 30% of patients give poor responses to these treatments. 3 In addition, the pharmacological treatments available produces undesirable effects such as hyperprolactinemia, extrapyramidal side effects, restlessness, and metabolic effects.7,8 These side effects may lead to poor medication adherence in patients. In developing economies, the cost of therapeutic management of schizophrenia may be unaffordable, and thus, herbal or nutritional remedies may provide an alternative yet effective means of preventing or managing schizophrenia.
Bromelain (Br) is a mixture of proteolytic enzymes 9 present in all parts of the pineapple plant. 10 Its anti-inflammatory and antianxiety functions have been reported, 11 while its use was also recommended to slow down the progression of Parkinson’s disease 12 and Alzheimer’s disease. 13 Traditionally, Br is used for bronchitis, angina pectoris, and debridement of wounds. 14 Considering the established involvement of oxidative stress, astrocytosis, and inflammation in the pathophysiology of schizophrenia, 15 as well as the demonstrated protective effects of Br in conditions such as depression, anxiety, and behaviors analogous to schizophrenia, Br may attenuate schizophrenia-associated behaviors in animal models of the disorder through its anti-inflammation, antioxidative effects, and other properties. Although studies have examined the beneficial effects of Br in neurological disorders, its detailed benefits in schizophrenia are still poorly understood.
Therefore, this study aims to examine the effects of Br on schizophrenia-associated behaviors using two distinct developmental models: the MD model and the SI model. Given the complex neurobiological mechanisms and diverse behavioral manifestations characteristic of schizophrenia, no single model fully captures the multifaceted nature of this disorder. Therefore, this investigation employs both MD and SI models to induce schizophrenia-related behaviors, allowing for a more comprehensive exploration of the condition. MD simulates early-life stress, which is a significant risk factor for schizophrenia. 16 Human studies show that adverse childhood experiences, such as neglect or abuse, increase the risk of developing the disorder. 17 Similarly, SI is a risk factor for schizophrenia-related behaviors in humans, and this model has been used successfully to induce schizophrenia in male BALB/c mice and other laboratory animals. 18 This evidence informed the decision to use the two models in this study.
The selection of male BALB/c mice for this study is appropriate due to specific traits that make them well-suited for investigating schizophrenia-related behaviors. BALB/c mice are known for their heightened sensitivity to environmental stressors, 19 which aligns with the stress–vulnerability hypothesis of schizophrenia. This strain exhibits behavioral symptoms such as heightened anxiety, social withdrawal, and impaired cognitive performance, which mirror core features of schizophrenia. Their extensive use in neurobehavioral research further enhances the reproducibility and relevance of the findings.
MATERIALS AND METHODS
Experimental animals and grouping
Experiment 1
Male BALB/c mice at postnatal day (PND) 21 were purchased from the animal house of University of Ilorin, Ilorin. They had free access to water and food. Animals were fed ad libitum and housed in perforated plastic cages. The mice were reared either in SI (one mouse/cage) or in groups (four mice/cage and three mice/cage) for 5 weeks. This experimental protocol lasted until the animals were used for behavioral and biochemical analyses.
Grouped-house group (control): seven male mice reared in groups.
SI group: seven male mice reared in isolation, i.e., one mouse/cage.
SI + Br group: seven male mice were reared in separate cages and administered 50 mg/kg of Br.
Experiment 2
Nine female mice and three male BALB/c mice were procured from the animal house of the University of Ilorin, Ilorin, Kwara State, Nigeria. The animals were grouped into three groups with a ratio of three female mice to one male mouse, given free access to food and water, and allowed to breed. Eight of the female mice gave birth to 57 pulps. MD was conducted on PND 9 by isolating the mother mice from their pups for a duration of 24 h, after which both the mother and pups were reunited in the same cage to resume breastfeeding. 20 After weaning on PND 21, mice were divided into three groups as follows:
Normal rearing group: seven male mice rearing without MD.
MD group: seven male mice with MD.
MD + Br group: seven male mice with MD with 50 mg/kg of Br administration from PND 21.
Experiment 2 lasted from 5 weeks, from PND 21 to PND 56.
Bakare and Owoyele 11 showed that 50 mg/kg of Br is the most effective dose at which Br exhibits anti-inflammatory and antioxidative properties. Br powder was procured from KAN Phytochemicals (Rai, India).
Animals (in both models) were given free access to water and food and maintained on a 12 h light–dark cycle. The animal house was under controlled humidity (45 ± 15%), controlled noise, and room temperature (27 ± 10°C). The research was approved by the University of Ilorin Ethical Review Committee, and all animal procedures were in compliance with the Guide for the Care and Use of Laboratory Animals.
Behavioral procedures
All behavioral assessments were conducted during the light cycle. The room designated for behavioral studies remained undisturbed by human presence throughout the procedures. The mice’s behavior during each test was recorded on video. Subsequently, the recorded footage was analyzed, and the animals’ behaviors were manually scored by an investigator who was blinded to their rearing conditions.
Nest-building test
The nest-building test was carried out as described by Nelly et al., 21 and their nest-building performance was scored on a scale of 1–5.
Open field test
Anxiety and locomotion of the mice were assessed using the open field test. The dimensions of the open field chamber used were 45 × 45 × 45 cm. The floor had 4 × 4 grid lines dividing it into equal squares. The animals were placed at the center of the chamber, and their movements were subsequently recorded for 6 min. Anxiety was assessed by measuring the time mice spent at the periphery, a measure known as thigmotaxis, 22 while locomotion was measured by recording the number of lines crossed.
Tail suspension test
Depression was measured in the mice using immobility time during the tail suspension test, following the procedure described by Can et al. 23
Preparation of brain tissues for biochemical assays
Brain tissues of mice from experiment 1 were collected after the mice were killed through cervical dislocation. The hippocampus and prefrontal cortex were obtained from the brain tissue and homogenized in phosphate buffer saline and centrifuged at 1000 × g for 10 min at 4°C. Brain-derived neurotrophic factor (BDNF), interleukin-1β (Il-1β), nuclear factor kappa-B (NF-κB), and tumor necrosis factor-α (TNF-α) concentration in the hippocampus and prefrontal cortex were determined using enzyme-linked immunosorbent assays kits. The kits were procured from Elabscience Biotech (TX, USA) with catalog numbers E-EL-R1235, E-EL-R0012, E-EL-R0674, and E-EL-R2856, respectively. The manufacturer’s protocols were followed strictly. The intensity of the color developed was measured using a spectrophotometer at 450 nm. Malondialdehyde (MDA) concentration was evaluated using the method described by Colado et al., 24 while colorimetric nonenzymatic assay procedure was used to determine nitric oxide (NO) concentration as described by Bryan and Grisham. 25
Preparation of brain tissue for immunohistochemistry
The expression of astrocyte in the hippocampus and prefrontal cortex was quantified and qualified using glial fibrillary acidic protein (GFAP) staining procedure. Mice used for the immunohistochemistry procedure were perfused using phosphate-buffered saline and then 10% formal saline. Perfusion was done transcardially. The whole brain was then harvested, while the two brain tissues were separated and fixed in 10% formal saline. Immunohistochemical staining procedure was used to determine astrocyte expression using mouse polyclonal rabbit anti-mouse GFAP primary antibody. The images were quantified for staining intensity using the open source Fiji.
Statistical analysis
The results obtained were expressed as mean ± standard error of the mean. Data were analyzed with GraphPad version 8.0.2 (GraphPad Software Inc., CA, USA) using a one-way analysis of variance followed by the least significant difference post hoc test. The significant level was set at P < 0.05. SI group was compared with control group 1, and the SI + Br group was compared with the SI group. Furthermore, MD group was compared with the normal rearing group, and the MD + Br group was compared with the MD group.
RESULTS
Effect of bromelain on nest-building test
Tables 1 and 2 show the effect of Br on the nest-building test in SI-induced schizophrenia. The SI group ranked low on the nest-building test (2.63 ± 0.24), which was reversed by Br administration (4.0 ± 0.46). However, MD rearing did not significantly affect nest-building capacity.
Effects of Bromelain on Nest-Building Test in Social Isolation Model

aSignificant difference from the control group at P < .05.
bSignificant difference from the SI + Br group at P < .05.
Br, bromelain; SEM, standard error of the mean; SI, social isolation.
Effect of Bromelain on Nest-Building Test in Maternal Deprivation Model

MD, maternal deprivation.
Effect of bromelain on line crossing
SI rearing led to a significant increase in line crossing (P < .05) in the SI group (217.75 ± 9.57) compared to the control group (154.50 ± 7.60), as shown in Table 3. Likewise, MD caused a significant increase in line crossing (P < .05) as shown in Table 4. However, this effect of rearing condition was reversed by Br administration, as line crossing (indicating locomotive activity) decreased significantly in the SI + Br group (177.25 ± 8.51) compared to the SI group, and in the MD + Br group (145.0 ± 4.58) compared to the MD group (218.75 ± 6.85).
Effects of Bromelain on Number of Lines Crossed in Social Isolation Model

aSignificant difference from the control group at P < .05.
bSignificant difference from the SI + Br group at P < .05.
Effects of Bromelain on Number of Lines Crossed in Maternal Deprivation Model

aSignificant difference from the control group at P < .05.
bSignificant difference from the SI + Br group at P < .05.
Effect of bromelain on tail suspension test
As shown in Tables 5 and 6, SI led to a significant increase (P < .05) in immobility time compared to the control group. However, immobility time was significantly reduced (P < .05) in the group administered Br (118.0 ± 6.18 sec) compared to the SI group (195.0 ± 13.74 sec). Likewise, in MD rearing, there was a significant increase in immobility time in the MD group (183.0 ± 5.1 sec) compared to the control group (139.3 ± 17.1 sec), an effect that was reversed by Br administration in the MD + Br group (107.25 ± 2.40 sec). Hence, administration of Br reversed depression-like symptom in both models of schizophrenia.
Effects of Bromelain on Immobility Time in Social Isolation Model

aSignificant difference from the control group at P < .05.
bSignificant difference from the SI + Br group at P < .05.
Effects of Bromelain on Immobility Time in Maternal Deprivation Model

aSignificant difference from the control group at P < .05. b Significant difference from the SI + Br group at P < .05.
Effect of bromelain on anxiety
Anxiety was assessed by measuring the time spent at the periphery (thigmotaxis). Both SI rearing and MD rearing caused a significant increase in the time spent at the periphery. However, Br administration was able to significantly reduce this in both models of schizophrenia (Tables 7 and 8).
Effects of Bromelain on Anxiety in Social Isolation Model
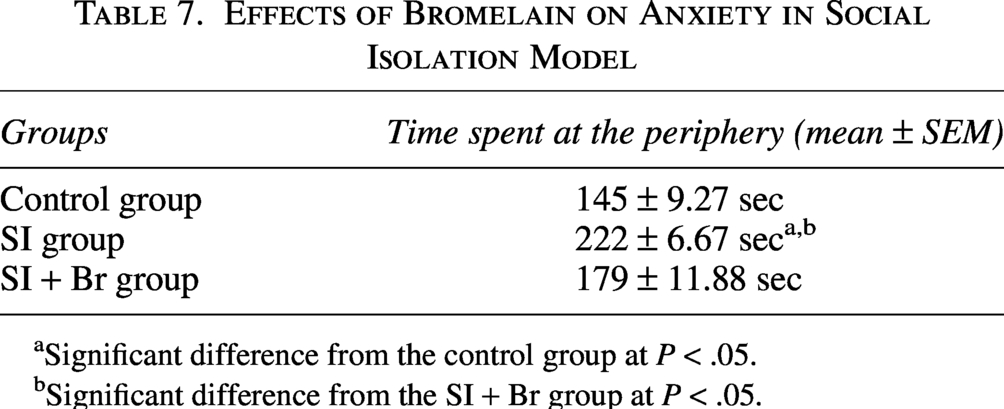
aSignificant difference from the control group at P < .05.
bSignificant difference from the SI + Br group at P < .05.
Effects of Bromelain on Anxiety in Maternal Deprivation Model
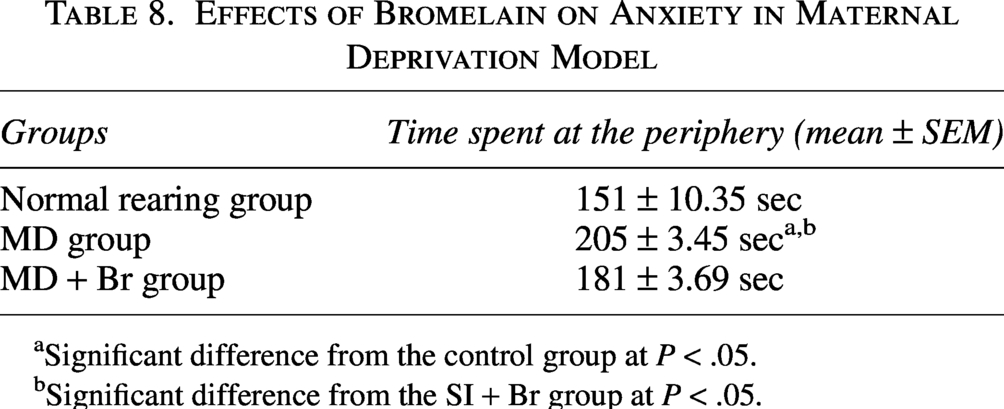
aSignificant difference from the control group at P < .05.
bSignificant difference from the SI + Br group at P < .05.
Effect of bromelain on oxidative stress in the prefrontal cortex and hippocampus
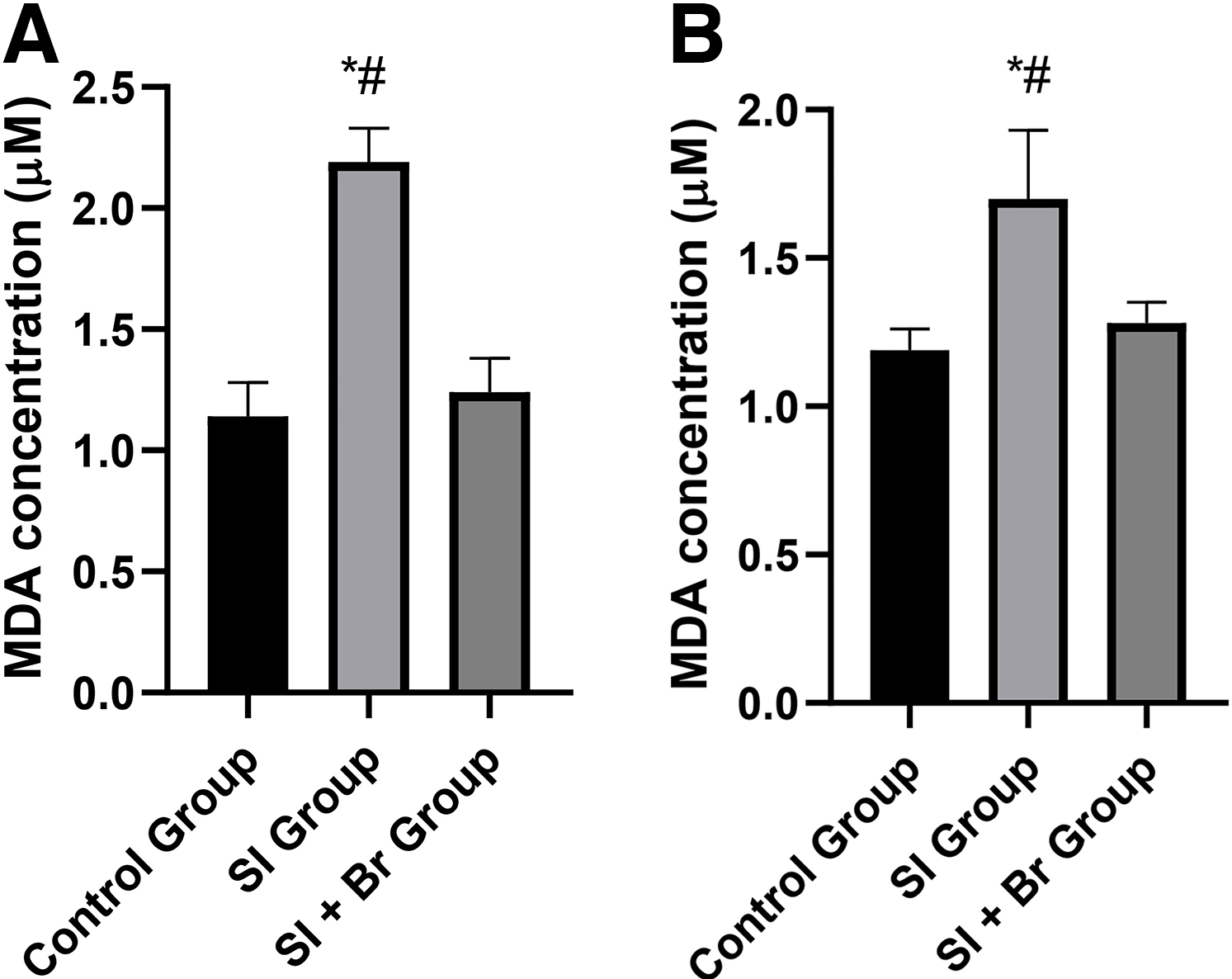
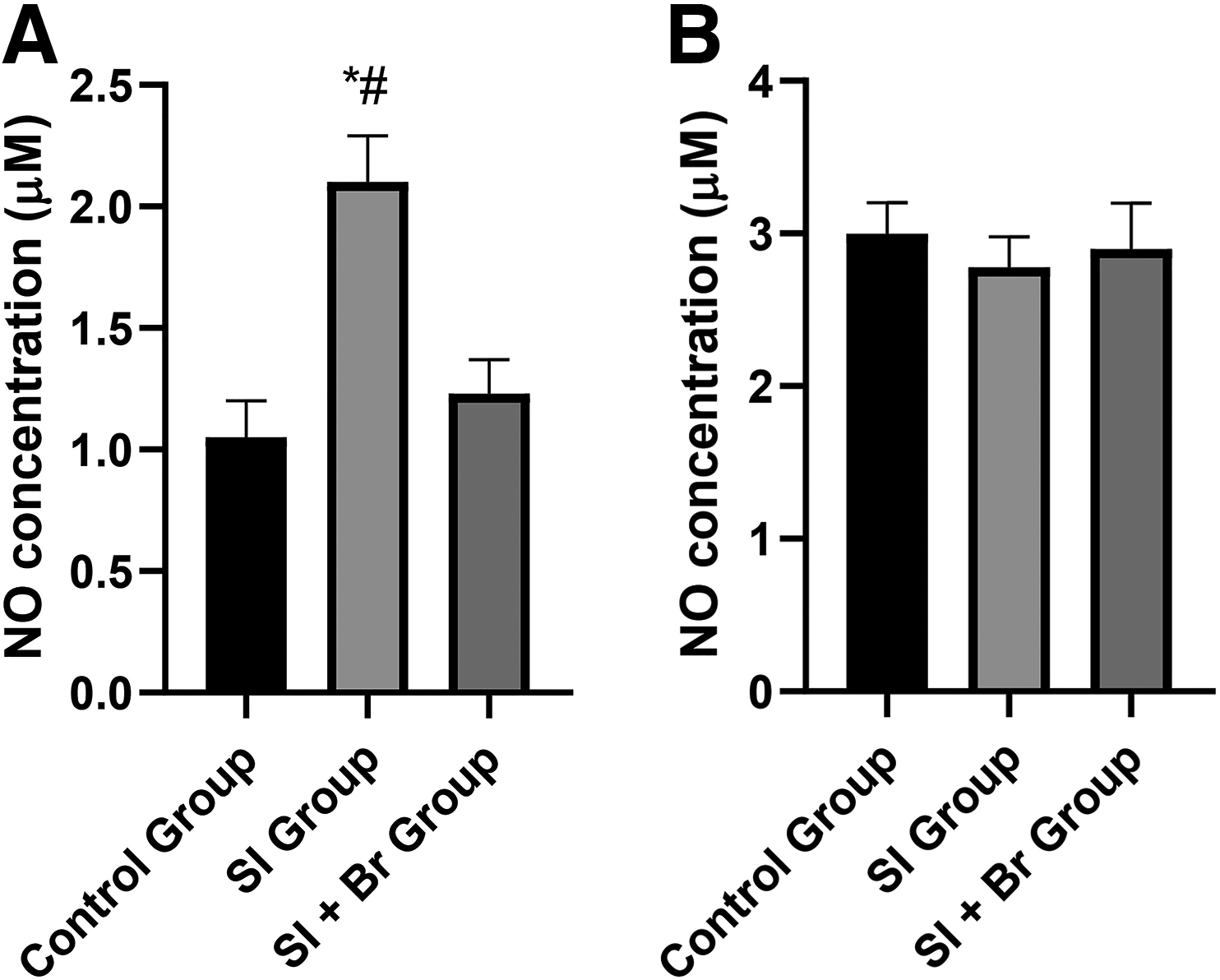
In the prefrontal cortex, there was a significant increase (P < .05) in MDA concentration in the SI group (2.19 ± 0.14 µM) compared to the control group (1.14 ± 0.14 µM). Nonetheless, administration of Br led to a significant decrease (P < .05) in the concentration of MDA (Fig. 1A). This same observation was made in the hippocampus, where there was a significant increase in MDA concentration in the SI group (P < .05) compared to both the control group and SI + Br group (Fig. 1B).
NO concentration was assessed in both the prefrontal cortex and hippocampus (Fig. 2). In the prefrontal cortex, SI led to a significant increase (P < .05) in nitric oxide concentration (2.19 ± 0.14 µM) compared to the control group, which was reversed in the SI + Br group (1.23 ± 0.14 µM). However, in the hippocampus, there was no difference in NO concentration across the three groups.
Effect of bromelain on neuroinflammation in the prefrontal cortex and hippocampus
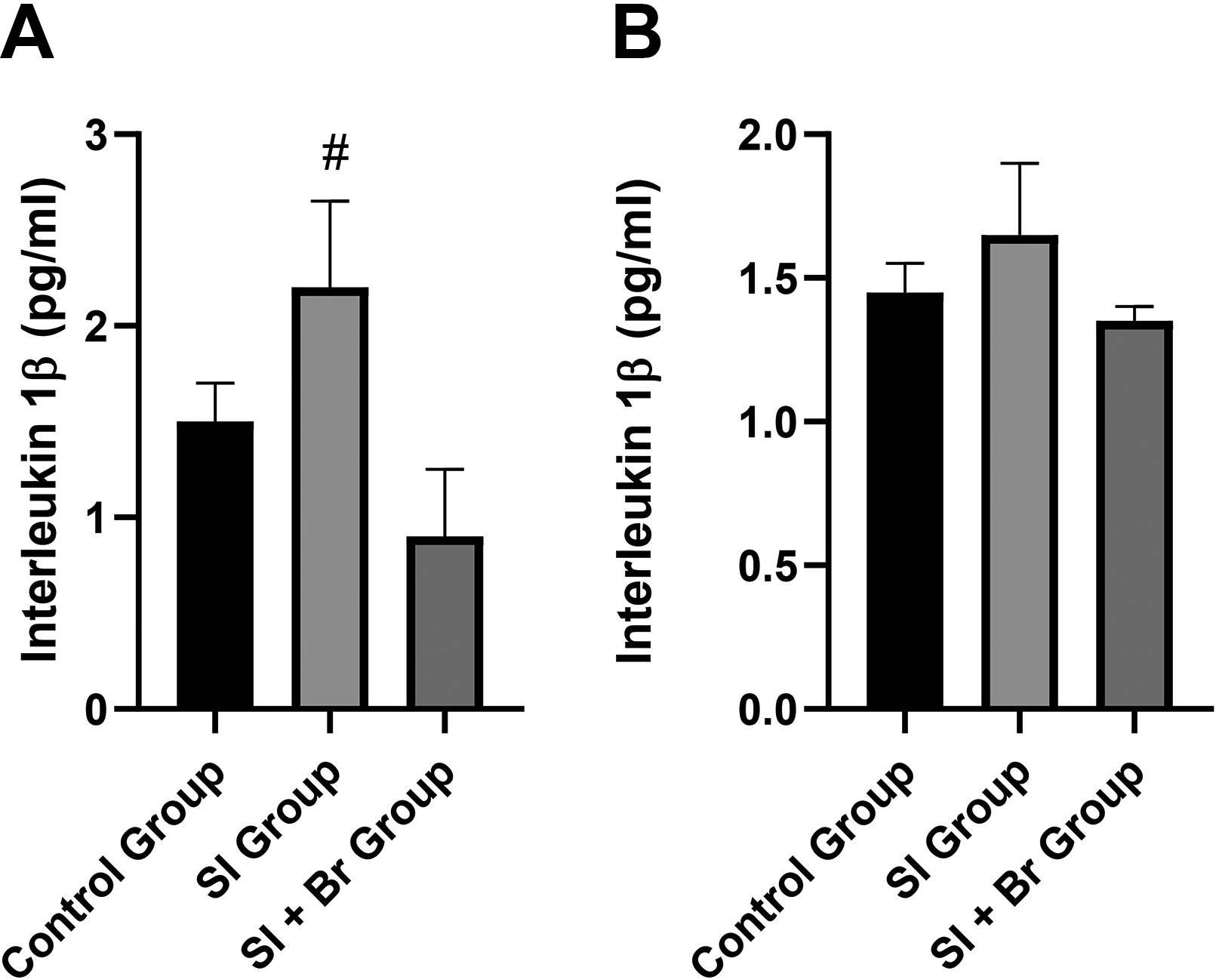
IL-1β, NF-κB, and TNF-α were assessed in the prefrontal cortex and hippocampus. In the prefrontal cortex, there was a nonsignificant increase in IL-1β concentration in the SI group compared to the control group (Fig. 3A). However, administration of Br led to a significant decrease (P < .05) in IL-1β concentration compared to the SI group. Similarly, in the hippocampus, SI led to a significant increase (P < .05) in the IL-1β concentration in the SI group compared to the SI + Br group (Fig. 3B).
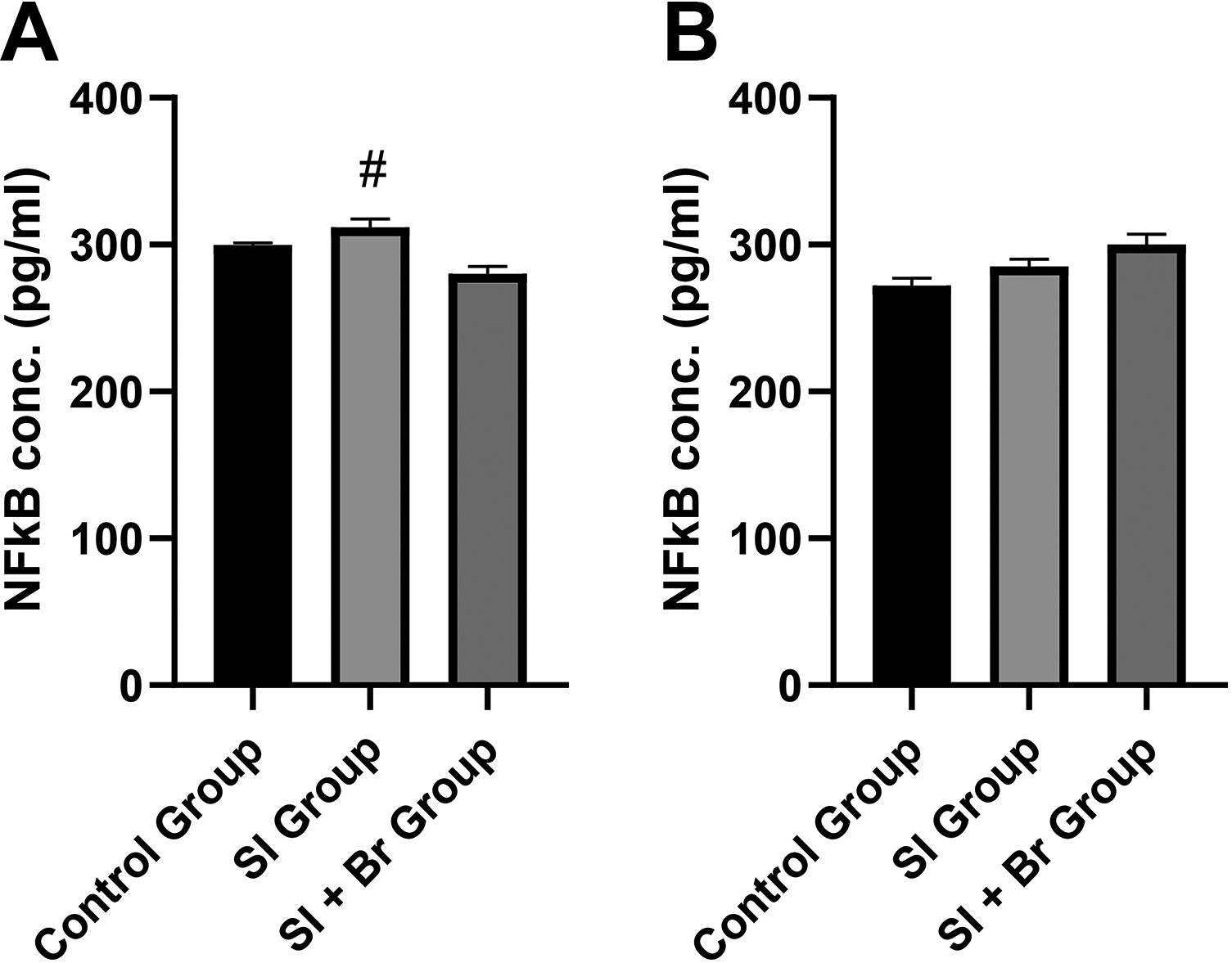
In the prefrontal cortex, SI led to a nonsignificant increase in the concentration of NF-κB (311.92 ± 5.59 pg/mL) compared to the control group (299.85 ± 1.41 pg/mL). However, there was a significant decrease in NF-κB concentration in the SI + Br group (Fig. 4) compared to both the SI and control groups (P < .05). This indicates that administration of Br is able to reverse neuroinflammation in the prefrontal cortex. However, in the hippocampus, Br administration was unable to reverse neuroinflammation.
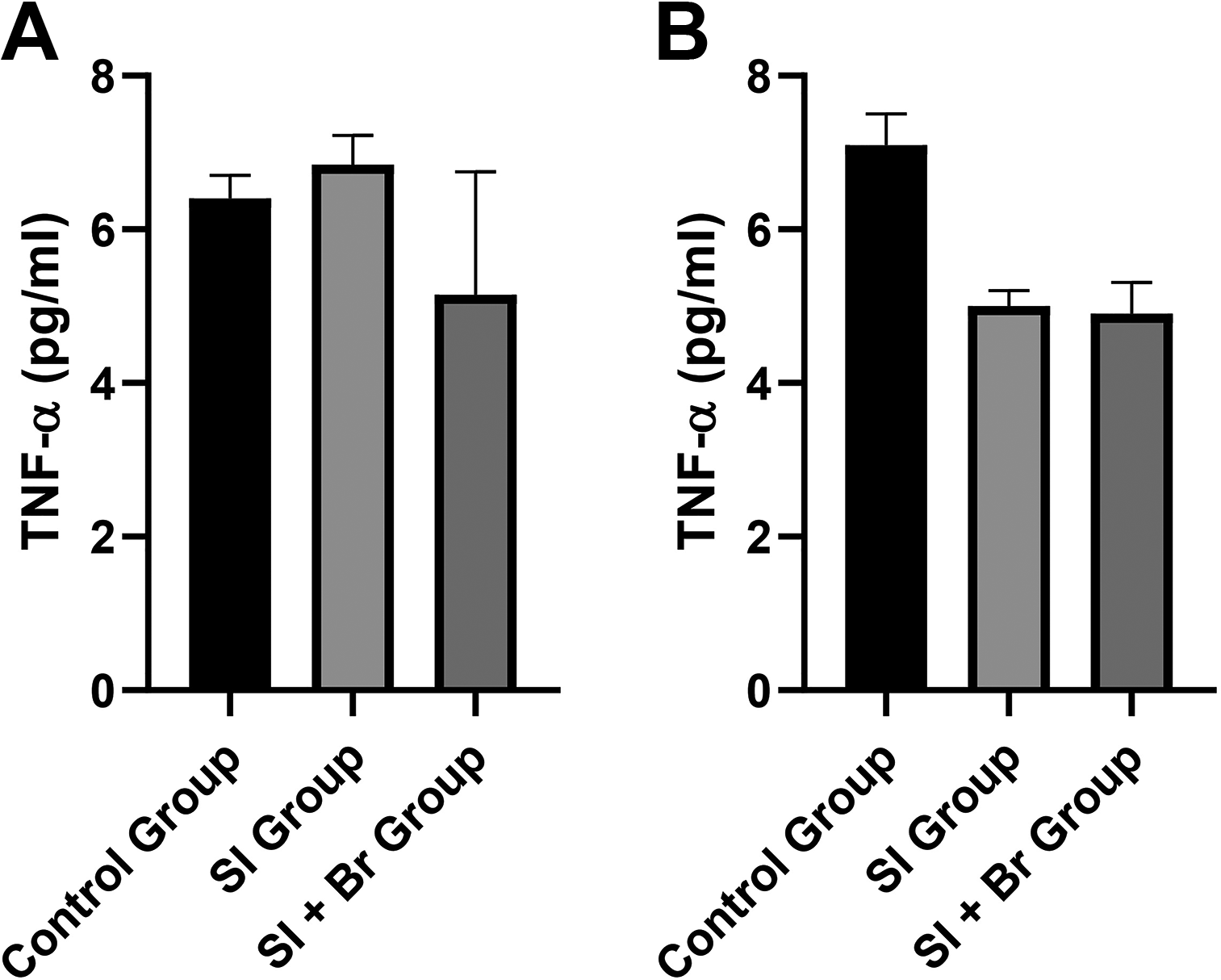
Although there appears to be an increase in TNF-α concentration in the prefrontal cortex of the SI group (Fig. 5), this was not significant compared to the control and SI + Br groups. Furthermore, both the SI and SI + Br groups showed a significant decrease in TNF-α concentration in the hippocampus.
Effect of bromelain on BDNF
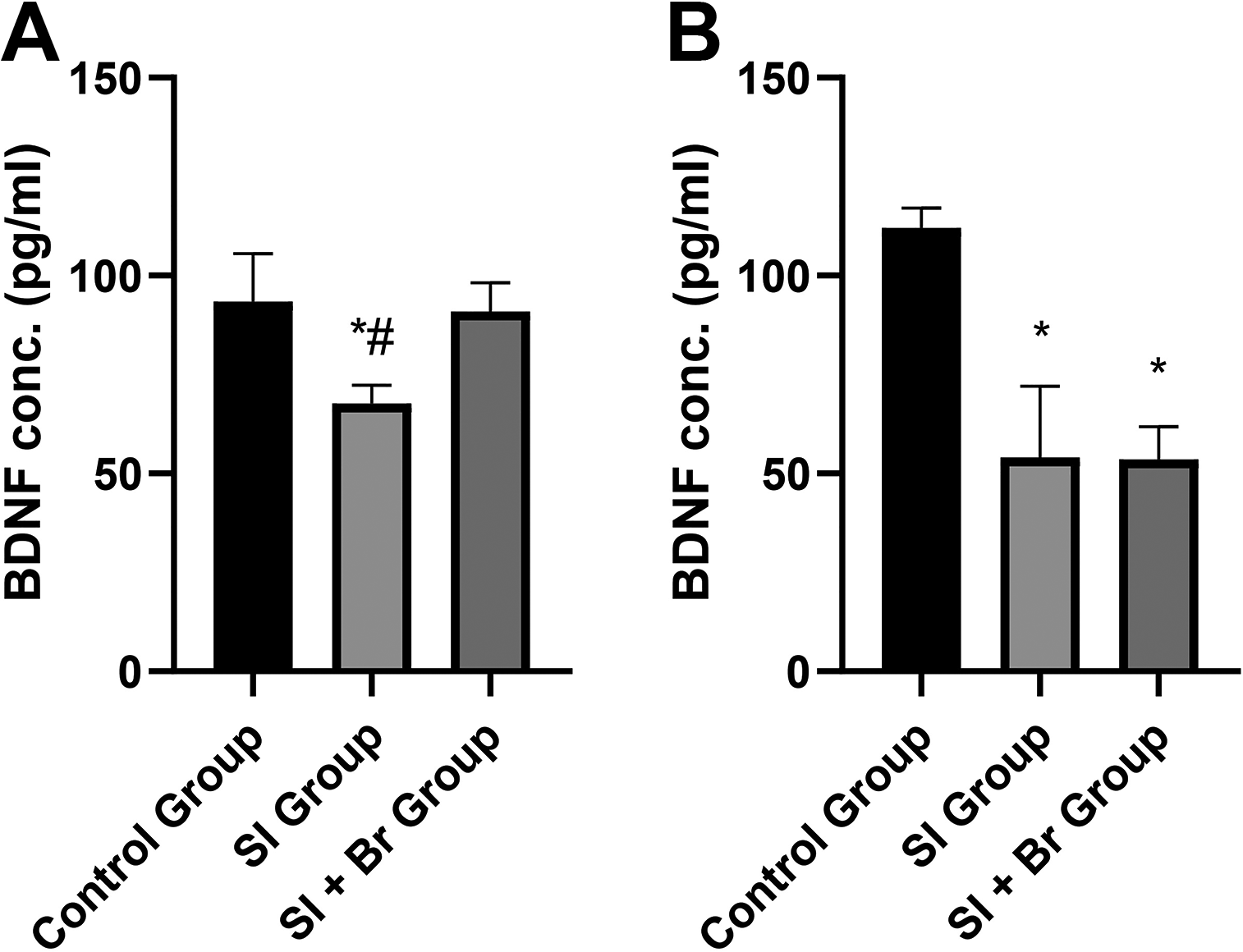
Figure 6 shows the effect of Br administration on SI-induced schizophrenia-related behaviors. There was a significant decrease (P < .05) in BDNF concentration in the prefrontal cortex in the SI group (67.73 ± 4.63 pg/mL) compared to the control group (93.38 ± 12.24 pg/mL). However, administration of Br was able to reverse this decrease in the BDNF concentration (90.99 ± 7.17 pg/mL) caused by SI. In the hippocampus, there was a significant reduction (P < .05) in BDNF concentration in both the SI and SI + Br groups compared to the control group. In other words, the administration of Br was unable to reverse the decrease in BDNF concentration caused by SI in the hippocampus.
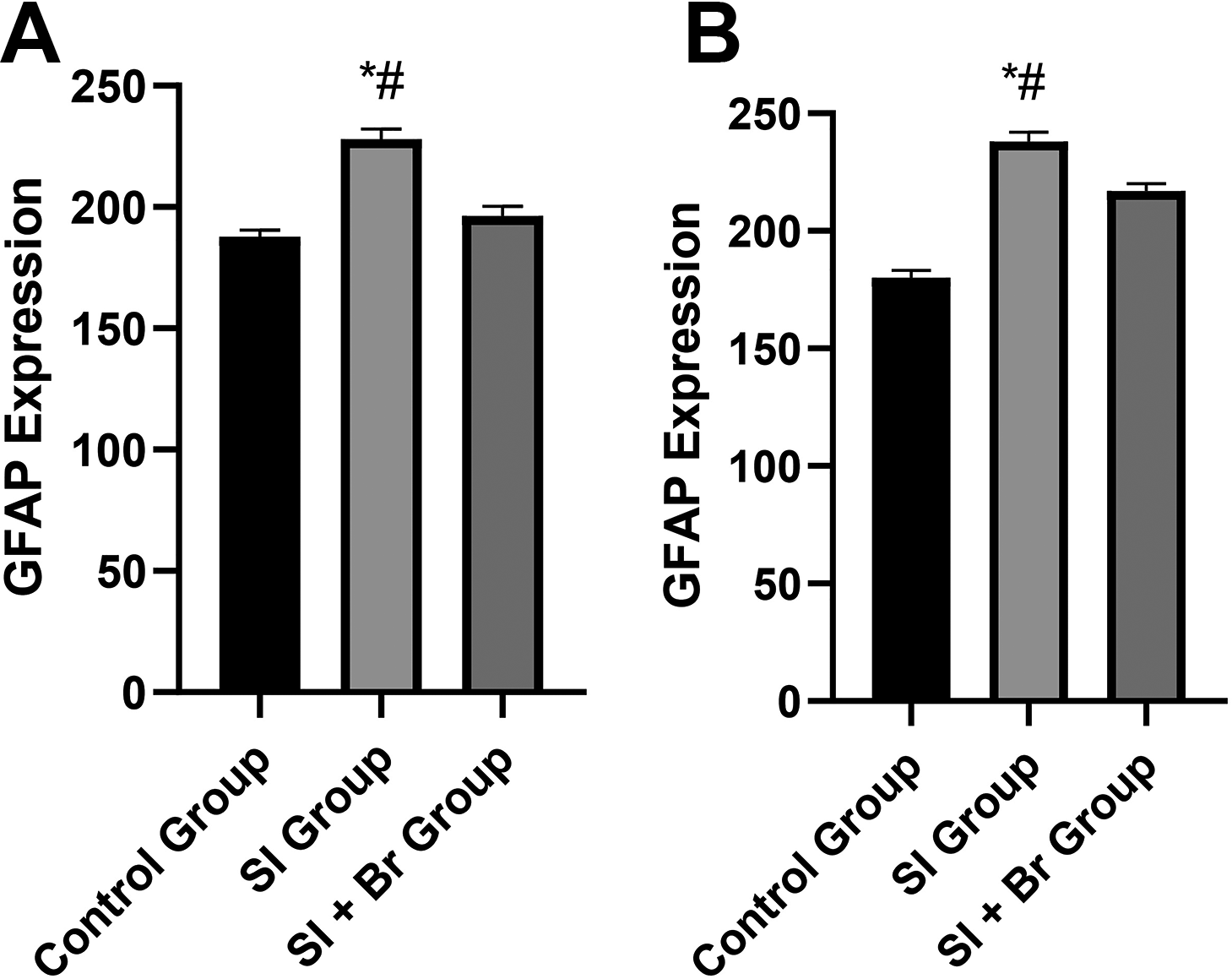
Effect of bromelain on astrocytes expression
Figure 7 shows the effect of SI and Br on GFAP expression in the astrocytes. In the SI group, there was a significant increase (P < .05) in the GFAP expression (227.93 ± 4.36) compared to the control group (187.88 ± 2.66) in the prefrontal cortex. However, in the SI + Br group, there was a significant reduction (P < .05) in GFAP expression (196.31 ± 4.02) compared to the SI group. The same pattern was observed in the hippocampus. While SI increased GFAP expression significantly, the administration of Br significantly reversed this effect.
DISCUSSION
This study shows the effects of Br on schizophrenia-related behaviors in SI and MD rearing models. The effect on anxiety, depression, locomotive activity, and nest-building test were found to be beneficial. In addition, Br was able to reverse oxidative stress and neuroinflammation caused by SI, while also improving the production of BDNF in the prefrontal cortex. This study found that Br reduced the GFAP expression in the astrocytes, an effect that might have caused the reduced neuroinflammation and oxidative stress.
The nest-building test has been used to assess moods, general well-being, sensorimotor impairments, and disease progression in psychiatric disorders. 26 Barkus et al. 27 indicated that there is a decrease in nest-building capacity in animal models of schizophrenia. In this study, SI led to mice’s poor performance in nest building, which was reversed by the Br administration. This indicates that Br was able to improve the mood and the general well-being in the mice. However, MD had no effect on nest-building capacity of the mice.
This study also observed heightened anxiety in the SI group and MD groups, as indicated by increased time spent at the periphery in the open field test. Administration of 50 mg/kg of Br was able to significantly reduce anxiety in the mice, as indicated by the reduced time spent at the periphery in both the SI + Br and MD + Br groups. Bakare and Owoyele showed earlier that administration of 50 mg/kg of Br in rats showed antianxiety effect, an effect mediated through its anti-inflammatory property. 11 Hence, the administration of Br may alleviate anxiety in schizophrenia and other psychiatric disorders.
SI has earlier been shown to produce depression,28,29 an important symptom of schizophrenia. Similarly, Khalifeh et al. 30 showed that MD rearing can induce depression in mice. Hence, the treatment of depression is important in the management of the disease. In this study, Br administration was found to reverse depression, as observed in the tail suspension test, in both MD and SI rearing models.
In this study, 50 mg/kg of Br was able to reverse hyperlocomotion caused by SI and MD rearing. Previous studies have established that both SI and MD lead to increased locomotor activity.31–33 Hyperlocomotion in rodents is considered a correlate of the positive symptoms of schizophrenia. 34 Hence, Br may therefore be beneficial in alleviating the positive symptoms of schizophrenia.
This study found that postweaning SI led to increased oxidative stress in the prefrontal cortex and hippocampus. The increased NO is able to provoke nitrosative stress, leading to activation of pro-apoptotic signaling and other debilitating effects on the brain. 35 In this study, Br reversed oxidative stress by reducing NO production in the prefrontal cortex. This study, however, did not find any significant difference in NO production in the hippocampus across the three groups. Similarly, MDA concentrations in both the prefrontal cortex and hippocampus were increased in the SI group but were reversed by Br. A study by Saptarini et al. 36 proved the antioxidative activity of Br. Br, being a thiol endopeptidase enzyme, may also exhibit its antioxidant property through its other components such as peroxidase and protease inhibitors.37,38 Therefore, Br might exert its effect on schizophrenia-related behaviors, through its antioxidant property.
Many studies have established the involvement of neuroinflammation in the pathophysiology of SI-induced schizophrenia-related behaviors.18,39 On the contrary, Du Preez et al. 40 found no altered neuroinflammation in SI. Furthermore, human studies that assessed the beneficial impact of nonsteroidal anti-inflammatory drugs have conflicting outcomes.41,42 In summary, there are still arguments on the role of neuroinflammation in the pathophysiology of schizophrenia. In this study, however, there was an increase in Il-1β and NF-κB only in the prefrontal cortex, but not in the hippocampus. Conversely, there was a significant decrease in TNF-α concentration in the hippocampus of the SI and SI + Br groups compared to the control group. Nonetheless, Br administration was able to significantly reduce the concentration of the neuroinflammatory markers in the prefrontal cortex, further confirming the anti-inflammatory effect of Br. 43 Elevation of Il-1β was reported in the cerebrospinal fluid of male patients with schizophrenia and was linked to the negative and cognitive symptoms of the disease, 44 while overexpression of NF-κB was linked to depressive-like behaviors. 45 Br, therefore, was able to improve nest-building score and immobility time in the mice, by regulating IL-1β and NF-κB.
BDNF is involved in brain development and functioning, and postweaning SI has been known to cause a reduction in BDNF expression in rodents.46,47 In this study, there was a significant reduction in BDNF concentration in both the prefrontal cortex and the hippocampus. However, Br administration increased BDNF levels only in the prefrontal cortex, not in the hippocampus. Individuals with an ultra-high risk of developing schizophrenia, as well as patients with first-episode psychosis, have been reported to exhibit lower levels of BDNF.48–50 These reduced BDNF levels have been linked to the cognitive symptoms of schizophrenia. 51 Hence, our findings suggest that such populations may benefit from Br administration at 50 m/kg.
This study found a significant increase in the expression of GFAP, an outcome confirmed in animal and human studies.52,53 The overactivation of astrocytes in schizophrenia is proposed to alter the extracellular matrix system, ultimately leading to dysfunction of neurons and inflammation. 6 The dysfunction in the neuronal activities may be reflected in the reduced BDNF and increased oxidative stress observed in the SI group in this study. However, our study also found that bromelain administration was able to reverse astrocyte hyperactivation caused by SI.
Many novel drugs identified to be effective in animal models of schizophrenia have failed in clinical trials. This may be because the animal models used to assess drug efficacy are not severe enough, or only few animal models were used to test the efficacy of these novel drugs. 54 We have earlier established that both male and female mice develop schizophrenia-like behaviors during SI rearing. 55 However, our earlier study only relied on one model to prove the efficacy of Br in schizophrenia. 55 The current study has thus established that Br can prevent the development of schizophrenia-like behaviors in two different animal models of early-life stress—MD and SI. Our future studies will explore other models such as genetic and pharmacological model to further establish the effectiveness of Br.
CONCLUSION
SI and MD rearing caused schizophrenia-related behaviors such as anxiety, depression, and hyperlocomotion, which were reversed by Br administration. Notably, MD rearing did not produce poor nest-building capacity. The schizophrenia-related behaviors observed in this study were accompanied by oxidative stress in the prefrontal cortex and hippocampus, as well as neuroinflammation. Administration of 50 mg/kg of Br was able to reverse both oxidative stress and neuroinflammation. Furthermore, Br improved BDNF concentration and may therefore be beneficial in improving the cognitive symptoms of schizophrenia. In conclusion, Br may benefit individuals at ultra-high risk and patients with schizophrenia by alleviating the positive, negative, and cognitive symptoms of the disease without the side effects reported in the use of psychotic drugs.
AUTHORS’ CONTRIBUTIONS
G.O.A.: Conceptualization, methodology, investigation, formal analysis, data curation, and writing of article. B.V.O.: Writing—review and editing and supervision. Both authors read and approved the final article.
COMPLIANCE WITH ETHICAL STANDARDS
This study was authorized by the University of Ilorin Ethical Review Committee.
Footnotes
ACKNOWLEDGMENT
The authors acknowledge Samuel Oluwasanmi, Ishmael Adeniyi, and Soji Olajide for the support provided during the course of the research work.
AUTHOR DISCLOSURE STATEMENT
The authors declare no conflict of interest.
FUNDING INFORMATION
This research was self-funded by both authors.