
Abstract
Objectives
In a previous study by Torres-Henderson et al, the probiotic Enterococcus lactis strain SF68 was clinically shown to lessen amoxicillin-clavulanate-associated diarrhea. The cause or mechanism as to why this clinical benefit occurred, however, was not apparent in that study. Since the completion of that work, a new feline dysbiosis index (DI) has been developed and new information concerning the microbiome and bile acid metabolism has been published. The objective of this study was to assess changes in the feline DI and fecal bile acid metabolism to explain how this probiotic lessened amoxicillin-clavulanate-associated diarrhea.
Methods
A prospective, randomized, placebo-controlled, double-blinded study was conducted with 27 healthy, young, adult, purpose-bred cats that were used for a previous study. The cats were randomized into the probiotic group (13 cats) or placebo group (14 cats). Each cat received amoxicillin-clavulanate for 7 days at a standard dose and either the probiotic or placebo for 14 days. Fecal samples from all cats were obtained at baseline, day 7 and day 14. All samples were frozen at −80°C until assessed. Targeted quantitative PCR assays were then used to determine the fecal feline DI, and unconjugated fecal bile acid concentrations were measured at each time point.
Results
An antibiotic-induced shift in fecal microbiota and unconjugated bile acids was observed, with the probiotic group showing normalization of the feline DI and increased secondary bile acids on day 14 compared with the placebo group (P = 0.0322).
Conclusions and relevance
The probiotic E lactis strain SF68 lessened amoxicillin-clavulanate-associated clinical signs of disease at least in part from the positive effects on the selected microbiota and bile acid metabolism. These findings support supplementing this probiotic to cats who have a clinical need for amoxicillin-clavulanate treatment.
Keywords
Introduction
Bacterial infections occur in some cats, necessitating the use of antibiotics. Amoxicillin-clavulanate is indicated for the treatment of cats with susceptible bacterial infections, often those in the Gram-positive class. However, antibiotic use in cats can increase antibiotic-resistant microbes, induce signs of gastrointestinal disease, and promote both acute and long-term dysbiosis in cats.1 –4 Thus, empirical use of antibiotics should be avoided in cats for syndromes not associated with bacterial infections, such as feline upper respiratory tract infections with viral etiology or feline idiopathic cystitis. Adhering to the American Association of Feline Practitioners and the American Animal Hospital Association antimicrobial stewardship guidelines 5 and other guidelines for the appropriate use of antibiotics can help decrease the misuse of antibiotics in dogs and cats.6,7
Probiotics are live microorganisms that provide health benefits when administered in adequate amounts. 8 Prebiotics are substrates that are selectively utilized by host microorganisms and confer a health benefit. When probiotics and prebiotics are supplemented together, the combination is called a synbiotic. Bacterial genera such as Lactobacillus, Bifidobacterium and Enterococcus are commonly used as probiotics in both humans and pets. Multiple studies have shown the clinical benefits of these probiotics.9 –12 In humans, one of the most studied uses for probiotics is to attempt to lessen antibiotic-associated side effects, with varying conclusions being made from the results of meta-analyses.12,13 The available data concerning the use of commercially available probiotic-containing products to lessen antibiotic-associated gastrointestinal side effects are minimal in cats, with only clindamycin and amoxicillin-clavulanate being studied to date.2 –4
A multi-strain synbiotic was used in clindamycin studies3,4 and a single strain probiotic was used in an amoxicillin-clavulanate study. 2 In each of the experiments, clinical benefits were noted in the groups of cats in which the probiotic or synbiotic were fed before the administration of the antibiotic when compared with cats in the control group. However, the mechanisms explaining the clinical responses were not clear using untargeted 16S PCR gene sequencing. Since the publication of those studies, a new dysbiosis index using targeted quantitative PCR (qPCR) assays that amplify the DNA of select, specific bacteria to attempt to discriminate healthy cats from those with dysbiosis was developed. 14 In addition, further information concerning the role of bacteria-associated unconjugated bile acids in cats with chronic enteropathy has now been published. 15 Therefore, the objective of this study was to assess microbial associated bile acid (BA) metabolism and characterize the fecal microbiota with a qPCR-based dysbiosis index using samples from a previous study. 2
Materials and methods
Animals and study design
This Institutional Animal Care and Use Committee approved (protocol 170.010) prospective, randomized, placebo-controlled, double-blinded study included 27 healthy, young, adult, purpose-bred research cats. 2 Each cat was considered healthy based on a normal physical examination, the presence of a normal fecal score throughout a 10-day equilibration period and negative results for parasite eggs, cysts and oocysts on microscopic examination of feces after sugar centrifugation. In addition, the cats were all negative for Giardia species and Cryptosporidium species by commercially available direct fluorescent antibody assay (Merifluor Giardia/Cryptosporidium; Meridian Diagnostics). The commercially available product containing Enterococcus lactis strain SF68 (FortiFlora; Purina Pet Care, previously classified as Enterococcus faecium strain SF68) 16 and a placebo (the identical product but no bacteria) was provided by the sponsor in marked capsules. The clinical scoring team was masked to the capsule contents and applied a previously published standardized scoring system to each fecal sample that was passed by each cat (fecal score: 0 = no stool; 1 = very hard and dry; 2 = firm but not hard; 3 = normal, little or no segmentation, moist; 4 = very moist, log shaped; 5 = very moist, piles; 6 = texture but no defined shape; or 7 = watery puddle). 2 The probiotic is susceptible to amoxicillin-clavulanate (Clavamox; Zoetis Animal Health) and so was fed with food 2 h before the antibiotic was administered to avoid potential inactivation. The cats were acclimated to individual cages for a 10-day equilibration period to exclude those prone to stress-associated diarrhea. After the study began, the cats were fed 1/4 cans of canned food mixed with either the probiotic or placebo at 6 am daily. Two hours later, all cats were administered amoxicillin-clavulanate at a dosage of 62.5 mg/cat, which was then repeated at 6 pm for 7 days. The probiotic and placebo administration continued for an additional 7 days after stopping the antibiotic. Fecal samples were collected at baseline, day 7 and day 14 and had been stored at −80°C until assayed in the study described herein.
Fecal microbiota analysis
An analytically validated targeted qPCR panel was applied to evaluate the fecal microbiota by measuring the abundances of Bacteroides and Bifidobacterium species, Peptoacetobacter (previously Clostridium) hiranonis, Escherichia coli, and Faecalibacterium, Streptococcus and Turicibacter species. The dysbiosis index (DI) was calculated based on the results of the targeted qPCR assays as previously described. 14 Briefly, DNA was extracted using the DNeasy PowerSoil Pro Kits (QIAGEN), normalized, and mixed with pre-mixed primer and SsoFast EvaGreen supermix (Bio-Rad Laboratories). The Bio-Rad C1000 Touch Thermal Cycler was set with initial denaturation at 98°C for 2 mins, followed by 40 cycles with denaturation at 98°C for 3 s and annealing for 3 s. Besides the targeted bacteria in the DI panel, Clostridium perfringens and Enterococcus species were also measured. These qPCR assays were analytically validated, and the conditions are described in previous studies.14,15
Fecal concentrations of sterols and unconjugated bile acids
An analytically validated targeted gas chromatography with mass spectrometry method was used to measure the fecal concentrations of five unconjugated bile acids, as previously described. 16 The targeted compounds included primary bile acids (cholic acid [CA] and chenodeoxycholic acid [CDCA]) and secondary bile acids (lithocholic acid [LCA], deoxycholic acid [DCA] and ursodeoxycholic acid [UDCA]).
Statistical analysis
The abundances of targeted bacterial taxa, the DI and fecal concentrations of targeted metabolites at three time points for all cats were analyzed using mixed-effects models to assess the effect of treatment (placebo vs probiotic) and time (baseline, antibiotics, off-antibiotics), followed by Tukey’s multiple comparison tests. In addition, the correlation between the abundance of P hiranonis and the fecal concentration of secondary BAs was assessed using Spearman’s rank correlation test. The effect of the probiotics on the DI and the abundance of P hiranonis on day 14 were evaluated using Fisher’s exact test. For this analysis, the DI was categorized into normal (DI <0) and abnormal (DI >0), while the abundance of P hiranonis was categorized as within or below the reference interval (RI) (4.5). 15 Statistical analyses were performed using GraphPad Prism 9.0 (GraphPad Software).
Results
Targeted fecal microbiota analysis
After the cats were administered amoxicillin-clavulanate for 7 days, the DI significantly increased (adjusted P <0.0001) in both groups (Figure 1). By day 14, the DI in the probiotic group was similar to baseline (adjusted P = 0.17), whereas the DI in the placebo group remained significantly increased (adjusted P = 0.0281). By day 14 (7 days off antibiotics), 7/8 (87.5%) cats in the probiotic group had a normal DI, while 6/8 (75%) cats in the placebo group remained dysbiotic (P = 0.041, Fisher’s exact test).

Dysbiosis index (DI) on days 0 (baseline), 7 (7 days of amoxicillin-clavulanate administration [Abx]) and 14 (7 days after the last dose of antibiotic) in cats fed a probiotic (red dots, n = 8) or a placebo (black dots, n = 8). The gray area represents the reference interval (DI <0), while the dashed lines (DI 0–1) represent the equivocal range. A DI >1 indicates significant dysbiosis
The fecal abundance of Enterococcus, which is the genus to which the probiotic strain belongs, significantly increased on day 7 and then decreased by day 14 in the placebo group (Figure 2). In contrast, a significant increase in Enterococcus species abundance was observed in the probiotic group on day 14 but not on day 7. In addition, the fecal abundance of C perfringens significantly decreased in both groups after antibiotic treatment and remained unchanged on day 14.

(a,b) Fecal abundance of Enterococcus species and Clostridium perfringens on days 0 (baseline), 7 (7 days of amoxicillin-clavulanate administration [Abx]) and 14 (7 days after the last dose of antibiotic) in cats fed a probiotic (red dots, n = 8) or placebo (black dots, n = 8)
The fecal abundance of P hiranonis significantly decreased by day 7 in both groups (Figure 3). By day 14, the P hiranonis abundance in the probiotic group – but not the placebo group – was significantly increased compared with day 7 results. Moreover, in the placebo group, the abundance remained significantly lower on day 14 compared with day 0. Specifically, by day 14, P hiranonis abundance had recovered into the RI in 6/8 cats in the probiotic group, while 5/8 cats in the placebo group still had reduced P hiranonis abundance.
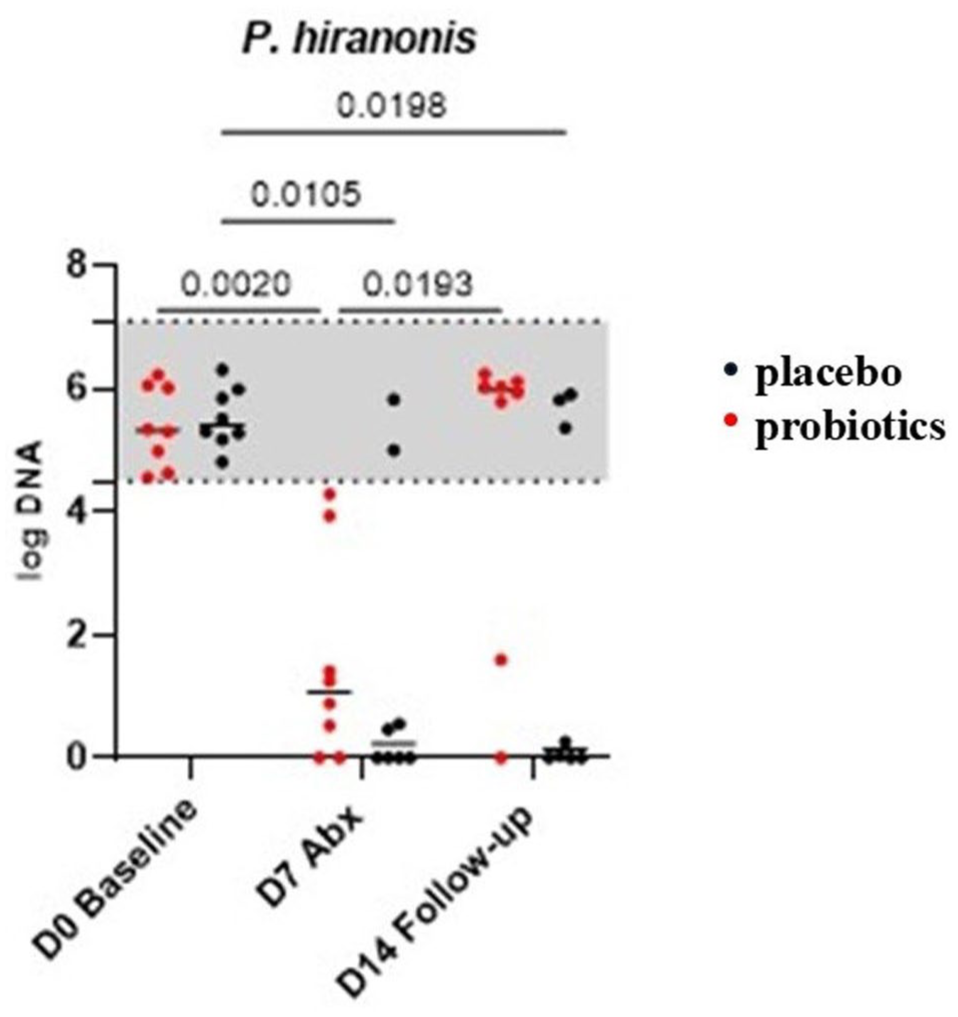
Fecal abundance of Peptacetobacter hiranonis on days 0 (baseline), 7 (7 days of amoxicillin-clavulanate administration [Abx]) and 14 (7 days after the last dose of antibiotic) in cats fed a probiotic (red dots, n = 8) or placebo (black dots, n = 8). The gray area represents the reference interval
In both groups, the fecal abundances of Bifidobacterium, Faecalibacterium and Turicibacter species significantly decreased after 7 days of antibiotic administration compared with baseline levels (Figure 4). However, Streptococcus species abundance significantly decreased on day 7 only in the placebo group. Conversely, E coli abundance significantly increased on day 7 in both groups. By day 14, the abundances of Bifidobacterium, Faecalibacterium, Turicibacter and Streptococcus species in both groups did not differ from baseline levels.

Fecal abundance of (a) Bifidobacterium, (b) Faecalibacterium, (c) Turicibacter, (d) Streptococcus, (e) Escherichia coli and (f) Bacteroides species targeted bacteria on days 0 (baseline), 7 (7 days of amoxicillin-clavulanate administration [Abx]) and 14 (7 days after the last dose of antibiotic) in cats fed a probiotic (red dots, n = 8) or placebo (black dots, n = 8). The gray area represents the reference interval
Fecal concentrations of unconjugated bile acids
Fecal concentrations of primary bile acids (CA and CDCA) remained consistent across the three time points (Table 1). However, fecal concentrations of individual secondary bile acids (LCA, DCA and UDCA) significantly decreased after 7 days of antibiotic treatment (P <0.01) in both groups. Notably, there was a statistically significant interaction between time (antibiotic) and treatment (probiotic) for UDCA (P = 0.02). Total measured secondary BAs also significantly decreased on day 7 compared with baseline and day 14 (Figure 5). In the probiotic group, fecal concentrations of secondary BAs on day 14 were significantly higher than on day 7, a trend not observed in the placebo group. Furthermore, the fecal abundance of P hiranonis was significantly negatively correlated with the percentage of primary BAs, with a Spearman’s ρ of −0.63 (−0.78 to −0.42; P <0.0001, n = 48).
Fecal concentrations of unconjugated bile acids (µg/mg) in cats (n = 16) on days 0, 7 and 14

Data are median (range). Amoxicillin-clavulanate was administered to all cats until day 7, and the probiotic or placebo was fed until day 14. Comparisons between days were analyzed using two-way ANOVA tests, followed by Tukey’s multiple comparisons tests. Median values within the same row with different superscript letters differ significantly (P <0.01). The P values in bold have values of < 0.05.
CA = cholic acid; CDCA = chenodeoxycholic acid; CON = placebo group; DCA = deoxycholic acid; LCA = lithocholic acid; primary BA = sum of CA and CDCA; PRO = probiotic group; secondary BA = sum of LCA, DCA and UDCA; total BA = sum of CA, CDCA, LCA, DCA and UDCA; Tx = treatment; UDCA = ursodeoxycholic acid

Fecal concentration of secondary bile acids (sum of unconjugated lithocholic acid, deoxycholic acid and ursodeoxycholic acid) on days 0 (baseline), 7 (7 days of amoxicillin-clavulanate administration [Abx]) and 14 (7 days after the last dose of antibiotic) in cats fed a probiotic (red dots, n = 8) or placebo (black dots, n = 8). The gray area represents the reference interval. Horizontal lines represent the medians
Discussion
This study applied targeted approaches to investigate the effect of a probiotic (E lactis strain SF68) on minimizing the disruption of intestinal microbiota and microbial-associated bile acid metabolism in cats undergoing oral amoxicillin-clavulanate treatment. An antibiotic-induced shift in fecal microbiota and unconjugated bile acids was observed, with the probiotic group showing a faster recovery based on the normalization of the feline DI and increased secondary BAs on day 14 compared with the placebo group. Enterococcus, the genus to which the probiotic belongs, increased on day 14 in the probiotic group, indicating that the probiotic strain could traverse the gastrointestinal tract and resist amoxicillin-clavulanate, even though susceptibility testing showed E lactis strain SF68 to be susceptible to this antibiotic.
In the study described here, an increased DI and disruption in BA metabolism were observed in all healthy cats administered amoxicillin-clavulanate. Similar DI results have been noted in other studies with amoxicillin-clavulanate1,17 and other antibiotics, including cefovecin, 18 clindamycin,3,4 doxycycline 1 and pradofloxacin. 19 Although the long-term impact of antibiotic exposure in cats has not been widely studied, early antibiotic exposure in humans has been linked to increased risks of developing gastrointestinal diseases and asthma.19 –21 Our results provide further evidence that antibiotics should only be administered to cats with true bacterial infections or those with syndromes usually associated with antibiotic responsive bacterial, protozoal or Rickettsial pathogens.
The disrupted BA metabolism observed in this study is likely due to antibiotic-induced dysbiosis. BA metabolism is heavily influenced by microbial activity. Host-derived conjugated primary BAs are secreted into the small intestine, where a broad array of bacteria, including Bifidobacterium, Bacteroides, Clostridium, Enterococcus and Lactobacillus species, possess bile salt hydrolases that deconjugate taurine from these primary BAs. Subsequently, a more limited group of bacteria with the bile acid-inducible (bai) operon convert these deconjugated bile acids into secondary BAs through 7 (alpha) α-dehydroxylation. In humans, the main bacteria with 7α-dehydroxylating activity include Clostridium scindens, Clostridium hylemonae, Clostridium sordellii and P hiranonis.22 –24 In cats, P hiranonis has been identified as a primary BA converter.16,25 Consistent with previous studies, this study found a significant decrease in the fecal abundance of P hiranonis in cats administered amoxicillin-clavulanate, which corresponded with a reduction in secondary BAs. The significant correlation between the fecal abundance of P hiranonis and the percentage of primary BAs further supports the role of P hiranonis as an important BA converter in cats.
In the previous study that was performed to generate the samples assayed in the experiments described here, 2 a number of clinical findings suggested that feeding E lactis strain SF68 before administering amoxicillin-clavulanate was beneficial. Notably, fecal scores of 7 were only detected in the placebo group; when total diarrhea scores were compared between groups for days 1–11, the scores for cats fed E lactis strain SF68 were statistically lower (P = 0.0058). In the prior publication, we were unable to provide an explanation for the apparent clinical effects with the methods used. 2 In the study described here, the E lactis strain SF68-supplemented group showed a faster recovery of the DI, particularly the abundance of P hiranonis, in concert with increased secondary BAs on day 14 compared with the placebo group. These changes are likely in part the explanation for the clinical effects noted. In addition, in the probiotic group, fecal concentrations of primary BAs decreased at the time of follow-up, but this change was not observed in the placebo group. Overall, the findings suggest that feeding the probiotic accelerated the recovery of the microbial community responsible for BA transformation and resolution of clinical signs induced by amoxicillin-clavulanate.
Compared with previously published sequencing results, 2 the effects of the probiotic were more easily discernible with quantitative data in this study. Although sequencing data indicated an overall microbial shift in principal component analysis plots, it did not provide specific details on the extent and direction of the changes from baseline to day 7 and day 14. By using RIs for the DI and each targeted bacterium, we could more clearly observe the trends in changes. For instance, the abundances of Bifidobacterium and Enterococcus species showed similar trends when comparing relative abundance (percentage) from sequencing data with quantitative abundance (log DNA) from qPCR assays. However, 16S rRNA gene sequencing lacks the resolution needed to accurately identify changes at the genus or species level. E coli, a member of the Enterobacteriaceae family, was not identified in the previous sequencing study. In contrast, the current study detected E coli in all samples, noting an abnormal abundance above the upper reference limit on day 7, which returned to RIs by day 14. This nuance could not be captured by sequencing data, where most cats exhibited a nearly negligible relative abundance of Enterobacteriaceae species at baseline and follow-up, with an increase of up to 90% on day 7. Because of the nature of relative abundance, interpreting the numbers is difficult.
The changes in fecal microbiota (increased DI, increased E coli, decreased Faecalibacterium species, decreased P hiranonis, decreased Bifidobacterium species) and BA profiles induced by antibiotics were similar to those observed in cats with CE.14,16,25 Although cats fed the probiotic showed a faster recovery, some did not return to baseline by day 14, suggesting that antibiotic-induced dysbiosis may persist in a subset of cats, even with probiotic co-administration. Similar to a previous study, oral administration of amoxicillin-clavulanate and pradofloxacin in kittens induced diarrhea and increased susceptibility to enteropathogens, even when a potential probiotic strain (Enterococcus hirae) was given before and during the inoculation of the enteropathogenic E coli. 26
The association between diarrhea and the isolation of C perfringens and Clostridioides difficile has not been definitively established in cats. 27 In this study, C perfringens was detected in all cats, all of which were clinically healthy and exhibited no gastrointestinal signs. This finding is similar to previous research in dogs, which found that C perfringens was present in both clinically healthy dogs and those with gastrointestinal signs. 28 After antibiotic treatment, the abundance of C perfringens decreased and remained stable on day 14 in all cats in the present study. The clinical significance of this change remains unclear, as the presence of C perfringens does not necessarily correlate with disease, and its role in feline gastrointestinal health requires further investigation.
This study has some limitations, including the lack of long-term follow-up, making it unknown when and if the microbiota returned to normal in the placebo group. However, a previous study has shown that a single subcutaneous antibiotic administration in cats led to at least 4-month dysbiosis. 26 In addition, it is unclear if the E lactis strain SF68 engrafted long term. However, based on other studies, it is likely that this strain requires continuous administration. 29 In this study, clinical signs were induced by administration of the antibiotic; therefore, the findings of this study cannot be directly applied to client-owned cats with chronic enteropathies. However, shelter cats fed E lactis strain SF68 were less likely to experience diarrhea lasting more than 2 days compared with the placebo group, suggesting that this probiotic has potential benefits in cats with gastrointestinal signs. 10
Future studies should address these gaps by incorporating long-term follow-up periods to assess the lasting impact of both antibiotics and probiotics on the feline microbiota. It would also be valuable to investigate whether probiotic supplementation can prevent the onset of dysbiosis or, with prolonged use, restore the microbiota to a healthy state after disruption. Moreover, expanding research to include cats with gastrointestinal diseases, such as chronic enteropathies, could provide critical insights into the therapeutic potential of probiotics in managing such conditions and offer a more comprehensive understanding of how these interventions may affect different populations within the species.
Conclusions
The probiotic E lactis strain SF68 was shown in this model to lessen amoxicillin-clavulanate-associated clinical signs of disease, likely through beneficial effects on selected microbiota and BA metabolism, and could be considered for use with client-owned cats receiving this antibiotic. The targeted qPCR assays and DI described here may be useful for monitoring treatment effects.
Footnotes
Author note
These data were presented in part at the 2023 American College of Veterinary Internal Medicine Forum in Philadelphia, Pennsylvania. The abstract program was published in the Journal of Veterinary Internal Medicine 2023; 38: 530–729.
Conflict of interest
Drs Lappin and Suchodolski are both consultants for Purina PetCare and Dr Suchodolski is the Purina Petcare Endowed Chair for microbiome research. However, this study was performed independently from the company in our two laboratories. Drs Suchodolski and Sung are employees of the Gastrointestinal Laboratory, which offers gastrointestinal function and microbiome testing on a fee-for-service basis.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Ethical approval
This work did not involve the use of animals and therefore ethical approval was not specifically required for publication in JFMS.
Informed consent
This work did not involve the use of animals (including cadavers) and therefore informed consent was not required. No animals or people are identifiable within this publication, and therefore additional informed consent for publication was not required.