
Abstract
Diarrhea is characterized by excessive intestinal secretion and motility, often linked to inflammatory activation. Although synthetic antidiarrheal drugs are effective, their prolonged use can cause side effects, underscoring the need for safe, natural alternatives.
This study investigated the antidiarrheal and anti-inflammatory activities of Bacillus subtilis DKU_09-fermented soybean (FS) powder in a mouse model of castor oil-induced diarrhea.
Male ICR mice were orally administered FS (100, 200, or 300 mg/kg) for 7 days before the induction of diarrhea with castor oil. Diarrheal onset, frequency, fecal water content, and gastrointestinal (GI) transit were recorded. Inflammatory mediators in the small intestine were analyzed using real-time polymerase chain reaction and Western blotting, while serum immunoglobulin A (IgA) and immunoglobulin G (IgG) were measured by enzyme-linked immunosorbent assay.
Pretreatment with FS significantly delayed the onset of diarrhea (FS200: 132.0 ± 9.8 min vs. castor oil-treated control [DC]: 76.9 ± 5.6 min, P < .05) and reduced diarrheal frequency (FS200: 32.9 ± 5.4% vs. DC: 51.9 ± 6.1%, P < .05). FS normalized the accelerated GI transit and markedly suppressed intestinal inflammation. The expression of Il-1β, Cox-2, mPGES-1, and NF-κB (p65) mRNA was significantly decreased by FS, accompanied by inhibition of mitogen-activated protein kinase (MAPK) phosphorylation (ERK, JNK, and p38). Serum IgG and IgA levels were unaffected, indicating that the effects were localized to intestinal tissue.
Soybeans fermented by B. subtilis DKU_09 exert potent antidiarrheal effects through suppression of the NF-κB/Cox-2/PGE2 and MAPK signaling pathways, thereby reducing intestinal inflammation and motility. These findings suggest that the FS used in the present study may serve as a safe and natural functional food for the management of acute diarrhea.
INTRODUCTION
Diarrhea is one of the most common gastrointestinal (GI) disorders worldwide and remains a significant cause of morbidity and mortality, particularly in developing countries. 1 Acute diarrhea is characterized by increased stool frequency and water content resulting from intestinal inflammation, increased mucosal permeability, and dysregulated electrolyte transport. 2 Pharmacological management of diarrhea commonly involves agents such as loperamide, which acts on μ-opioid receptors to suppress intestinal motility. 3 However, long-term or excessive use of synthetic antidiarrheal drugs may cause adverse effects, including constipation, abdominal discomfort, and alterations in the gut microbiota.4,5 Consequently, there is growing interest in developing safe, natural, and microbiota-friendly functional foods with antidiarrheal and anti-inflammatory activities. 6
To evaluate potential antidiarrheal compounds, the castor oil-induced diarrhea model in rodents is widely utilized.7,8 Castor oil is metabolized by intestinal lipases into ricinoleic acid, which exerts its powerful laxative effect through two primary mechanisms.9,10 First, ricinoleic acid directly stimulates intestinal smooth muscle, accelerating GI transit. Second, it induces intestinal secretion by increasing mucosal permeability and stimulating the release of inflammatory mediators. This secretory component is closely linked to activation of the NF-κB signaling pathway and the subsequent upregulation of pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α. 11 Critically, this pathway also leads to increased expression of inflammatory enzymes such as cyclooxygenase-2 (COX-2) and microsomal prostaglandin E2 synthase-1 (mPGES-1), resulting in increased production of prostaglandin E2 (PGE2), a potent secretagogue that drives fluid accumulation and diarrhea. 12 Therefore, modulating the inflammatory cascade, particularly the NF-κB/COX-2/PGE2 axis, presents a viable strategy for novel antidiarrheal development.
Soybeans (Glycine max) have long been a staple food and are renowned for their nutritional value, containing proteins, peptides, and various bioactive compounds such as isoflavones (daidzein, genistein), which possess anti-inflammatory and antioxidant properties.13,14 Fermentation technology, especially using beneficial microorganisms like Bacillus subtilis, significantly enhances the bioavailability and potency of these compounds by converting complex molecules into simpler, more active forms. B. subtilis-fermented soybean (FS) products, such as cheonggukjang, are traditionally recognized as health-promoting functional foods in East Asia and are known sources of potent probiotics and prebiotics that can restore the balance of the gut microbiota.
Given the potential of B. subtilis-FS powder to improve gut health and its well-documented anti-inflammatory effects, we hypothesized that FS could exert significant protective activity against secretory diarrhea. B. subtilis is widely used in soybean fermentation because it converts complex macromolecules into bioactive forms, thereby enhancing the nutritional and functional properties. The strain B. subtilis DKU_09, isolated from traditional Korean cheonggukjang, was selected as the fermentation starter for this study because it is a well-characterized, food-grade microorganism with stable growth and spore-forming capacity under aerobic conditions. It has been safely used in previous food fermentation research and is recognized for its consistent fermentation performance. Based on these characteristics, DKU-09 was considered suitable for producing FS powder with improved functional potential. 15
Therefore, this study aimed to investigate the antidiarrheal effects of FS fermented with the B. subtilis DKU_09 strain in a mouse model of castor oil-induced diarrhea. Specifically, we examined whether FS could alleviate diarrhea-related symptoms (onset time, frequency, and GI transit) and explored its underlying mechanism of action by analyzing the suppression of intestinal inflammation, focusing on the gene expression of pro-inflammatory cytokines (IL-6, IL-1β, and TNF-α) and key enzymes of the prostaglandin pathway (NF-κB p65, COX-2, and mPGES-1) in small intestinal tissues.
MATERIALS AND METHODS
Isolation and identification of DKU_09
The B. subtilis DKU_09 strain was obtained from cheonggukjang and cultivated in Tryptic Soy Broth (BD Bioscience, NJ, USA) at 40°C with continuous agitation at 150 rpm for 24 h. Subsequently, a single colony was isolated by streaking onto Tryptic Soy Agar (TSA; BD Bioscience, NJ, USA). For taxonomic identification, genomic DNA was extracted from the isolated colony, and the 16S rRNA gene was amplified and sequenced using the universal primer sets (5′-AGA GTT TGA TCM TGG CTC AG-3′), 1492R (5′-TAC GGY TAC CTT GTT ACG ACT T-3′), 785F (5′-GGA TTA GAT ACC CTG GTA-3′), and 907R (5′-CCG TCA ATT CMT TTR AGT TT-3′) by Macrogen (Seoul, Korea). The obtained sequences were compared with reference sequences in the National Center for Biotechnology Information database using the Basic Local Alignment Search Tool to determine sequence similarity. The B. subtilis DKU_09 strain has been deposited in the Korean Collection for Type Cultures (KCTC, Korea) under accession number KCTC 15693BP, in accordance with the Budapest Treaty. The genome sequence of the strain is also available in the National Center for Biotechnology Information under accession number NZ_CP141283.1 (assembly no. ASM3531960v1).
Preparation of fermented soybean
Soybeans were pretreated by soaking at 4°C for 8 h, followed by sterilization at 121°C for 1 h. The sterilized soybeans were inoculated with the DKU_09 starter culture and incubated at 37°C for 24 h. This temperature was selected as it is optimal for both microbial growth and enzymatic activity during fermentation, facilitating the bioconversion of functional compounds. After fermentation, the samples were sequentially dried at 42°C for 48 h and then at 70°C for an additional 48 h to ensure efficient moisture removal while minimizing degradation of bioactive components. To evaluate viable bacterial counts, the FS powder was diluted to 108 CFU/mL, plated on TSA, and incubated at 37°C for 20 h, after which colony-forming units were determined.
Morphological characteristics of the DKU_09 strain, including endospore formation during fermentation, were previously examined using scanning electron microscopy in our earlier study. Furthermore, the bioactive compound profile of the FS powder was previously characterized using high-performance liquid chromatography (HPLC), as described in our earlier study. 15 In that study, dried powder samples were extracted with 70% ethanol and analyzed by HPLC using a UV detector and a C18 reversed-phase column to quantify major isoflavones, including daidzein, genistein, daidzin, and genistin. Detailed chromatographic conditions and quantitative results have been reported previously. 15 Therefore, in the present study, HPLC analysis was not repeated, and compositional interpretation was based on the previously validated data.
Animals, oral administration, and antidiarrheal activity model
ICR male mice (6 weeks old, 25–30 g) were purchased from Raonbio (Yongin, Korea). Following a 1-week acclimatization period under controlled conditions (temperature: 21 ± 2°C, humidity: 50 ± 5%), the mice were housed individually with ad libitum access to standard chow and water. The experimental protocol was approved by the Dankook University Institutional Animal Care and Use Committee (DKU-25-039).
The mice were randomly assigned to six groups: normal control (NC, n = 12), castor oil-treated control (DC, n = 12), loperamide-treated group (LOP, 3 mg/kg, n = 12), and FS-treated groups (FS100, 100 mg/kg, n = 10; FS200, 200 mg/kg, n = 10; FS300, 300 mg/kg, n = 10). FS was dissolved in saline and administered orally once daily for 1 week prior to the induction of diarrhea.
Before the experiment, all mice were fasted for 6 h. One hour after oral administration of the vehicle, extract, or positive control, single dose of castor oil (0.3 mL) was administered by gavage to each mouse, except those in the normal control group. All mice were then placed in individual cages to evaluate antidiarrheal activity.
Briefly, after castor oil administration, the animals were observed for defecation for up to 4 h. Filter paper was placed beneath each cage to observe the characteristics of diarrheal droppings. During the 4 h observation period, the onset time of diarrhea, total fecal output, and fecal water content were recorded for an individual mouse. All mice were euthanized, and blood and small intestinal tissues were immediately collected. Briefly, a 5 cm segment of the ileum located 5 cm from the ileocecal junction, rinsed the intestinal contents out with saline, divided it into two parts, and each part was placed in a microtube and stored in a –80°C refrigerator.
Gastrointestinal motility test
Mice were fasted for 6 h with free access to water. One hour after treatment with the vehicle, extract, or positive control, each mouse received 0.3 mL of castor oil. One hour after castor oil administration, all mice received 0.2 mL of 5% activated charcoal suspension. The mice were then euthanized 30 min after the administration of charcoal meal. The abdomen of each mouse was opened, and the small intestine was immediately removed. The length of the small intestine, from the pylorus to the cecum, and the distance traveled by the charcoal marker were measured. The peristaltic index for each mouse was calculated as the percentage of the distance traveled by the charcoal meal relative to the total length of the small intestine.
RNA extraction and cDNA synthesis
Total RNA was extracted from small intestine tissues using the Axen Total RNA kit (Macrogen, Korea). One microgram of total RNA was reverse-transcribed into complementary DNA (cDNA) using the TOPscript™ cDNA Synthesis Kit (Enzynomics, Korea) with random primers according to the manufacturer’s instructions.
For reverse transcription, 1 μg of RNA was mixed with 1 μL of 50 μM random primers and 1 μL of 10 mM dNTP mix, and the volume was adjusted to 10 μL with RNase-free water. The mixture was heated at 65°C for 5 min to denature secondary RNA structures and then immediately cooled on ice. Subsequently, 4 μL of 5× TOPscript RT buffer, 0.5 μL of RNase inhibitor (40 U/μL), and 1 μL of TOPscript reverse transcriptase were added, and the final volume was adjusted to 20 μL with RNase-free water. The reverse transcription reaction was carried out at 42°C for 60 min, followed by enzyme inactivation at 85°C for 5 min. The synthesized cDNA was stored at –20°C until use.
Quantitative real-time polymerase chain reaction
Quantitative real-time polymerase chain reaction (qPCR) was performed using Axen™ qPCR Master Mix (2×, Probe) (Macrogen, Korea) according to the manufacturer’s instructions. Each 20 μL reaction mixture contained 10 μL of 2× qPCR Master Mix, 1 μL of template cDNA, 0.4 μM each of forward and reverse primers, and 0.2 μM of TaqMan probe, with the remaining volume adjusted using nuclease-free water. Primer information for qPCR is shown in Table 1.
Primers Used Real-Time Polymerase Chain Reaction
All reactions were performed in triplicate using the QuantStudio™ 5 Real-Time PCR System (Thermo Fisher Scientific, USA). Thermal cycling conditions were as follows: an initial denaturation at 95°C for 10 min, followed by 40 cycles of denaturation at 95°C for 15 sec and annealing/extension at 60°C for 1 min.
The threshold cycle (Ct) values obtained from qPCR were analyzed using the comparative Ct (2–ΔΔCt) method. The Ct value for the target gene in each sample was normalized to that of the housekeeping gene to calculate the ΔCt value. Subsequently, the ΔCt values of the experimental groups were compared with those of the control group to obtain the ΔΔCt values. Relative gene expression levels were calculated using the 2–ΔΔCt method and expressed as fold changes. All reactions were performed in triplicate to ensure reproducibility.
Western blotting
Proteins from small intestine tissues were extracted using radioimmunoprecipitation assay (RIPA) buffer, and whole-tissue lysates were prepared. Each protein sample was separated by sodium dodecyl sulfate–polyacrylamide gel electrophoresis and transferred to polyvinylidene fluoride membranes. After transfer, membranes were rinsed three times with Tris-buffered saline containing 0.1% Tween-20 (TBS-T), blocked for 1 h in BCodePro™ blocking solution (BIOCODE, Daejeon, Korea), and then incubated overnight with primary antibody, followed by incubation with horseradish peroxidase–conjugated secondary antibodies for 1 h at 37°C. Primary antibodies against phosphorylated and total extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), and p38 mitogen-activated protein kinase (p38 MAPK) (Cell Signaling Technology, Beverly, MA, USA) were diluted 1:1000. Finally, proteins were visualized using an enhanced chemiluminescence system (Amersham, NJ, USA). Images were captured using the Amersham™ ImageQuant 500 imaging system (Cytiva, MA, USA).
Enzyme-linked immunosorbent assay
Immunoglobulin G (IgG) and immunoglobulin A (IgA) concentrations in mouse serum were measured using ELISA kits (Invitrogen, Waltham, MA, USA) according to the manufacturer’s instructions.
Statistical analysis
Data are expressed as the mean ± standard error of the mean (SEM). Statistical analyses were performed using one-way analysis of variance followed by Tukey’s post hoc test using SPSS 28.0 software. A P value <.05 was considered statistically significant.
RESULTS
Effects of fermented soybean powder in castor oil-induced diarrhea in mice
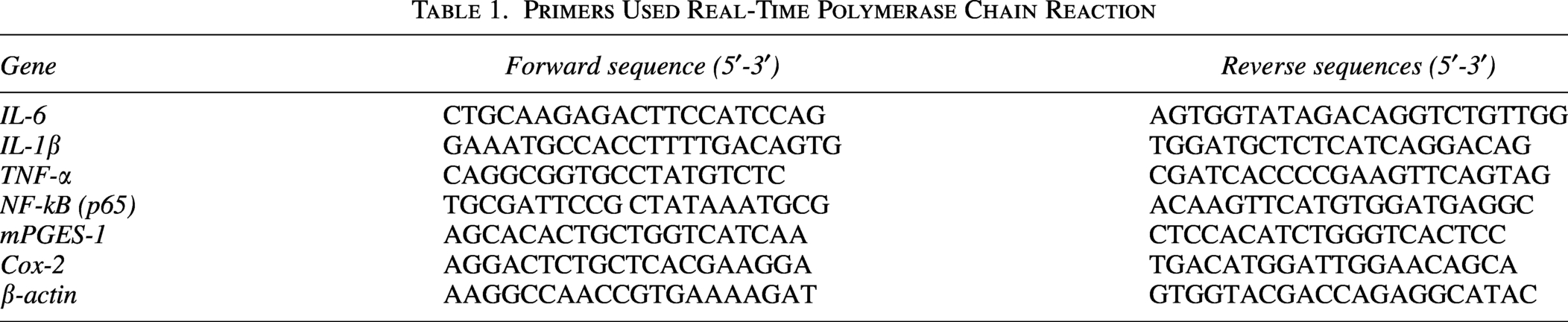
First, we examined the protective effects of FS on castor oil-induced diarrhea in vivo. In the DC group, diarrhea onset occurred 76.86 ± 5.62 min after castor oil administration. FS administration significantly delayed the onset of acute diarrhea (Fig. 1A). The onset times for diarrhea were 127.44 ± 16.15, 132.00 ± 9.77, and 133.13 ± 13.57 min in the 100, 200, and 300 mg/kg groups, respectively. Although FS treatment delayed the onset of diarrhea, no clear dose-dependent trend was observed. The effects were comparable to those observed with loperamide (124.33 ± 12.24 min) treatment. The frequency of diarrhea was determined as the ratio of watery stools to total stools for 4 h after castor oil administration. In the group administered castor oil, the frequency of diarrhea was 51.96 ± 6.15%, and the group administered FS showed a tendency for diarrhea to decrease. In particular, the group administered 200 mg/kg showed the most significant decrease in the frequency of diarrhea, at 32.93 ± 5.40%. In the group administered loperamide, the frequency of diarrhea was 40.76 ± 4.70%, which was lower than the group administered only castor oil, but it was not statistically significant (Fig. 1B).
Effects of fermented soybean powder on castor oil-induced diarrhea in mice.
In the castor oil-administered group, both fecal water content and feces weight increased. However, no significant reductions was observed in the FS-administered groups. This result was similar to that observed in the loperamide-administered group, which served as the positive control group (Fig. 1C, D).
Effects of fermented soybean powder on gastrointestinal transit in castor oil-induced diarrhea in mice
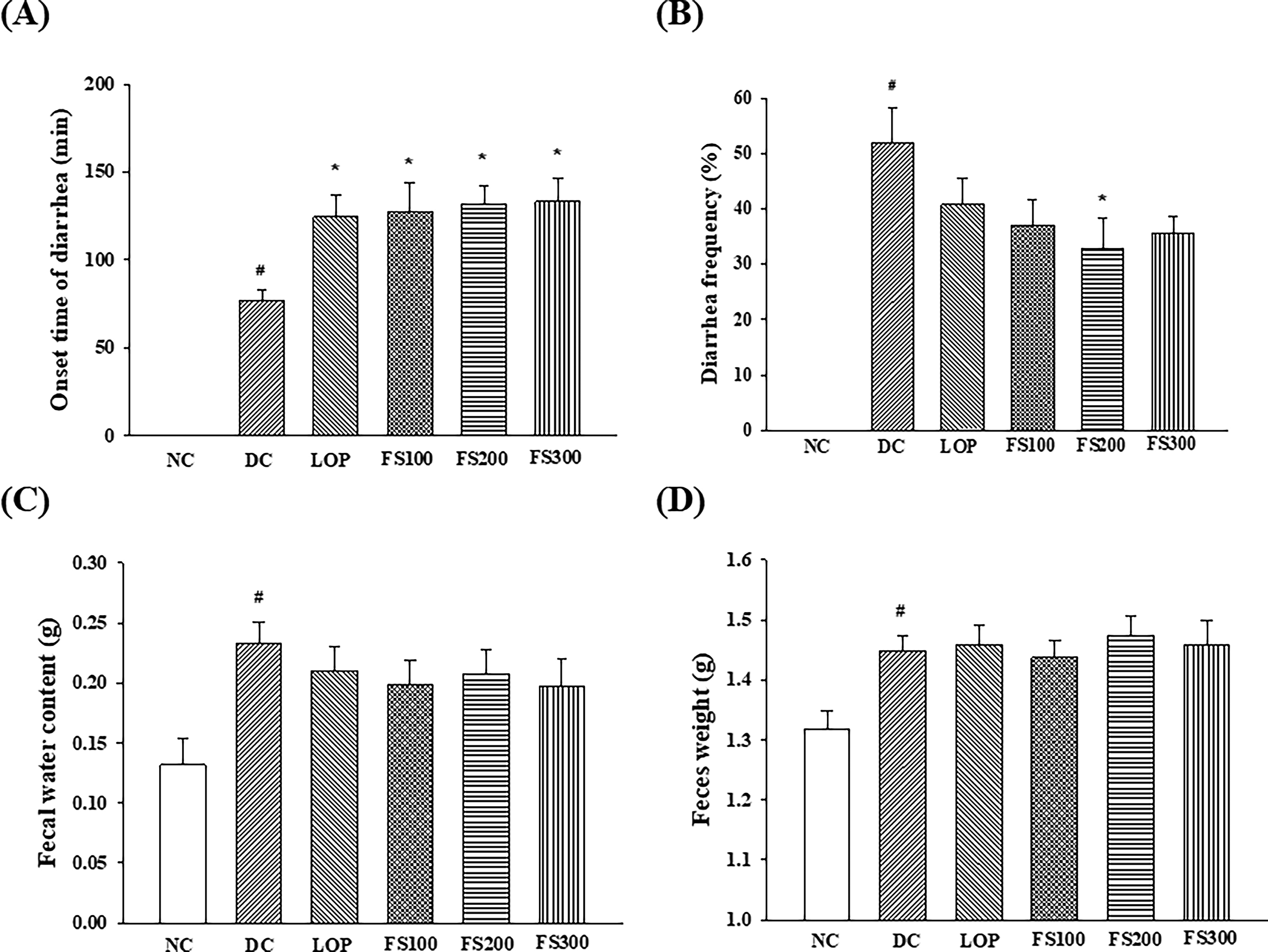
As shown in Figure 2, castor oil administration significantly increased the intestinal transit index in the DC group compared with the NC group, indicating enhanced GI motility. FS treatment significantly attenuated this increase. In particular, FS at doses of 200 mg/kg and 300 mg/kg markedly reduced the intestinal transit index compared with the DC group, restoring values close to those of the NC group. FS100 also showed a moderate reduction, although the effect was less pronounced. Notably, the effects of FS at higher doses were comparable to those of loperamide, suggesting that FS effectively normalizes castor oil-induced intestinal hypermotility.
Effects of fermented soybean powder on gastrointestinal transit in mice with castor oil-induced diarrhea. Values are expressed as the mean ± SEM. NC, normal control; DC, diarrheal control; LOP, loperamide 3 mg/kg; FS100, fermented soybean 100 mg/kg; FS200, fermented soybean 200 mg/kg; FS300, fermented soybean 300 mg/kg; SEM, standard error of the mean. #P < .05 compared to NC;*P < .05,** P < .01 compared to DC.
Effects of fermented soybean powder on pro-inflammatory cytokines level in castor oil-induced diarrhea in mice
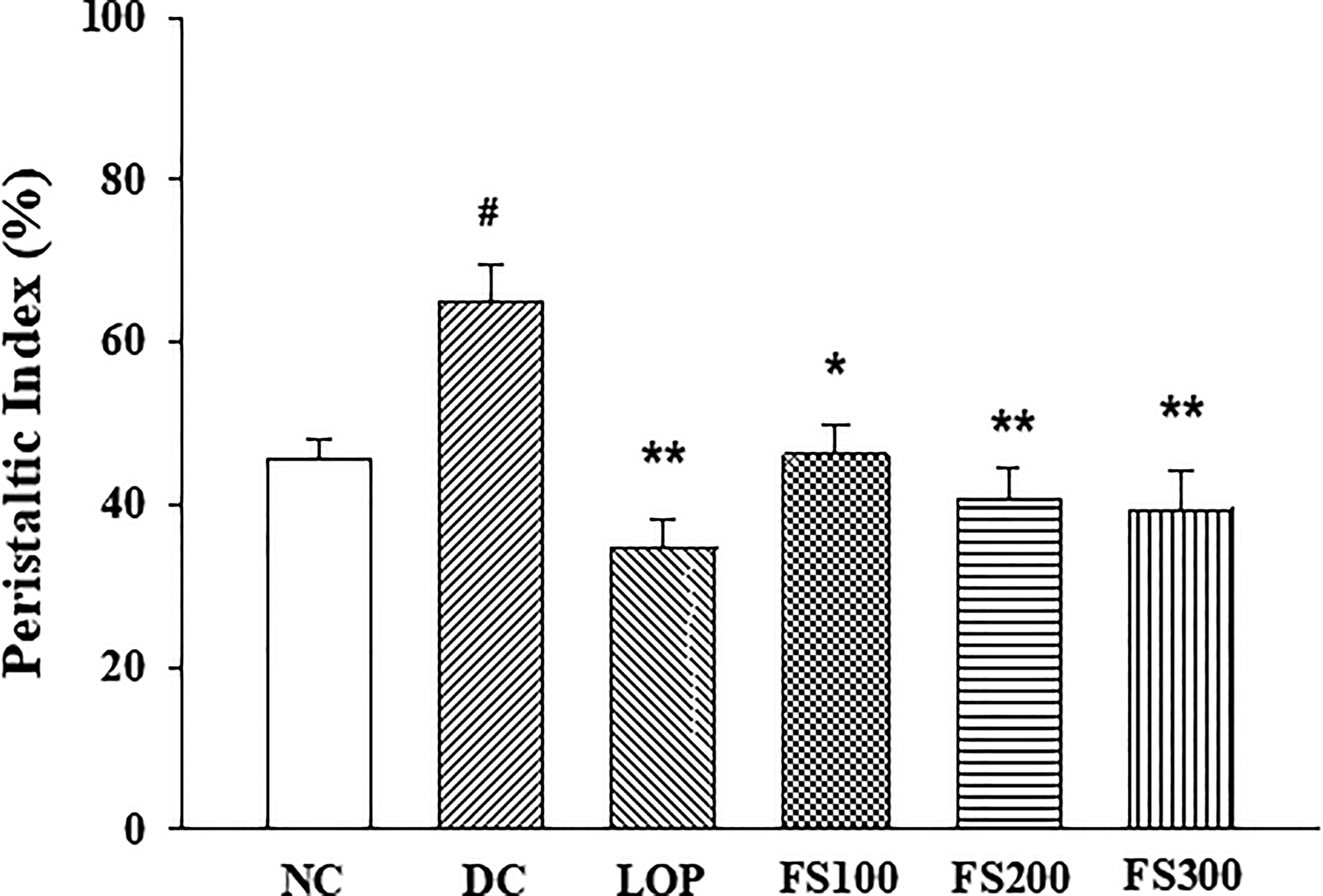
To confirm the intestinal immunoregulatory function of FS, changes in the expression of pro-inflammatory cytokines Il-6, Il-1β, and Tnf-α genes were investigated in small intestinal tissue. First, the Il-6 gene showed a significant increase in expression in the castor oil-induced diarrhea group. The FS-treated group showed a dose-dependent decrease in expression, but this was not statistically significant (Fig. 3A). The Il-1β gene also showed a significant increase in expression in the castor oil-induced diarrhea group, and this increase in gene expression was significantly suppressed in the group administered 200 mg/kg or more of FS (Fig. 3B). Tnf-α gene expression increased in the diarrhea group and tended to decrease with FS administration (Fig. 3C). Loperamide did not significantly affect the increased expression of inflammatory cytokines following castor oil administration.
Effects of fermented soybean powder on pro-inflammatory cytokines levels in mice with castor oil-induced diarrhea. The mRNA expression levels of
Effects of fermented soybean powder on inflammation-related gene expression in mice with castor oil-induced diarrhea
To investigate the anti-inflammatory effects of FS in a castor oil-induced diarrhea mouse model, the mRNA expression levels of Cox-2, mPGES-1, and NF-κB (p65) were evaluated in small intestinal tissues using real-time PCR (Fig. 4).
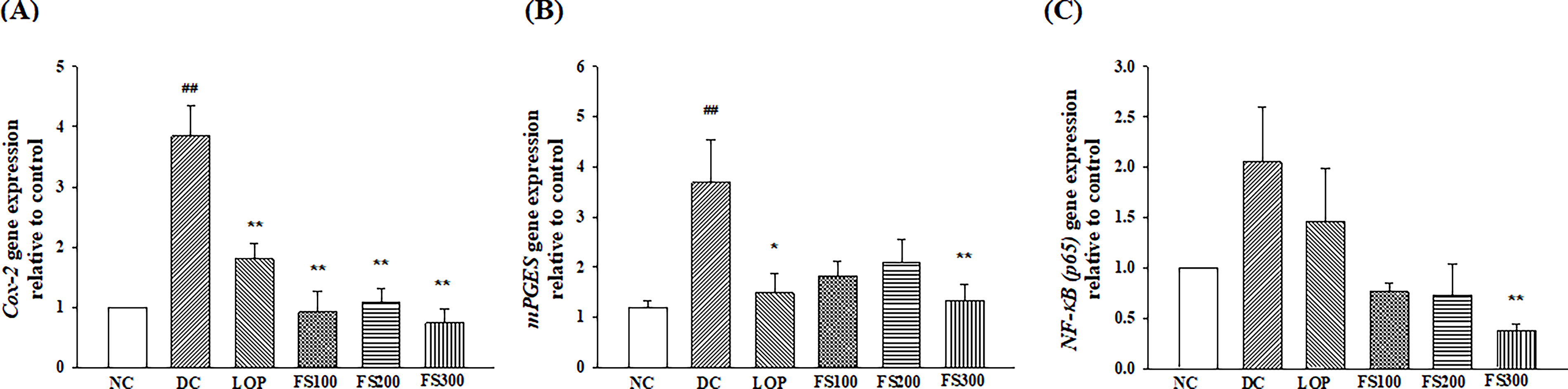
Effects of fermented soybean powder on inflammation-related gene expression in mice with castor oil-induced diarrhea. The mRNA expression levels of
As shown in Figure 4, the castor oil-induced diarrhea group exhibited a significant upregulation of Cox-2, mPGES-1, and NF-κB (p65) gene expression compared with the normal control group, indicating activation of intestinal inflammatory signaling by castor oil administration. Treatment with FS (FS100, FS200, and FS300 mg/kg) markedly attenuated these inflammatory responses. In particular, FS300 mg/kg treatment significantly downregulated NF-κB (p65) expression to below baseline levels (P < .01 vs. DC), suggesting strong suppression of transcriptional activation of inflammatory mediators. Similarly, Cox-2 and mPGES-1 expression levels were significantly reduced in all FS-treated groups (P < .05 or P < .01 vs. DC), comparable to the effect observed in the loperamide positive control group. These findings indicate that FS powder effectively inhibits the activation of the NF-κB/Cox-2/PGE2 inflammatory pathway, thereby alleviating intestinal inflammation induced by castor oil administration.
Effects of fermented soybean powder on serum immunoglobulin levels in castor oil-induced diarrheal mice
To determine whether FS affects intestinal immunity, plasma IgG and IgA concentrations were measured. In a model of acute diarrhea induced by castor oil administration, serum IgG and IgA levels were not affected. Similarly, short-term administration of FS or loperamide did not affect serum IgG and IgA levels (Fig. 5).
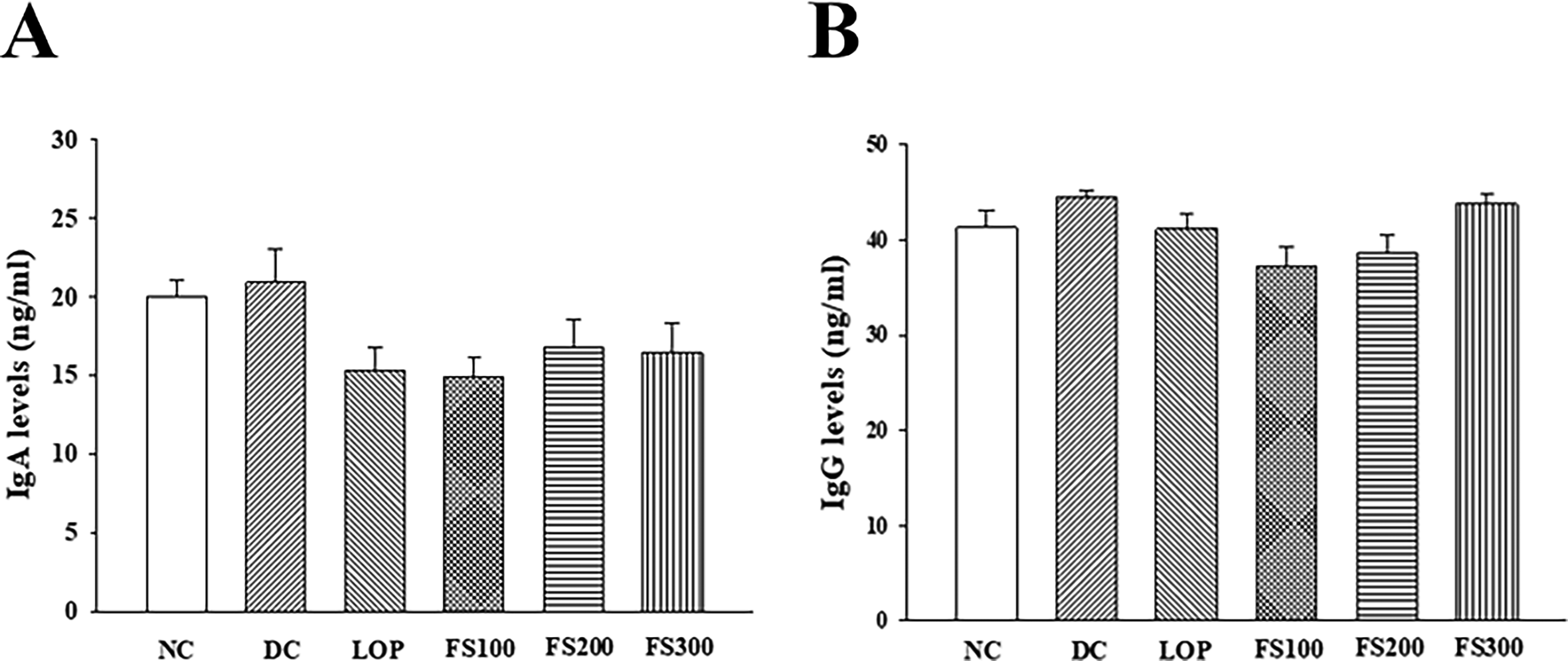
Effects of fermented soybean powder on serum immunoglobulin levels in mice with castor oil-induced diarrhea.
Effects of fermented soybean powder on phosphorylation of MAPKs in mice with castor oil-induced diarrhea
The phosphorylation levels of MAPKs, ERK, JNK, and p38 were examined to investigate the regulatory effects of FS on intestinal inflammatory signaling in mice with castor oil-induced diarrhea. As shown in Figure 6, the DC group exhibited a significant increase in the phosphorylation of ERK, JNK, and p38 compared with the NC group, indicating activation of MAPK-mediated inflammatory pathways. Treatment with FS markedly attenuated this activation. In particular, FS300 (300 mg/kg) significantly decreased ERK phosphorylation compared with the DC group (P < .05). Moreover, phosphorylation of JNK and p38 was significantly suppressed in all FS-treated groups (P < .01). The LOP group also showed decreased expression of p-ERK, p-JNK, and p-p38. These findings suggest that FS effectively mitigates castor oil-induced activation of MAPK signaling, thereby contributing to its antidiarrheal and anti-inflammatory effects in intestinal tissue.
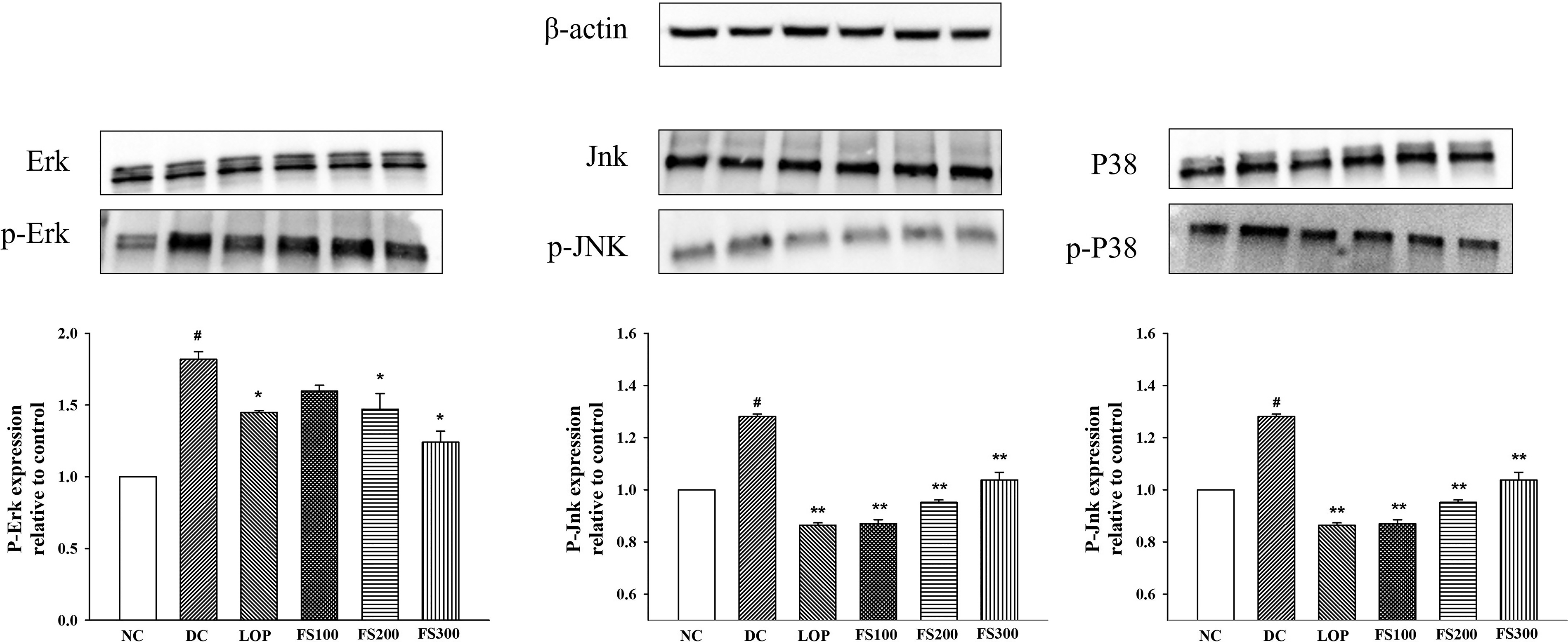
Effects of fermented soybean powder on phosphorylation of MAPKs in mice with castor oil-induced diarrhea. The phosphorylation of MAPKs, including ERK, JNK, and p38 in the small intestine was examined by western blot analysis. Expression levels are presented relative to the normal control group. Values are expressed as the mean ± SEM. NC, normal control; DC, diarrheal control; LOP, loperamide 3 mg/kg; FS100, fermented soybean 100 mg/kg; FS200, fermented soybean 200 mg/kg; FS300, fermented soybean 300 mg/kg; SEM, standard error of the mean. #P < .05 compared to NC; *P < .05, **P < .01 compared to DC.
DISCUSSION
The present study demonstrated that FS prepared using B. subtilis DKU_09 exerts potent antidiarrheal and anti-inflammatory effects in a castor oil-induced diarrhea mouse model. Oral administration of FS significantly delayed the onset of diarrhea, reduced the frequency of diarrhea, and normalized GI motility, with effects comparable to those of loperamide, a standard antidiarrhea drug. Moreover, FS markedly suppressed the intestinal expression of pro-inflammatory cytokines (Il-1β, Il-6, and Tnf-α) and inflammation-related genes (NF-κB, Cox-2, and mPGES-1), suggesting that its protective action are mediated through modulation of the NF-κB/Cox-2/PGE2 signaling pathway.
Castor oil-induced diarrhea is characterized by increased intestinal secretion and motility, mediated primarily by ricinoleic acid, the active metabolite of castor oil. A previous study showed that ricinoleic acid stimulates the release of prostaglandins, especially PGE2, which enhances intestinal fluid accumulation and peristalsis through activation of EP3 receptors on smooth muscle cells and enterocytes. 16 Although direct evidence for castor oil-induced diarrhea is limited, similar inflammatory responses involving NF-κB activation and COX-2/mPGES-1-mediated PGE2 synthesis have been reported in other studies, suggesting that these pathways may contribute to mucosal inflammation and secretory diarrhea.17,18 In this study, FS treatment downregulated these inflammation-associated markers, indicating that FS alleviates diarrhea not only by reducing intestinal secretion and motility but also by mitigating prostaglandin-driven inflammatory responses.
Inhibition of NF-κB activation is a key mechanism underlying many natural anti-inflammatory and antidiarrheal compounds. 19 Activation of NF-κB involves phosphorylation and nuclear translocation of the p65 subunit, which promotes the transcription of inflammatory mediators such as COX-2, iNOS, and pro-inflammatory cytokines. In the present study, FS treatment significantly reduced NF-κB (p65) mRNA expression, accompanied by the downregulation of multiple NF-κB-regulated inflammatory genes. These findings suggest that bioactive isoflavones generated during fermentation may contribute to suppression of NF-κB-mediated inflammatory responses. This observation is consistent with previous studies reporting that soybean-derived isoflavones, including genistein and daidzein, exert anti-inflammatory effects by inhibiting NF-κB and MAPK signaling pathways in intestinal epithelial cells and macrophages.20,21 In addition to NF-κB-related changes, FS also reduced the phosphorylation of MAPKs (ERK, JNK, and p38), which are key regulators of inflammatory gene expression and cytokine production during intestinal inflammation. These results support the notion that FS exerts broad anti-inflammatory effects by modulating multiple intracellular signaling pathways. Although NF-κB expression was evaluated at the mRNA level in the present study, the consistent downregulation of NF-κB-associated inflammatory genes suggests suppression of NF-κB-mediated signaling. However, further studies assessing NF-κB activation at the protein level, such as p65 phosphorylation or nuclear translocation, are required to confirm this mechanism.
Unlike traditional cheonggukjang, which is produced through spontaneous fermentation involving a complex and heterogeneous microbial community, the present study employed a single, well-defined strain, B. subtilis DKU_09, to ensure greater reproducibility and compositional consistency. The use of a defined starter culture enables controlled fermentation conditions and facilitates more precise evaluation of the biological effects of FS products.
Importantly, prior to the present study, we conducted a preliminary investigation using the same B. subtilis DKU_09 FS powder in a loperamide-induced constipation model. 15 In that study, FS powder significantly improved GI motility, fecal output, and fecal water content. In addition, HPLC analysis demonstrated that fermentation increased the levels of bioactive aglycone isoflavones while decreasing their glycoside forms, suggesting enhanced bioavailability of functional compounds.
Based on these findings, we hypothesized that B. subtilis DKU_09-FS powder may exert regulatory effects on intestinal physiology. Whereas our previous study demonstrated pro-motility effects under conditions of reduced intestinal activity, the present study extends these observations by demonstrating that the same FS preparation also alleviates diarrhea through suppression of intestinal inflammation and normalization of GI motility. These findings suggest that FS powder may function as a bidirectional regulator of intestinal homeostasis.
Fermentation by B. subtilis not only enhances the digestibility of soy proteins but also increases the concentration of bioavailable aglycone isoflavones and small peptides with antioxidant and anti-inflammatory potential. 22 Through HPLC analysis, we found that during the fermentation of soybeans with B. subtilis DKU_09, peaks related to glycoside forms (daidzin and genistin) gradually decreased, while peaks related to aglycone forms (daidzein and genistein) increased. 15 Isoflavones, including daidzein and genistein, have been widely reported to exert anti-inflammatory effects by suppressing pro-inflammatory cytokines such as IL-1β, IL-6, and TNF-α and by inhibiting NF-κB-mediated signaling pathways that regulate inflammatory mediators such as COX-2 and iNOS. 23 Therefore, the increased abundance of aglycone isoflavones observed after fermentation may contribute, at least in part, to the anti-diarrheal and anti-inflammatory effects observed in the present study. However, further studies are required to establish a direct causal relationship between compositional changes and biological activity.
Moreover, B. subtilis-based fermented foods have been reported to exhibit probiotic-like effects including stabilization of the gut microbiota and suppression of enteric pathogens. Although the present study did not directly assess microbiota composition, the observed reduction in inflammatory gene expression suggests that FS may help restore intestinal homeostasis disrupted by castor oil-induced inflammation.
IgA plays a critical role in mucosal immunity by protecting the intestinal epithelium against pathogens and maintaining intestinal barrier function, whereas IgG is primarily associated with systemic immune responses. Diarrhea is often accompanied by disruption of the intestinal barrier and alterations in mucosal defense mechanisms. Therefore, assessment of immunoglobulin levels can provide insight into whether the observed effects are associated with changes in mucosal or systemic immune responses. In the present study, serum IgA and IgG levels were not significantly altered. This observation suggests that the short-term intervention used in this acute diarrhea model primarily affected local mucosal inflammation rather than systemic immune responses. Chronic or infection-based diarrhea models may be more suitable for assessing the long-term immunomodulatory effects of FS products on mucosal immunity.
Taken together, these findings indicate that FS prepared using B. subtilis DKU_09 alleviates castor oil-induced diarrhea through multiple mechanisms: (1) delaying the onset of diarrhea and reducing fecal water loss, (2) normalizing GI motility, and (3) inhibiting NF-κB and MAPK signaling, leading to downregulation of Cox-2, mPGES-1, and pro-inflammatory cytokines. Collectively, these effects highlight the potential of FS as a natural functional food with both antidiarrheal and intestinal anti-inflammatory properties.
Future studies should explore the long-term efficacy of FS in chronic or infection-associated diarrhea and evaluate its impact on gut microbiota composition and metabolite production. Moreover, identification of the specific bioactive components such as isoflavone derivatives or Bacillus-derived peptides that are responsible for these protective actions will provide further insight into the molecular mechanisms and therapeutic potential of FS products in GI disorders.
Footnotes
ETHICS APPROVAL
The experimental protocol described as follows was approved by the Dankook University Institutional Animal Care and Use Committee (DKU-25-039).
AVAILABILITY OF DATA AND MATERIAL
The data that support the findings of this study are available from the corresponding author upon reasonable request.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This study was supported by the