
Abstract
Cognitive dysfunction has become a major health issue in the global aging society. For treating cognitive dysfunction or dementia, several small molecules or innovative drugs, including a monoclonal antibody against the amyloid β protein, have been prescribed. However, their adverse effects require the exploration of alternative treatments. Artemisia argyi (Asteraceae) has been used as traditional medicine for gastrointestinal disorders and inflammation. Recent reports suggest it has potential neuroprotective effects; however, evidence regarding its cognitive benefits and underlying mechanisms remains limited. We examined the memory-enhancing effects of an ethanolic extract of A. argyi (EAA) (10, 30, or 100 mg/kg, once per each behavioral test) in a mouse model of scopolamine-induced cognitive dysfunction and explored its mode of action on memory-related signaling pathways. Memory-related performance was assessed using the Y-maze, novel object recognition, Morris water maze, and passive avoidance tests. Next, Western blotting was used to evaluate changes in signaling molecules, such as CaMKII, ERK, CREB, and brain-derived neurotrophic factor (BDNF) in the hippocampus. EAA administration significantly improved cognitive performance across all tests. It restored the phosphorylation levels of CaMKII, ERK, and CREB, respectively, as well as the expression level of BDNF, after 12 h of treatment. Moreover, it enhanced long-term potentiation without altering basal synaptic transmission within the hippocampus. In summary, EAA facilitated cognitive enhancement during a cholinergic impaired state, attributable to its influence on synaptic plasticity and activation of the CaMKII–ERK–CREB–BDNF signaling cascade.
INTRODUCTION
Globally, cognitive decline has become a significant health issue, affecting those with neurodegenerative diseases like Alzheimer’s disease (AD), as well as the elderly population in general. 1 The World Health Organization estimates over 55 million people living with dementia worldwide, a figure which is expected to triple by 2050. 2 Current pharmacological treatments, such as cholinesterase inhibitors (e.g., donepezil), offer only symptomatic relief and are often associated with adverse effects, including nausea, diarrhea, bradycardia, and disturbed sleep. 3 Monoclonal antibody-based therapies targeting amyloid-beta (Aβ) plaques, such as lecanemab, have gained attention as an innovative AD treatments. 4 However, they pose several issues, including high costs, invasive administration routes, and adverse effects.5,6 As a result, the development of alternative therapeutic agents that are safer, more effective, and more suitable for long-term use is becoming pertinent.
Medicinal plants used in ethnomedicine have gained attention due to their established traditional use supported by emerging pharmacological evidence. 7 Several herbal medicines, including Salvia miltiorrhiza, Polygala tenuifolia, and Astragalus membranaceus, possess promising anti-cognitive dysfunction activity.8–10 Even if effective, their toxicity issues have not yet been clarified.11,12 Therefore, drug repositioning emerges as a promising method to develop a novel drug for cognitive dysfunction, since its safety has been previously established.
Artemisia argyi H. Lév. & Vaniot (Asteraceae) is a perennial herb used in traditional medicine for treating various ailments, including gastrointestinal discomfort and gynecological disorders. 13 It has also been widely employed in moxibustion therapy due to its body-warming and circulation-stimulating properties. 14 Based on such findings, an A. argyi extract is being clinically prescribed as an ethical drug for gastrointestinal discomfort in the Republic of Korea. 15 A. argyi contains a wide range of bioactives, including flavonoids, phenolic acids, essential oils, and coumarin derivatives.16,17 Among them are isochlorogenic acid (a phenolic) or scopoletin (a coumarin derivative), which enhance cognitive function.18,19 Other compounds have also been reported to demonstrate anti-inflammatory and cholinergic-modulating activities. 20 In addition, Artemisia princeps alleviated scopolamine-induced memory dysfunction in mice by suppressing acetylcholinesterase (AChE) and promoting hippocampal function. 21 Therefore, we hypothesize that A. argyi extracts can ameliorate cognitive impairment.
This study aimed to assess the pharmacological activities of an A. argyi extract on cognitive performance and explore the underlying mechanisms of action in the hippocampus. Scopolamine, a muscarinic acetylcholine receptor antagonist, is commonly utilized to induce transient cognitive dysfunction in animal models, 22 which can effectively mimic the aspects of cholinergic deficits observed in dementia, 23 and was employed in the present study.
MATERIALS AND METHODS
Animals
For this study, 6-week-old male ICR mice, 26–32 g, were purchased from Koatech Co., Ltd. (Gyeonggi-do, Korea). They were housed in groups of five per cage with ad libitum access to food and water. All animals were maintained in a university facility under controlled conditions: temperature at 23°C ± 1°C, humidity of 60% ± 10%, and a 12 h light/dark cycle (illumination from 07:30 to 19:30). The mice underwent acclimation for 1-week before each experiment. All procedures involving animals were conducted in accordance with the guidelines prescribed by Kyung Hee University for the care and use of laboratory animals and were approved by the Institutional Animal Care and Use Committee of the University (approval number: KHSASP-24-492).
Materials
Scopolamine hydrobromide and donepezil hydrochloride were obtained from Sigma-Aldrich Chemical Co., Ltd., (St. Louis, MO, USA). Rabbit polyclonal antibodies against phosphorylated Ca2+/calmodulin-dependent protein kinase II (pCaMKII) (#3361), phosphorylated extracellular signal-regulated kinase (pERK) (#9101S), phosphorylated cAMP response element-binding protein (pCREB) (#9198), and total CREB (#9197) were purchased from Cell Signaling Technology, Inc. (Danvers, MA, USA). Mouse monoclonal antibodies against CaMKII (sc-5306), ERK (sc-514302), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH; sc-166545) were procured from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA). A rabbit monoclonal antibody against BDNF (ab108319) was obtained from Abcam (Cambridge, UK). All other reagents were procured from reputed suppliers and were of the highest quality.
Preparation of A. argyi extract
Dried leaves of sorted A. argyi were purchased from the Gyeongdong Oriental Medicine Market (Seoul, Korea). They were extracted with 66.5% ethanol for 24 h at room temperature, and the extract was passed through 10 μm filters. The filtrate was vacuum-evaporated, and the resulting concentrate was vacuum-dried at 70°C to obtain a powdered ethanolic extract of A. argyi (EAA) with a yield of 10%. The EAA was standardized based on the content of its marker compound, isochlorogenic acid C. Reverse-phase high-performance liquid chromatography was performed using an Ultimate 3000 system equipped with a photodiode array detector (Thermo Scientific, Waltham, MA, USA). Separation was performed on an INNO C18-HC column, 5 μm, 4.6 × 250 mm (Young Jin Biochrom Co., Ltd., Gyeonggi-do, Korea) at 25°C with 1% aqueous acetic acid (solvent A) and acetonitrile containing 1% acetic acid (solvent B) as the mobile phases at a flow rate of 0.7 mL/min. The detection wavelength was set at 328 nm. The isochlorogenic acid C content was calculated using a calibration curve, and the mean extract content was 18.31 ± 0.18 mg/g (Supplementary Fig. S1).
Drug administration
EAA was dissolved in distilled water, whereas scopolamine and donepezil were solubilized in 0.9% saline. For each behavioral experiment, EAA (10, 30, or 100 mg/kg, p.o.), donepezil (5 mg/kg, p.o.), or vehicle solution was delivered to the mice 1 h before the Y-maze test, the training phase of the novel object recognition test (NORT), each training session of the Morris water maze test (MWMT), and the acquisition trial of passive avoidance test (PAT). Scopolamine (1 mg/kg, i.p.) or an equivalent volume of 0.9% saline was injected 30 min before each test, as mentioned with EAA.
Behavior tests
Cognitive function-related performances were assessed via the Y-maze test, NORT, MWMT, PAT. The Y-maze test and NORT were recorded and analyzed by an investigator who was blinded to the treatment conditions. The MWMT was analyzed using the EthoVision tracking system. More detailed behavioral procedures are presented in Supplementary Data.
Western blot analysis
For Western blot analysis, mice were sacrificed by cervical dislocation, and the brains were isolated. Hippocampi were rapidly dissected from the brains and homogenized in an ice-cold 20 mM Tris-HCl buffer (pH 7.4). Thereafter, routine procedures were conducted. The dilution of each antibody is provided in Supplementary Data.
Electrophysiology
For electrophysiological studies, mice were anesthetized with 2% isoflurane and euthanized by cervical dislocation. The brains were removed immediately and immersed in ice-cold artificial cerebrospinal fluid (aCSF; 4°C), after which the hippocampi were isolated. Transverse hippocampal slices—400 μm thick—were prepared with a McWill tissue chopper and incubated in continuously oxygenated aCSF (5% CO2, 95% O2) at 25°C ± 1°C for 1 h. The slices were then incubated for 2 h in aCSF containing a predesignated concentration of drug (EAA) before transfer to the recording chamber for electrophysiological measurements of the synaptic function associated with the Schaffer collateral pathway.
Field responses were evoked at 30 sec intervals, and the period when stable responses were obtained for at least 20 min was defined as the baseline. Paired-pulse ratio (PPR) and input/output (I/O) responses were subsequently recorded. To induce long-term potentiation (LTP), high-frequency stimulation (HFS), 100 pulses at 100 Hz, was delivered twice at an interval of 30 s, and changes in synaptic responses were monitored for 80 min.
Statistical analysis
All data are presented as the mean ± standard error of the mean. The Y-maze, PAT, MWMT, and Western blot results were evaluated using one-way analysis of variance (ANOVA) followed by Newman–Keuls post hoc comparisons. Additionally, two-way ANOVA followed by Bonferroni’s post hoc test was employed to analyze the object preference ratio in the NORT, escape latency in the MWMT, and BDNF levels. Differences were considered statistically significant at P < .05. All statistical analyses were conducted using Prism 9.0 software (GraphPad, La Jolla, CA, USA) and presented in Supplementary Table S1.
RESULTS
EAA improved short-term memory measured by the Y-maze test
The effects of EAA on short-term memory performance were evaluated through the Y-maze test. The spontaneous alternation scores differed significantly between groups (Fig. 1A). Scopolamine reduced spontaneous alternation (P < .05), whereas EAA (100 mg/kg) markedly improved it, as was observed with donepezil (P < .05). No significant inter-group variations in total arm entries were observed (Fig. 1B), suggesting that spontaneous locomotor behavior was not impacted by EAA treatment. 24
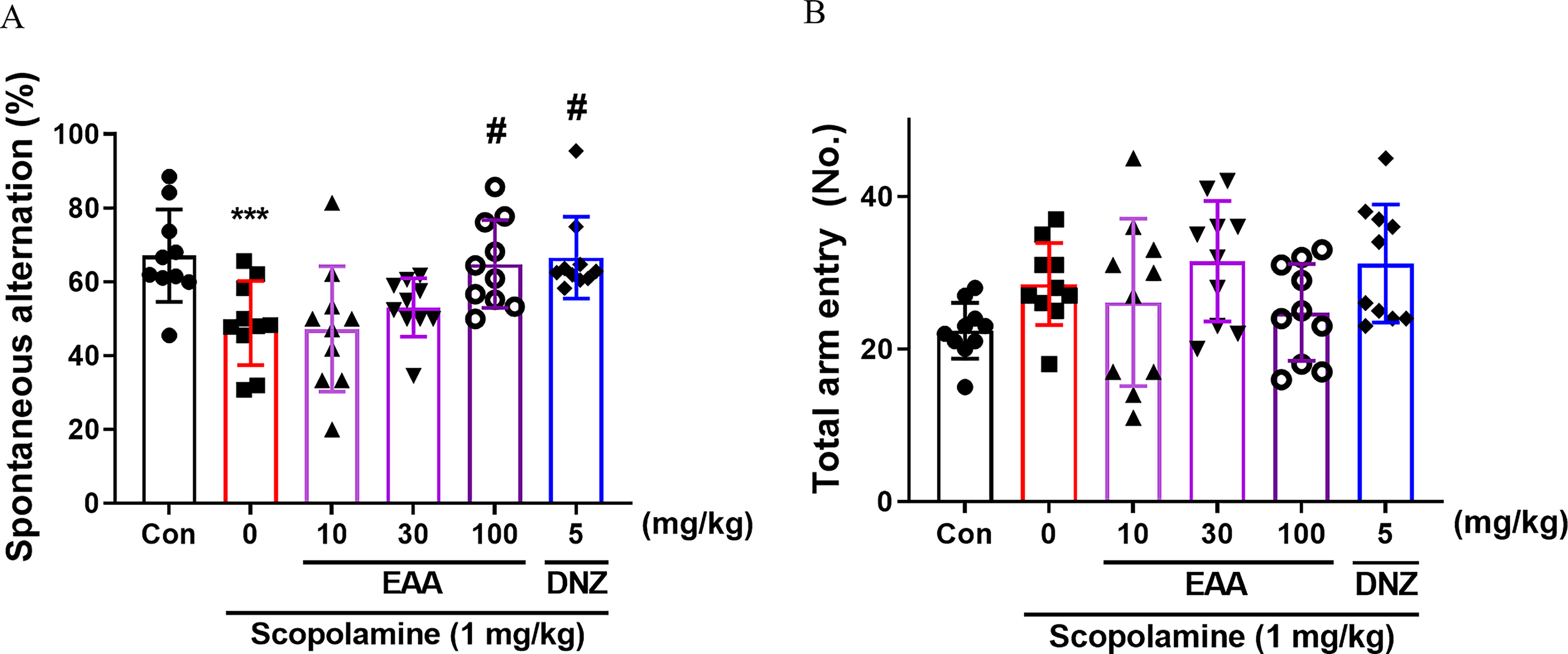
Effects of EAA on short-term memory measured by the Y-maze test. The spontaneous alternation
EAA ameliorated recognition memory as assessed by the NORT
The influence of EAA on object recognition memory in scopolamine-induced cognitively impaired mice was assessed using the NORT. Group-specific differences in object preference ratio were observed, as indicated by a two-way ANOVA (Fig. 2A). The EAA (30 and 100 mg/kg) and donepezil-treated groups demonstrated a substantially higher preference ratio to novel items compared to the scopolamine-treated group (P < .05). The inter-group discrimination and preference ratios exhibited similar trends in their graphical patterns (Fig. 2B). The EAA (30 and 100 mg/kg) and donepezil-treated groups exhibited a significant enhancement in the discrimination ratio compared to the scopolamine-treated group (P < .05).
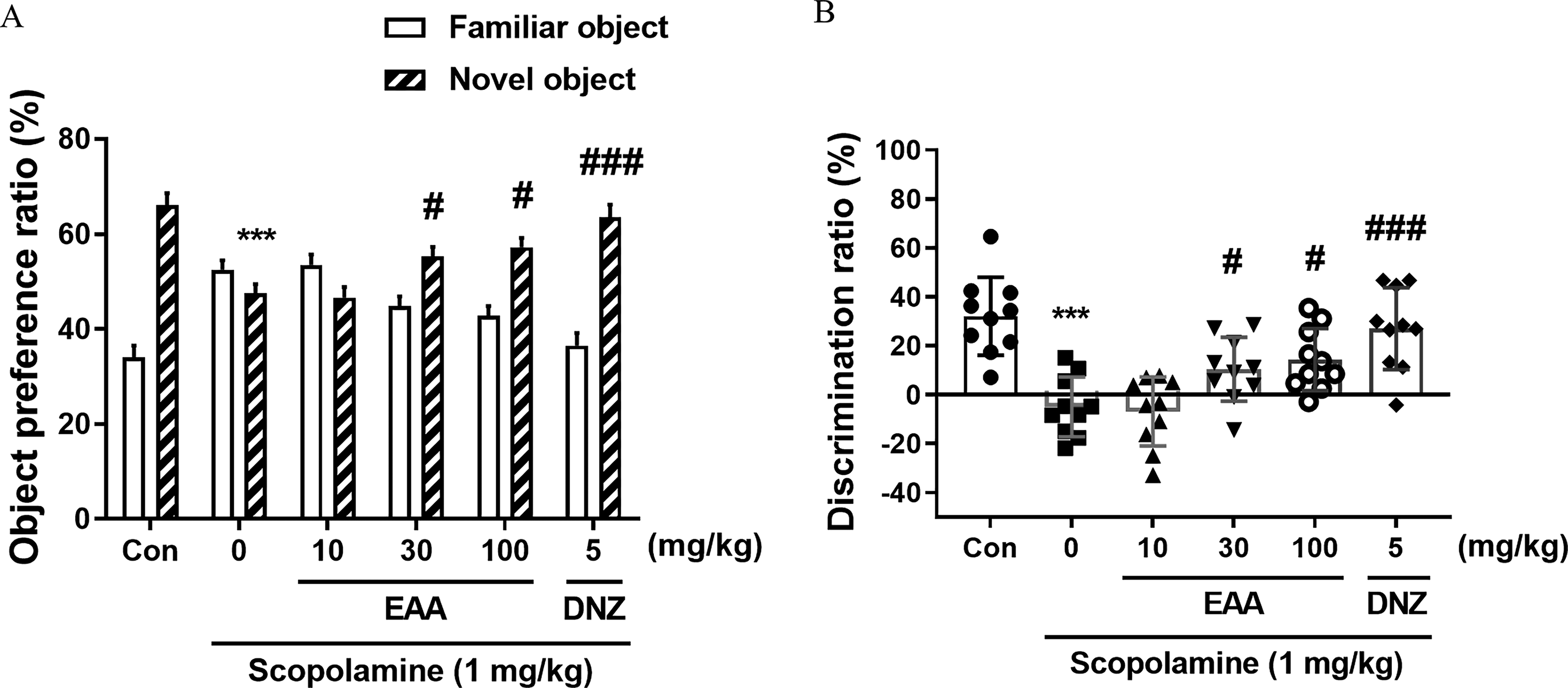
Effects of EAA on object recognition memory measured by the novel object recognition test (NORT). The object preference ratio
EAA ameliorated long-term spatial memory as measured by the MWMT
The MWMT was employed to investigate the effects of EAA on long-term spatial memory. 25 Significant inter-group variations in escape latency were observed during the training session, as indicated by two-way ANOVA (Fig. 3A). EAA administration (30 and 100 mg/kg) on training day 2 significantly suppressed the increase in escape latency induced by scopolamine during the training session (P < .05), as did donepezil. The time spent within the target quadrant significantly differed between groups (Fig. 3B). Time spent within the target quadrant declined with the scopolamine-treated group, which was markedly reversed by EAA (30 and 100 mg/kg) and donepezil (P < .05). The number of platform crossings and swimming speed did not vary significantly among groups (Fig. 3C, D). A heatmap was plotted, which showed that the EAA-treated group of mice exhibited an improved search behavior and stayed in the target quadrant longer (Fig. 3E).
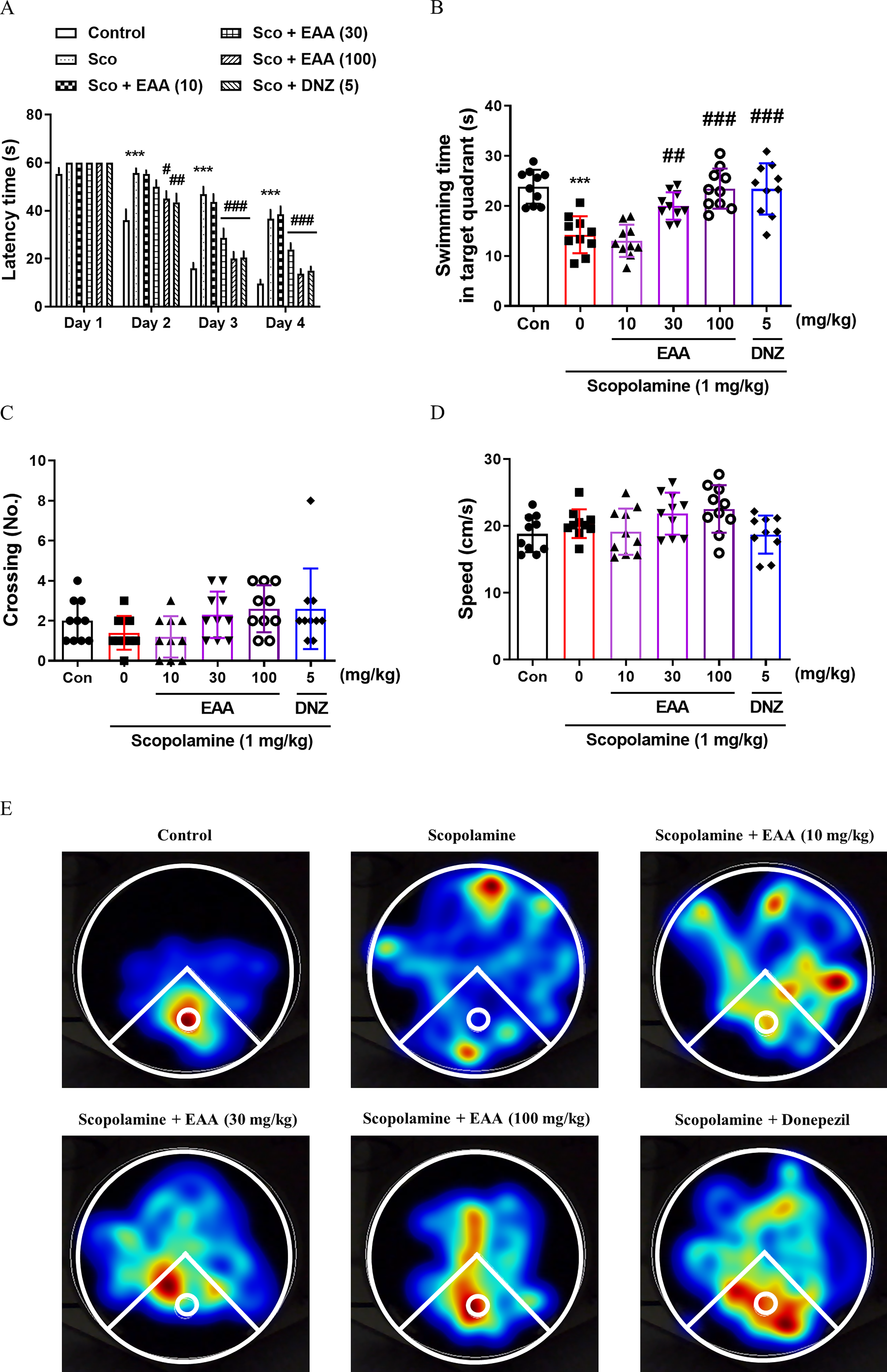
Effects of EAA on spatial long-term memory measured by the Morris water maze test (MWMT). The time required to locate the platform during the training sessions in MWMT was presented
EAA restored long-term memory as estimated by the PAT
The PAT was used to assess whether EAA affected long-term memory in cholinergic blockade-based cognitively impaired mice. In the acquisition trial, step-through latency did not differ significantly across groups (Fig. 4). In the retention trial, significant inter-group differences were observed (Fig. 4). The group treated with scopolamine had a considerably lower latency than the control group (P < .05), whereas the groups that received EAA (30 and 100 mg/kg) and donepezil showed a significantly enhanced latency compared to the scopolamine-treated group (P < .05).
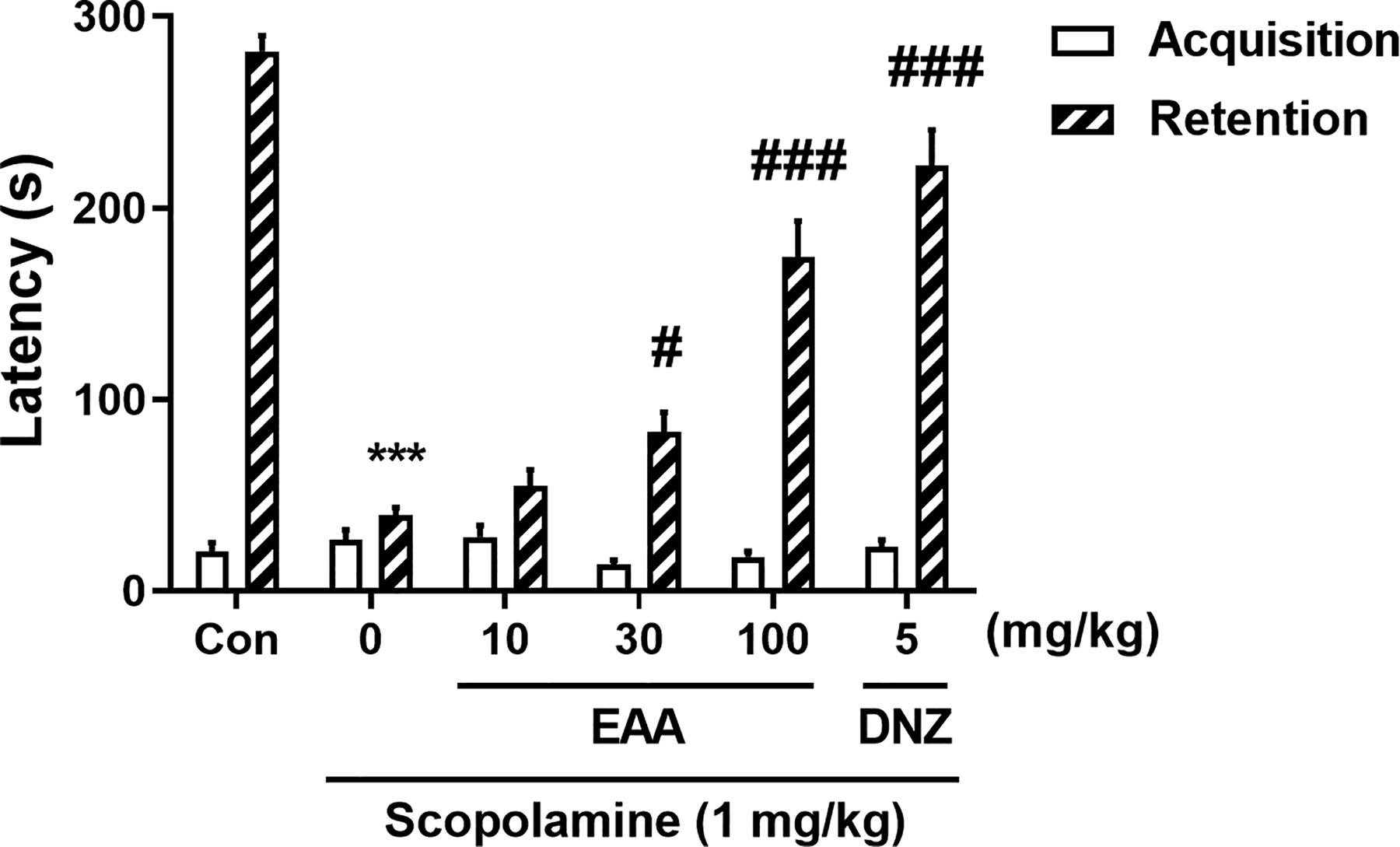
Effects of EAA on long-term memory measured by the passive avoidance test (PAT). The step-through latencies of the acquisition trial and the retention trial in the PAT were presented. In the acquisition trial, mice entered the dark chamber and received a foot shock. After 24 h, memory retention was assessed by measuring step-through latency during the retention trial. Data were presented as mean ± SEM (n = 10/group).***P < .001 versus the vehicle-administered control group; #P < .05 and ###P < .001 versus the scopolamine-administered group. Con, control; DNZ, donepezil; EAA, ethanolic extract of A. argyi; SEM, standard error of the mean.
Effects of EAA on the hippocampal CaMKII–ERK–CREB signaling cascade
The phosphorylation levels of hippocampal CaMKII, ERK, and CREB were evaluated following EAA administration. Significant inter-group differences were observed in the phosphorylation status (Fig. 5A–D). The phosphorylation levels of CaMKII, ERK, and CREB were significantly diminished in the scopolamine-treated group compared to the control group, which were markedly reversed by EAA (P < .05).
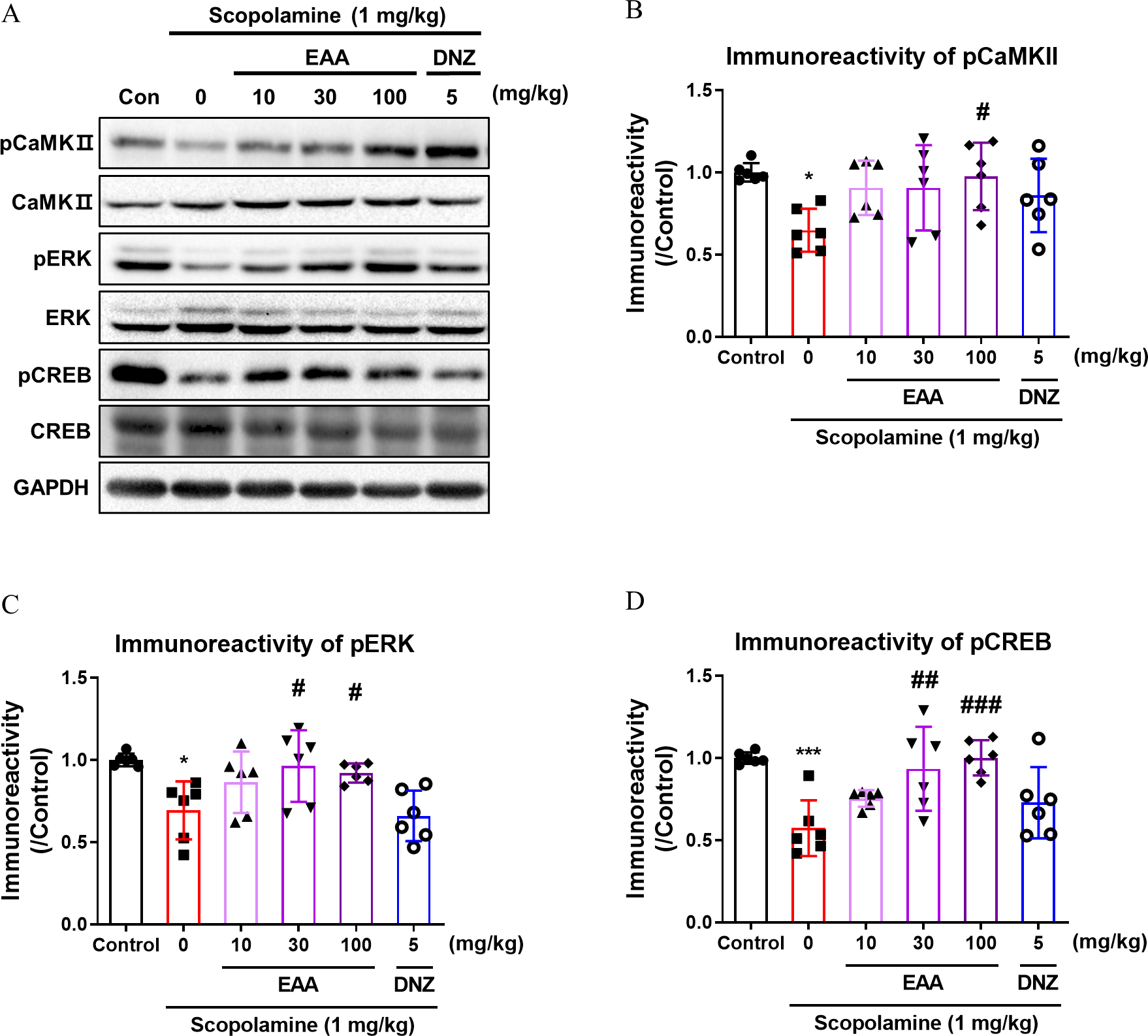
Effects of EAA on the phosphorylation levels of CaMKII, ERK, and CREB in the hippocampus of mice. Representative immunoblots
Effects of EAA on BDNF expression in the hippocampus
Acquired memory can be consolidated through elevated BDNF expression ∼12 h after training. 26 Therefore, we investigated temporal changes in BDNF expression in the hippocampus following EAA administration and found significant inter-group differences (Fig. 6). At 12 h post treatment, EAA (100 mg/kg) significantly increased BDNF expression in the hippocampus compared with the control group (P < .001).
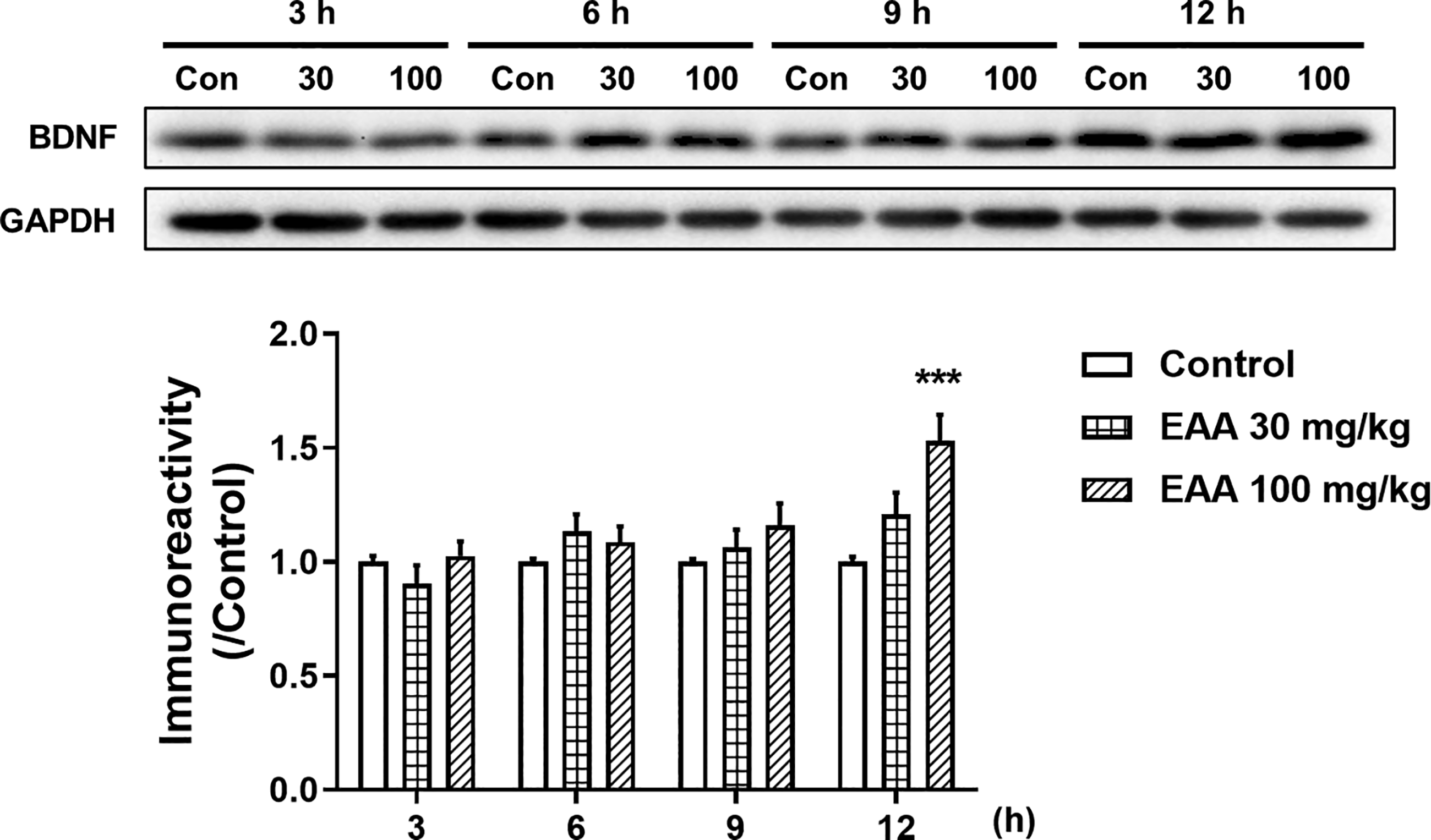
The temporal changes of the BDNF expression levels in the hippocampus after administration with EAA. Representative immunoblots and densitometric analysis of BDNF in the hippocampus of mice. Band intensity was normalized to GAPDH and expressed relative to the control group (taken as 1.0). Data were presented as mean ± SEM (n = 6/group) (*P < .05, versus the vehicle-administered control group). Con, control; SEM, standard error of the mean.
Effects of EAA on basal synaptic transmission and LTP
The impacts of EAA on synaptic transmission and LTP were analyzed. The slope of the postsynaptic response (field excitatory postsynaptic potential [fEPSP]) presented in Figure 7A was measured to compare synaptic responsiveness. No significant variations were observed between the EAA-treated and control groups, indicating that EAA did not affect basal synaptic responses. Similarly, presynaptic response levels were comparable across all groups (Fig. 7B), suggesting that EAA does not alter presynaptic signaling. In addition, the PPR values were similar to those of the controls under all test conditions (Fig. 7C), confirming that EAA did not influence presynaptic neurotransmitter release. These findings suggest that EAA did not affect excitatory synaptic basal activity within the hippocampal Schaffer collateral pathway, which plays a key role in hippocampal function.
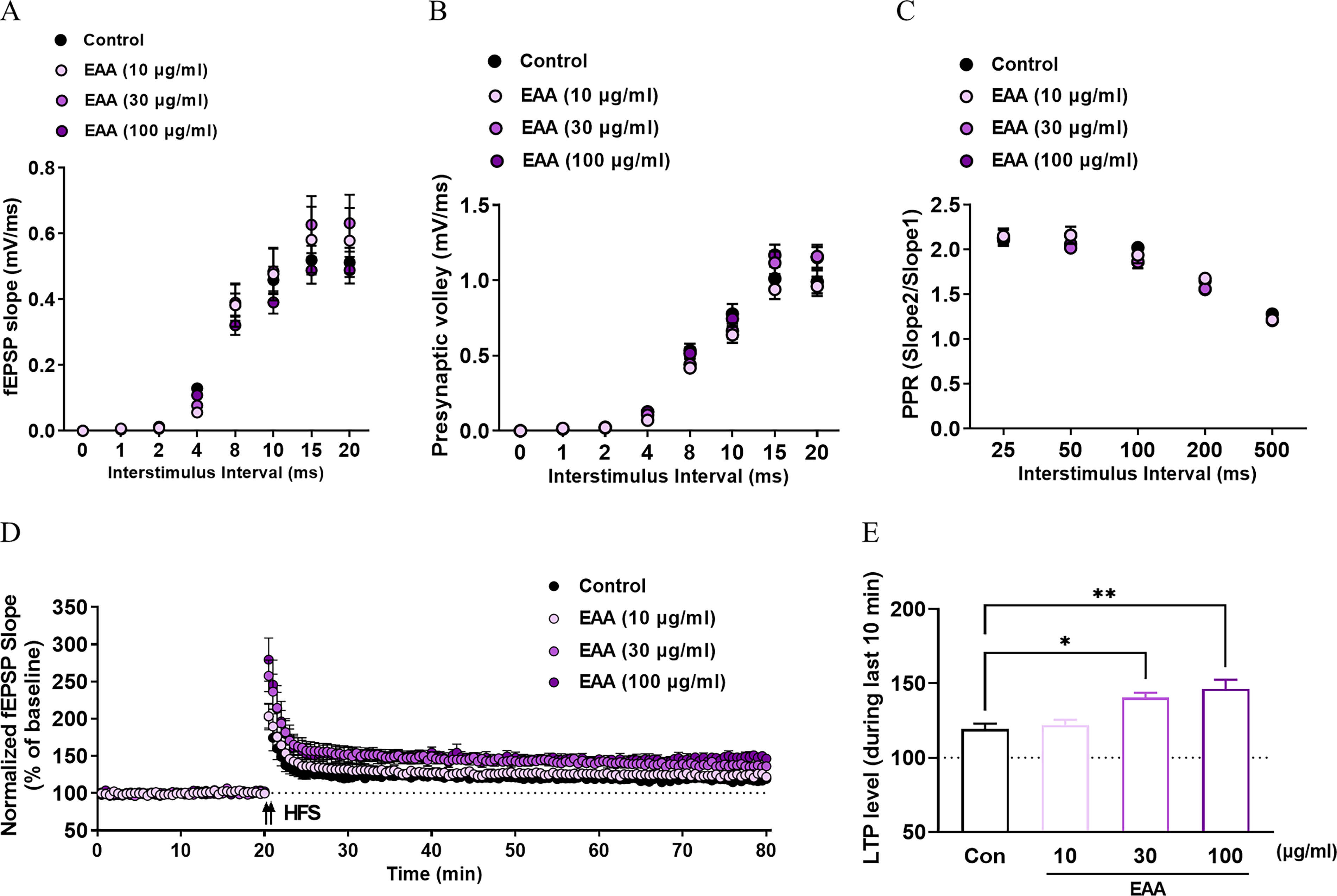
Effects of EAA on basal synaptic transmission and long-term potentiation (LTP) in hippocampal slices. Field excitatory postsynaptic potential (fEPSP) slope as a function of interstimulus interval (ISI) in control and EAA-treated slices
To evaluate LTP, the temporal enhancement of synaptic responses following HFS was measured. The EAA-treated groups exhibited dose-dependent increases in synaptic potentiation (Fig. 7D). Furthermore, quantitative statistical analysis revealed that EAA at 30 and 100 μg/mL significantly enhanced LTP compared with the LTP of the control group (P < .05 or P < .01), indicating that EAA facilitated synaptic plasticity (Fig. 7E).
DISCUSSION
The present study demonstrated that EAA administration markedly improved cognitive performance in mice, as evidenced by behavioral assessments, such as the Y-maze test, NORT, MWMT, and PAT. In addition, it also enhanced the CaMKII–ERK–CREB signaling pathway activity, hippocampal BDNF expression, and synaptic plasticity.
A. argyi has been traditionally used for treating gastrointestinal discomfort and gynecological disorders. 13 Additionally, A. princeps reversed scopolamine-induced memory impairment by inhibiting AChE. 21 As A. princeps and A. argyi belong to the same genus, as well as demonstrating overlapping phytochemical profiles, 13 it is reasonable to speculate that A. argyi would also induce cognitive-enhancing effects. During the behavioral experiments, we observed that EAA administration in a scopolamine-induced memory-impaired mouse model improved short-term, long-term, recognition, and spatial memory, as ascertained via the Y-maze test, PAT, NORT, and MWMT, respectively. Next, we explored whether this influence of EAA may be derived from its inhibition of AChE, as was observed in A. princeps. 21 Fermented A. argyi by Monascus purpureus also reversed cognitive deficits through AChE inhibition. 27 However, contrary to expectations, EAA did not exhibit any remarkable effects on AChE activity (data not shown). M. purpureus-fermented products exhibited neuroprotective or cognitive ameliorating activities. 28 Although the compound(s) that mainly play a role in cognitive function were not explored, likely novel compound(s) produced during fermentation by M. purpureus might protect against oxidative stress or cognitive impairments. Therefore, it can be speculated that these compound(s) may suppress AChE, which explains the varying effects of EAA or fermented A. argyi on AChE activity. Further research may clarify and confirm this hypothesis. Instead, we investigated whether the effects of EAA are mediated by signaling pathway(s) within the hippocampus.
Western blotting revealed that EAA normalized the levels of pCaMKII, pERK, and pCREB. ERK is associated with the expression of BDNF via CREB activation and plays a vital role in memory consolidation and synaptic plasticity.29,30 CREB is a transcription factor that regulates a variety of central nervous system-associated functions, including long-term memory, neurogenesis, and neuronal survival; it is also essential for maintaining neuronal health and function. 31 Thus, the present results suggest that mitogen-activated protein kinase (MAPK) or CaMKII signaling pathways related to memory consolidation could be activated by EAA. 32 In addition, we observed a significant elevation in BDNF expression 12 h after EAA administration. BDNF is a crucial factor in the pathogenesis of neurodegenerative diseases such as AD, as its expression is reduced in AD patients. 33 BDNF is considered an essential molecule in modulating memory-impairment-related disorders as it increases synaptic transmission efficiency and contributes to memory consolidation.34,35 Therefore, our data suggest that restoring ERK activity by EAA treatment may enhance synaptic plasticity, possibly through the CREB-dependent upregulation of BDNF expression. In addition, activated CaMKII can phosphorylate CREB and plays a critical role in the regulation of LTP.36,37 Therefore, based on these results, therefore, electrophysiological experiments were performed to analyze the influence of EAA on synaptic transmission and LTP. The findings demonstrated that EAA did not affect basal synaptic transmission in the hippocampus; however, it significantly enhanced HFS-induced synaptic potentiation in a dose-dependent manner, thereby promoting synaptic plasticity. These results suggest that the cognitive-enhancing impacts of EAA are closely associated with its ability to facilitate synaptic plasticity. Therefore, it may be suggested that EAA improves cognitive function by activating the MAPK and CaMKII signaling pathways or enhancing synaptic transmission.
The present study has several limitations. The CaMKII–ERK–CREB–BDNF signaling cascade suggests an increase in synaptic plasticity, 38 and this study observed that EAA facilitated such signaling pathways. However, we could not identify the molecule(s) that play a major role in EAA-induced enhancement in cognitive function, as we did not precisely investigate the involvement of each molecule. Second, EAA was standardized by isochlorogenic acid C, as a marker compound. However, considering its concentration in EAA, other cognition-activity-related compound(s) may be better candidates. In addition, as mentioned, we found that EAA did not affect AChE activity and, therefore, the present results could not support the use of EAA as a cholinergic targeting drug. Instead, the enhanced MAPK and CaMKII signaling pathways, as well as synaptic plasticity, are associated with the glutamatergic neurotransmitter system. 39 Therefore, further studies are needed to specify the target(s) or the bioactive principal compound(s) of EAA.
CONCLUSIONS
In conclusion, standardized EAA exerts memory-enhancing effects in mice, as evidenced by several behavioral tests, mediated by activation of CaMKII–ERK–CREB signaling and upregulation of BDNF expression. While the precise compounds remain to be identified, these results suggest its potential as a functional food or therapeutic candidate for memory impairment. Collectively, the present study suggests that A. argyi can be a potential medicinal herb for treating cognitive dysfunction observed in impaired cholinergic neurotransmitter systems.
AUTHORS’ CONTRIBUTIONS
S.H.B. and K.P.: Investigation, formal analysis, and writing—original draft. S.M.: Investigation and data curation. T.H.L., S.H.L., H.H.J., and E.H.H.: Investigation and validation. J.E.H. and S.Y.J.: Formal analysis and data curation. D.H.K.: Supervision and writing—review and editing. J.H.R.: Conceptualization, supervision, and writing—review and editing.
Footnotes
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
FUNDING INFORMATION
This research was partially funded by the BK21 FOUR program of the Graduate School at Kyung Hee University (
Supplemental Material
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.