
Abstract
Autism spectrum disorder is a neurodevelopmental disorder characterized by a wide range of behavioral alterations, including impaired social interaction and repetitive behaviors. Numerous pharmacological interventions have been developed for autism spectrum disorder, often proving ineffective and accompanied by a multitude of side effects. The gut microbial alterations observed in individuals with autism spectrum disorder, including elevated levels of Bacteroidetes, Firmicutes, and Proteobacteria, as well as reduced levels of Bifidobacterium, provide a basis for further investigation. Recent preclinical studies have shown favorable outcomes with probiotic therapy, including improvements in oxidative stress, anti-inflammatory effects, regulation of neurotransmitters, and restoration of microbial balance. The aim of this review is to explore the potential of probiotics for the management and treatment of autism spectrum disorder, by investigating insights from recent studies in animals.
Lay abstract
Autism spectrum disorder is a neurodevelopmental disorder characterized by a wide range of behavioral alterations, including impaired social interaction and repetitive behaviors. Numerous pharmacological interventions have been developed for autism spectrum disorder, often proving ineffective and accompanied by a multitude of side effects. The gut microbiota is the reservoir of bacteria inhabiting our gastrointestinal tract. The gut microbial alterations observed in individuals with autism spectrum disorder, including elevated levels of Bacteroidetes, Firmicutes, and Proteobacteria, as well as reduced levels of Bifidobacterium, provide a basis for further investigation into the role of the gut microbiota in autism spectrum disorder. Recent preclinical studies have shown favorable outcomes with probiotic therapy, including improvements in oxidative stress, anti-inflammatory effects, regulation of neurotransmitters, and restoration of microbial balance. The aim of this review is to explore the potential of probiotics for the management and treatment of autism spectrum disorder, by investigating insights from recent studies in animals.
Introduction
Autism spectrum disorder (ASD) is a neurodevelopmental disorder characterized by repetitive behaviors and impaired social interaction (Genovese & Butler, 2020). Currently, there is a notable rise of approximately 2% in the number of individuals diagnosed with ASD compared to earlier statistics (Romano et al., 2021). Furthermore, evaluations indicate that the occurrence of this disorder is more prominent in boys than girls (Behaviors & Bergmann, 2020). In ASD children, the disorder can be accompanied by alterations in the gastrointestinal (GI) system. In fact, evidence suggests that GI dysfunction may sometimes manifest before the appearance of neurological symptoms (Margolis et al., 2021). The signs of ASD become noticeable during early life, with symptoms such as avoiding eye contact and repetitive behaviors, playing a role in facilitating the diagnosis (Behaviors & Bergmann, 2020). The gut microbiota of individuals with ASD has shown alterations, including a change in the Bacteroides/Firmicutes ratio, along with an increase in Lactobacillus and Desulfovibrio (Ng et al., 2019). The factors contributing to ASD are both genetic and environmental (Genovese & Butler, 2020). Although significant progress has been made in identifying genes associated with ASD, these efforts have yielded limited results (Alharthi et al., 2022). Furthermore, exposure to environmental factors such as certain chemical substances, air pollution, stress, alterations in the microbiota, and infections during pregnancy or after birth could all be involved in the pathogenesis of ASD (De Angelis et al., 2015).
Neurological disorders and inflammatory conditions can be accompanied by an imbalance in gut microorganisms. The depletion of the microbes in the murine intestinal tract could lead to organ dysfunction, particularly within the central nervous system (CNS), likely through the so-called “gut-brain axis” (Srikantha & Mohajeri, 2019). Moreover, the microbiota-gut-brain axis influences neurological disorders by regulating neurotransmitters such as dopamine and serotonin (Wiktorczyk-kapischke et al., 2022). Dopamine production has been detected, for example, in Staphylococcusss in the human intestine which can take up the precursor l-3,4-dihydroxy-phenylalanine (l-DOPA) and convert it into dopamine by staphylococcal aromatic amino acid decarboxylase, encoded by the gene SadA (Luqman et al., 2018). Concerning serotonin, in the gut enterochromaffin cells take up tryptophan from the diet as substrate to synthesize serotonin, and this process is finely regulated by the bacterial kynurenine synthesis pathway. Predominantly Clostridia promote the biosynthesis of serotonin by increasing the gene expression of its rate-limiting enzyme Tryptophan hydroxylase 1 (TPH1) in colonic enterochromaffin cells (Legan et al., 2022). Serotonin production has also been detected in staphylococci, which use SadA to decarboxylate the precursor 5-hydroxytryptophan (5-HTP) into serotonin (Luqman et al., 2018).
The gut microbiota also modulates CNS function through other pathways, such as the vagus nerve and the production of short-chain fatty acids (SCFAs) (Silva et al., 2020). Gut microbes also play a role in preventing the translocation of lipopolysaccharides (LPS), a bacterial toxin, into the bloodstream (Liu et al., 2019).
Considering the complexity of the pathogenesis of ASD and its spectrum of symptoms, along with the fact that most pharmacological treatments result in a wide range of side effects (Matson & Hess, 2011), an effective therapy remains to be fully discovered (He et al., 2023). Researchers are now uncovering a correlation between gut microbes and the CNS by investigating the effects of probiotics on ASD and other mental disorders (Alharthi et al., 2022). The definition of probiotics by the World Health Organization (WHO) is “live microorganisms that, when administered in adequate amounts, confer a health benefit on the host” (Zendeboodi et al., 2020). The gut microbiota, vital for the physiology of the host, is mainly influenced by lifestyle. A well-balanced microbiota greatly influences the regulation of the GI tract and CNS (Snigdha et al., 2022). Given that probiotic supplementation plays a critical role in balancing the microbiota and thereby ameliorating the symptoms of ASD, this approach has received more and more focus in both experimental models and human investigations. In this review, we not only acknowledge the role of the microbiota-gut-brain axis in mental disorders and ASD but also focus on how probiotics might affect ASD in various animal models within the context of recent preclinical studies.
Probiotics for mental disorders
Probiotics refer to “live microorganisms which when administered in adequate amounts confer a health benefit on the host” as defined by the WHO (Kumar et al., 2015). These microorganisms should be nonpathogenic, and they could be normal flora (Islam, 2016). Bacteria from the genera Lactobacillus, Lactococcus, Streptococcus, and Bifidobacterium are among many species that are used as probiotics, for prevention and treatment of many disorders, and as an alternative to antibiotics or anti-inflammatory medications (Golbaghi et al., 2022; Zaib et al., 2023).
Looking at how probiotics influence the gut-brain axis, we conclude that they have several benefits for neurological and psychological disorders. Among these disorders, depression is one of the most common psychiatric conditions, and studies indicate that transplanting fecal microbiota from depressed subjects to healthy mice can induce depressive-like symptoms, suggesting that the microbiota plays a crucial role in this depression (Jang et al., 2021). Bipolar disorder, another serious psychiatric condition, also shows an association with the gut microbiota. Lactobacilli appear to be increased in bipolar disorder (Million et al., 2013). Anorexia nervosa, another common psychiatric disorder, seems to be linked to microbiota peptide signaling. The balance between anorexigenic hormones and orexigenic peptides is dependent on the interactions between the gut microbiota and the CNS. For example, gut microbial alterations affected appetite and feeding behavior in a piglet model where the lysine restriction-shaped microbiota was associated with decreased circulating satiety hormones and increased food intake (Yin et al., 2017). The gut microbiota and appetite are highly associated and microbial metabolites may serve as a potential mechanisms. Indeed, a review has summarized the integrative homeostatic model of appetite control related to the gut microbial metabolites (Fetissov, 2017). This, in turn, could have an effect on the feeding patterns in ASD (refer to Margari et al., 2020 for a recent review on this topic).
Moreover, anxiety and depression in anorexia nervosa patients can lead to gut inflammation through microbial changes (Navarro-Tapia et al., 2021). Since the gut microbiota plays a crucial role in mental disorders and probiotics can reduce anxiety symptoms and improve mood, it is plausible to hypothesize that probiotics may also have a favorable effect on ASD. The impairment of the gut microbiota appears to be a fundamental piece in the puzzle of ASD, as a microbiota imbalance can cause changes in the activation of microglia, the main brain-resident macrophages. Central inflammation and changes in neurogenesis have been recorded in ASD, suggesting a modulation of microglial function in this disorder (Davoli-Ferreira et al., 2021).
The microbiota–gut–brain axis in ASD
According to the notion of the “gut-brain axis,” a bidirectional bridge between the brain and the gut, any damage at intestinal level could potentially lead to the onset or development of neurological and neuropsychiatric disorders (Socała et al., 2021).
Recently, researchers have shown an increased interest on the impact of probiotics on symptoms of ASD (X.-J. Kong et al., 2021; Sivamaruthi et al., 2020) and much effort has been dedicated to the study of animal models (Adıgüzel et al., 2022; Mintál et al., 2022; Qiu et al., 2023; Sgritta et al., 2019).
To gain insights into the effects of probiotics on an ASD model, it is essential to have a comprehensive understanding of the microbial populations in the GI tracts of both humans and laboratory animals. In healthy individuals, the gut microbiota comprises four primary phyla: Bacteroidetes, Firmicutes, Proteobacteria, and Actinobacteria (Fattorusso et al., 2019). According to a recent meta-analysis (Iglesias-Vázquez et al., 2020), there is a significant increase in the abundance of the first three phyla in individuals with ASD compared to healthy individuals. However, regarding Actinobacteria, there was no difference in abundance between healthy and ASD individuals. In parallel with this, there are less dominant phyla such as Fusobacteria and Verrucomicrobia (Fattorusso et al., 2019) the abundance of which is not influenced by ASD (Iglesias-Vázquez et al., 2020). Inconsistencies arise regarding the Bacteroidetes/Firmicutes ratio; this ratio has been observed to increase in some studies (Qiu et al., 2023; M. Zhang et al., 2018) and decrease in others (Ha et al., 2021; Q. Kong et al., 2021; Strati et al., 2017) among either ASD patients or animal models of ASD. These inconsistencies appear to be influenced by various factors, including environmental and dietary habits. In this regard, the ability of probiotics to restore a microbial balance whether through elevation (Adıgüzel et al., 2022; Q. Kong et al., 2021; Sgritta et al., 2019) or reduction (Qiu et al., 2023) of the Bacteroidetes/Firmicutes ratio toward baseline, is of great importance. Research indicates that there is an elevated presence of bacteria belonging to the Ruminococcus torques and Peptostreptococcaceae families in individuals with ASD when compared to healthy subjects (Chen et al., 2022; Fu et al., 2021). In addition, it has been demonstrated that the abundance of the genera Parabacteroides, Bacteroides, and Clostridium is significantly higher in individuals with ASD compared to healthy controls. Conversely, the prevalence of Bifidobacterium and Parasutterella decreases in ASD children (Iglesias-Vázquez et al., 2020).
Various pathways connect the gut microbiota to the CNS, including bacterial metabolites, oxidative stress, immune responses, neurotransmitter modulation, the kynurenine pathway, the hypothalamic–pituitary–adrenal (HPA) axis, the vagus nerve, and brain-derived neurotrophic factor (BDNF), all of which play an important role in the development of ASD (Figure 1).
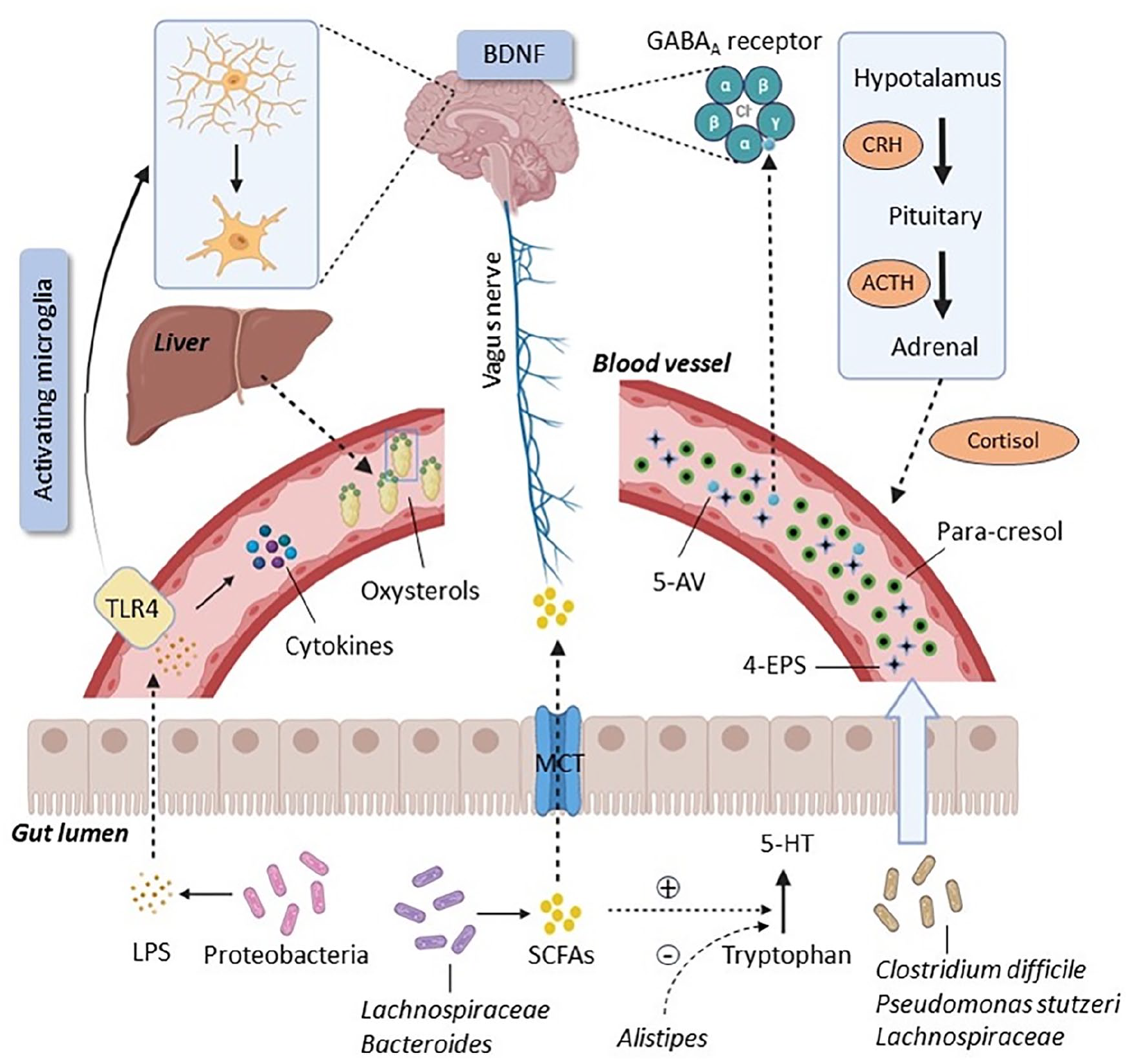
The microbiota-gut-brain axis in ASD.
One of the pathways by which the microbiota has an impact on the CNS is through metabolites production. For instance, para-cresol (4-methylphenol) is a phenolic metabolite that is mainly produced by Clostridium difficile and Pseudomonas stutzeri, and its high rate in urine is considered a biochemical marker for ASD (Altieri et al., 2011). This metabolite interferes with dopamine metabolism in the brain and was shown to worsen the social interaction of BTBR mice, a model of idiopathic autism (Pascucci et al., 2020). Lachnospiraceae, a family in the Clostridia phylum, raises the serum level of the uremic toxin metabolite 4-ethylphenyl sulfate (4-EPS), also reported to be a possible urinary biomarker for autism (Hsiao et al., 2013). Another bacterial metabolite, 5-aminovaleric acid (5-AV), which is considered a neuroactive compound due to its affinity for the GABA receptor, was decreased in the serum and intestine in offspring of mice colonized with ASD human donor microbiota (Ming et al., 2012; Sharon et al., 2019). It is worth noting that augmentation of serum levels of 5-AV may be one of the mechanisms by which Lacticaseibacillus rhamnosus attenuates social impairment in BTBR mice (Pochakom et al., 2022). Gut bacteria are also involved, via their enzymatic machinery, in bile acid metabolism. Primary hepatic bile acids such as chenodeoxycholic acid and cholic acid produced in the liver from cholesterol (Monteiro-Cardoso et al., 2021), are subsequently converted into secondary bile acids by the microbiota. Deficits in the physiological intestinal balance may therefore lead to a decrease in the level of bile acids and subsequently an increase in cholesterol, which can be converted to neurotoxic oxysterols (J. H. Kim et al., 2021). In addition, bile acid metabolism was disrupted in the BTBR mouse model of ASD, indicating a possible putative mechanism underlying GI dysfunction in these animals (Golubeva et al., 2017).
Short-chain fatty acids (SCFAs) are important beneficial bacterial metabolites. SCFAs, including butyric acid (BUT), propionic acid (PPA), and acetic acid, are generated through fermentation by gut bacteria such as Lachnospiraceae and Bifidobacterium (Y. Wang et al., 2020). These SCFAs actively participate in a range of physiological processes, including the regulation of blood pressure, circadian rhythms, and neuroinflammation (Tahara et al., 2018; Wu et al., 2021). Notably, reduced SCFA levels have been observed in certain disorders including ASD (Cryan et al., 2019; Thomas et al., 2012). However, there is some inconsistency in the field, as it has been shown that a high amount of PPA produced by Clostridium is responsible for autism-like behavior in rodents (Thomas et al., 2012). In preclinical studies germ-free (GF) mice, which are depleted of their microbiota since birth and grown in gnotobiotic isolators, showed a deficiency in microglia, and at the same time, the SCFA receptor FFAR2 was deficient in these mice, suggesting a connection between SCFAs and the regulation of microglial activity (Erny et al., 2015). By H +-dependent monocarboxylate transporters or sodium-dependent monocarboxylate transporters, SCFAs are absorbed by colonocytes, bind to their G protein-coupled receptors, and impact neuroendocrine cells by systemic circulation or the vagal pathway (Silva et al., 2020). Furthermore, an excessive amount of PPA has been widely used as a neurotoxic agent to induce experimental models of ASD in laboratory animals (Aabed et al., 2019; Abujamel et al., 2022; Mirza & Sharma, 2019). Acetate changes the levels of glutamate, glutamine and GABA in the hypothalamus (Frost et al., 2014), and PPA and BUT can alter intracellular potassium levels, impacting cell signaling (Oleskin & Shenderov, 2016). It is worth noting that both the genera Bacteroides and Desulfovibrio also have the ability to produce PPA (Duque et al., 2021).
The phylum Proteobacteria comprises gram-negative bacteria elevated in ASD that can produce the bacterial endotoxin lipopolysaccharide (LPS) (Mehra et al., 2023). LPS can indirectly influence the CNS, after crossing the impaired intestinal barrier, via increasing cytokine levels or by binding to TLR4 and activating microglia (Redinbo, 2014; Santocchi et al., 2016). Furthermore, rats exposed to LPS during pregnancy have lower levels of glutathione (GSH), an essential antioxidant in the brain (Zhu et al., 2007).
The role of oxidative stress in ASD development could be significant due to its correlation with mitochondrial dysfunction. Since children with ASD often have a higher abundance of Clostridium, there is an increased production of PPA and BUT by this bacteria, leading to a disruption of the normal mitochondrial energy production cycle (Hu et al., 2020). In addition, the presence of fatigue symptoms in certain children, along with the significant elevation in levels of oxidative stress markers, has led to the hypothesis that ASD might have an underlying mitochondrial dysfunction component (Hu et al., 2020). Furthermore, many researchers have acknowledged the attenuation of plasma levels of antioxidants such as superoxide dismutase (SOD), GSH, and L-carnosine in ASD patients (Chez et al., 2002; Söğüt et al., 2003; Yorbik et al., 2002).
Another pathway through which the gut microbiota influences the brain is the immune system. Studies have reported that the microbes within the GI tract not only have regulatory effects on central microglia and astrocytes and on immune cells within the bloodstream (Hu et al., 2020), but also affects the gut mucosal immune system through their own pathogen-associated molecular patterns (PAMPs). These PAMPs bind to the pattern recognition receptors of the intestinal immune system, resulting in the production and release of cytokines (Ansari et al., 2020). Analysis of blood serum and cerebrospinal fluid in both ASD humans and rodents reveals an activation of microglia and an increase in pro-inflammatory cytokines such as TNF-α, interleukins, and interferon-gamma (IFN-γ) (Sivamaruthi et al., 2020). In turn, a dysregulation of cytokine levels has adverse effects on behavior through the interruption of activity of neurons (Ashwood et al., 2011). The levels of inflammatory cytokines such as IL-10 and IFN-γ are diminished in the bloodstream of the PPA-induced rat model of ASD after treatment with probiotics (Alonazi et al., 2022).
The CNS can be modulated by the kynurenine pathway, which is responsible for tryptophan metabolism in mammals (Kennedy et al., 2017). Previous studies carried out on ASD children have reported alterations in downstream metabolites of tryptophan, including increased serum levels of kynurenic acid, quinolinic acid, 3-hydroxykynurenine, and the kynurenine/kynurenic acid ratio (Bilgiç et al., 2022; Bryn et al., 2017; Lim et al., 2016), with some of them having neuroactive properties (Evrensel et al., 2020). Being tryptophan the essential precursor of the neurotransmitter serotonin (5-hydroxytryptamine, 5-HT) (Young & Leyton, 2002), alterations in its metabolic pathways can lead to disruptions in serotonergic signaling.
The release of neurotransmitters into the bloodstream is another pathway of the gut–brain axis (Fattorusso et al., 2019). Several studies have indicated that 5-HT and its transporters are significantly higher in children with ASD than in healthy individuals (Abdulamir et al., 2018; Muller et al., 2016). SCFAs also participate in regulating 5-HT levels by increasing the expression of tryptophan 5-hydroxylase 1, an enzyme involved in 5-HT synthesis, in enterochromaffin cells (Reigstad et al., 2015; Yano et al., 2015). Lactococcus could potentially produce 5-HT, while Bacillus might produce dopamine in in vitro. Moreover, Lactobacillus rhamnosus could play a role in regulating GABA receptors in mice, and Bifidobacterium could be involved in the production of GABA in humans (Clark & Mach, 2016). On the other hand, Alistipes presumably due to having a gene for tryptophanase, could disrupt the 5-HT synthesis pathway by converting tryptophan to indoles (C. Zhang et al., 2015).
The HPA axis, as a part of the neuroendocrine system involved in emotion and mood, is considered one pathway playing a role in the gut–brain axis of ASD patients (Cussotto et al., 2018). To date, various mechanisms have been suggested to explain the association between the gut microbiota and the HPA axis, including an enhanced release of cytokines (such as interleukin (IL)-1β, IL-6, and TNF-α) which might pass the BBB and activate the HPA axis (Banks, 2005). In addition, the HPA axis can also be activated by the release of LPS (Vakharia & Hinson, 2005) and by E. coli, which can produce the ClpB protein that mimics α-melanotrophin (α-MSH) (Breton et al., 2016). In turn, α-MSH can stimulate the release of proopiomelanocortin that is a substrate for adrenocorticotropic hormone (ACTH) synthesis. In children with autism, plasma cortisol levels are elevated compared to healthy counterparts (Spratt et al., 2012). Also, a milestone paper by Sudo and colleagues revealed the importance of regulation of gut bacteria on the HPA axis since germ-free mice exposed to an acute stress showed a hyperactivation of this axis (Sudo et al., 2004). Moreover, treatment with Lactobacillus farciminis as a probiotic strain lowers the ACTH in the plasma of rats, suggesting the crucial role of the gut microbiota in regulating the HPA axis. More interestingly, it has been indicated that a reduced LPS level in the colon and plasma can alleviate HPA axis activity (Ait-Belgnaoui et al., 2012).
Surveys, such as that conducted by Bercik et al. (2011), have shown that alterations in the microbial population result in elevated BDNF levels in mice brains (Bercik et al., 2011). BDNF is a key protein from the neurotrophin family that helps neurons survive, grow, and differentiate (Barbosa et al., 2020). According to studies, this protein is involved in the pathophysiology of ASD (Halepoto et al., 2014). However, there is a contradiction in the BDNF level changes in the serum of ASD patients; some of them exhibit an increase (Bryn et al., 2015; Meng et al., 2017) while others exhibit a decrease (Hashimoto et al., 2006; Taurines et al., 2014). Hence, further research is needed to explore the exact pathway by which BDNF plays a role in the gut-brain axis and the pathogenesis of the disorder.
The gut microbiota communicates with the CNS via the afferent vagus nerve (Kraneveld et al., 2016). For instance, stimulation of the vagus nerve immediately after intraperitoneal administration of LPS in mice inhibited the microglia synthesizing pro-inflammatory IL-6 and TNF-α (Huffman et al., 2019). Several investigations demonstrated that probiotics could not benefit animal model of ASD following a surgical vagotomy (Bravo et al., 2011; Sgritta et al., 2019). Also in a clinical study, low vagal tone has been observed in ASD patients (Cohen et al., 2015).
Probiotics for ASD: recent preclinical findings
Although there are some differences between the composition of the gut microbiota in humans and laboratory animals, they share similarities up to approximately 90% in terms of phyla and genera (Park & Im, 2020). Preclinical studies have been conducted with the aim of providing extrapolatable data to understand the potential effects of probiotics on the improvement of ASD (Table 1).
Preclinical studies on animal models of ASD.
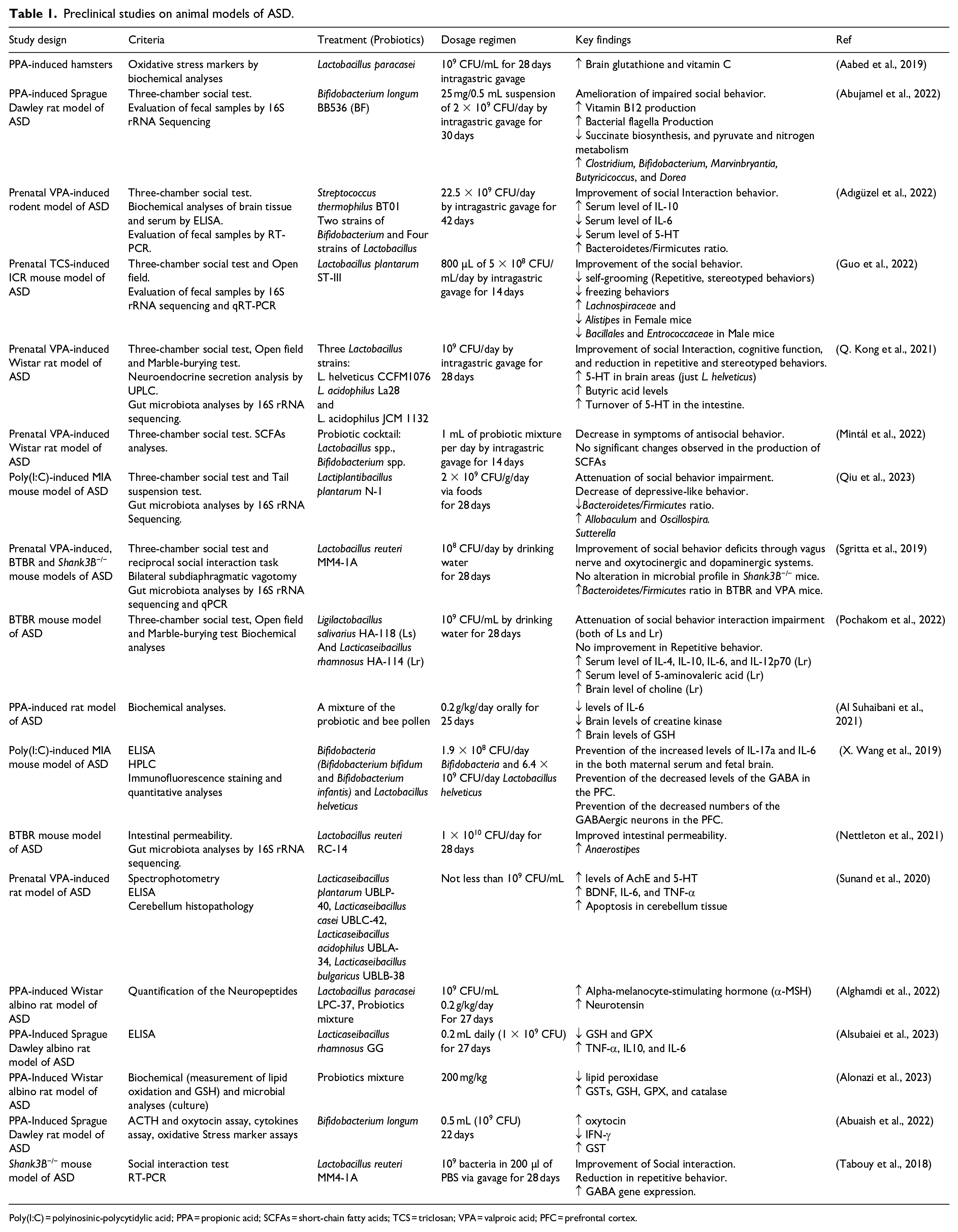
Poly(I:C) = polyinosinic-polycytidylic acid; PPA = propionic acid; SCFAs = short-chain fatty acids; TCS = triclosan; VPA = valproic acid; PFC = prefrontal cortex.
Currently, various animal models of ASD are used in research. These include genetic, idiopathic, and environmental models. Genetic models simulate ASD behaviors through different methods; for example, manipulation and mutation of certain genes such as SHANK3, can lead to the development of autism-like behaviors by causing glutamatergic transmission impairments. In addition, BTBR mice serve as idiopathic models and exhibit certain autism-like symptoms such as social deficits and repetitive behavior due to inherent anatomical differences in the brain. In environmental models, excessive prenatal exposure to a chemical compound such as valproate can induce autism-like behaviors in animals (Varghese et al., 2017).
A study performed on a PPA-induced hamster model of ASD indicated that treatment with Lactobacillus paracasei for a duration of 28 days favorably influences cerebral oxidative stress by elevating the levels of vitamin C and brain glutathione (Aabed et al., 2019).
In another preclinical study conducted on a VPA (valproic acid)-induced rodent model, the application of probiotics demonstrated anti-inflammatory impacts. This was achieved through an increase in serum levels of the anti-inflammatory cytokine interleukin-10 and a reduction in serum levels of IL-6 which is a pro-inflammatory cytokine. Furthermore, the authors reported also a decrease in serum 5-HT levels (Adıgüzel et al., 2022).
A recent study showed that prenatal exposure of mice to poly(I:C) results in behavioral changes and the manifestation of autism-like behaviors, such as increased depressive-like and impaired social behaviors. Treatment with Lactiplantibacillus plantarum N-1 for 28 days led to improvements in behavioral symptoms in the three-chamber social behavior and tail suspension tests. In addition, probiotic treatment establishes a gut microbial balance by restoring the Bacteroidetes/Firmicutes ratio, elevating the abundance of Oscillospira, and reducing the abundance of Sutterella which could be a source of LPS (Figure 2(a)) (Qiu et al., 2023).
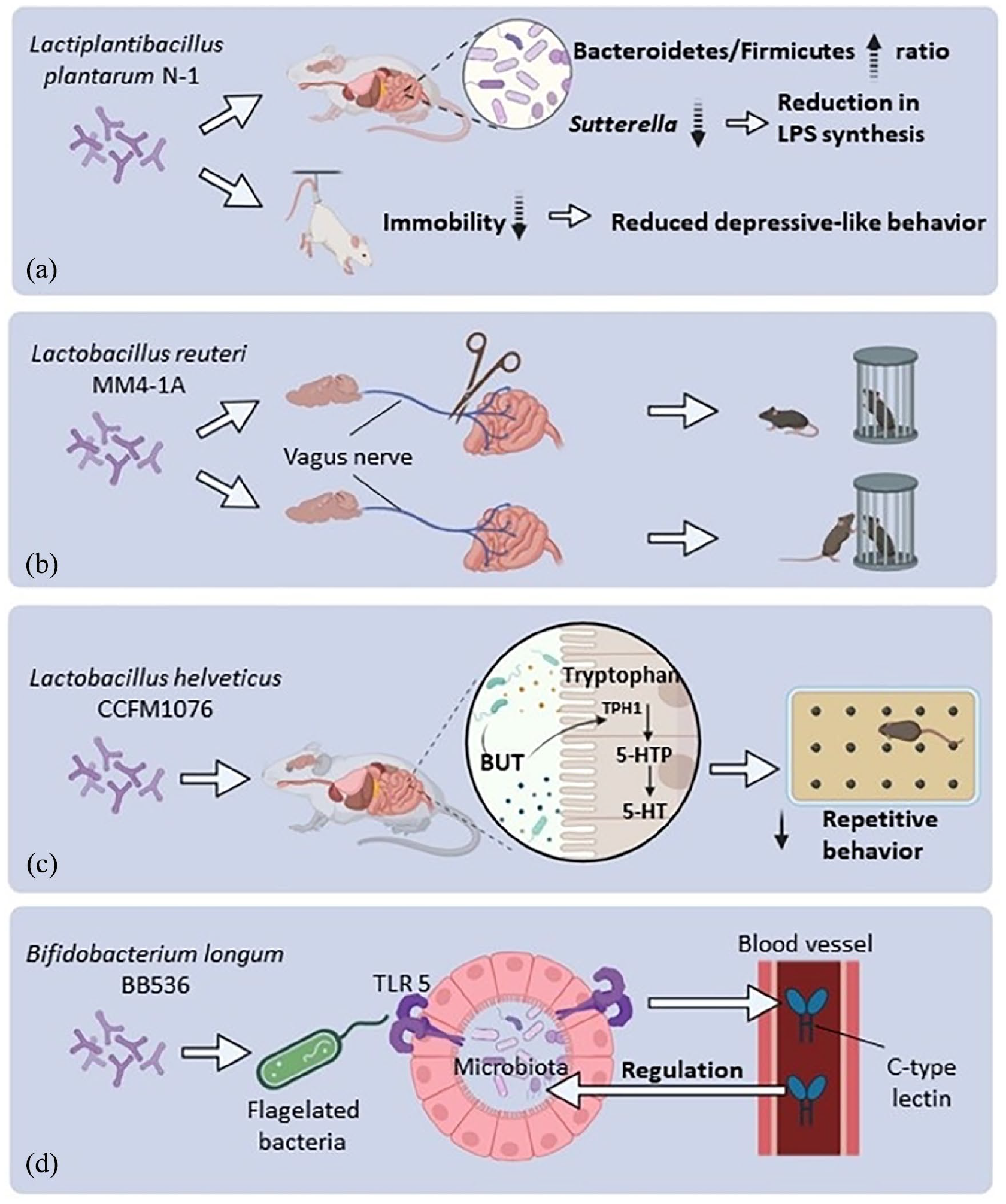
The effects of different probiotic strains on animal models of ASD. (a) Long-term use of Lactiplantibacillus plantarum increases Bacteroidetes/Firmicutes ratio and reduces Gram-negative bacteria like Sutterella that produce LPS. This treatment also lowers immobility time in the tail suspension test, reducing depressive-like behavior. (b) Lactobacillus reuteri does not improve social behavior in vagotomized mice as shown in the three-chamber test but does so in sham mice, showing the vagus nerve’s crucial role in probiotic effects. (c) Lactobacillus helveticus increases Butyrate (BUT), which may speed up serotonin production, reducing repetitive behaviors during the marble burying test. (d) Bifidobacterium longum enhances the growth of flagellated bacteria, triggering Toll-like receptors 5 and elevating blood C-type lectin levels. The antimicrobial properties of C-type lectin aid in restoring the gut microbiota balance to a physiological state.
A recent study was conducted using a genetic (Shank3B-/-) mouse model of ASD. In this model, animals were administered Lactobacillus reuteri 108 CFU/day by drinking water for a period of 28 days. The study revealed that these animals reversed their social deficits in the three-chamber and reciprocal social tests. Notably, L. reuteri’s capacity to enhance social behavior in GF mice suggests that its effects are mediated through by mechanisms beyond changes in the gut microbiota composition. L. reuteri has been found to elevate plasma oxytocin, and this increase in oxytocin is attributed to the amelioration of social behavior. Interestingly, this effect is absent when oxytocin receptors are lacking due to genetic deletion or pharmacological blockade, particularly on dopaminergic neurons in the ventral tegmental area. These findings highlight the significant involvement of both the oxytocinergic and dopaminergic systems in the enhanced social behavior induced by L. reuteri. More specifically, since there were no alterations in the gut permeability of Shank3B-/- mice, and vagotomized mice did not exhibit any sociability improvements, it has been demonstrated that the rescue of social behavior by L. reuteri operates through the vagus nerve (Figure 2(b)) (Sgritta et al., 2019).
BTBR mice exhibited impairments in social interaction and increased repetitive behaviors in behavioral tests including the three-chamber social behavior, open field, and marble burying tests. However, treatment with two species, Lactobacillus salivarius and Lactobacillus rhamnosus, for a duration of 28 days managed to reverse social interaction impairment. In addition, Lactobacillus rhamnosus could elevate serum levels of inflammatory cytokines such as IL-6 and IL-10 (Pochakom et al., 2022).
Some studies suggest that there might be a sex difference in the response to probiotics in animal models of ASD. For instance, a study investigating the impact of Lactobacillus plantarum ST-III on mice prenatally exposed to triclosan (TCA) demonstrated a reduction in repetitive behaviors, such as self-grooming, in female mice only. In addition, gut microbiota profiling using 16S rRNA sequencing revealed that treatment with this probiotic increased the abundance of Lachnospiraceae in female mice, while in their male counterparts, the abundance of Bacillales and Enterococcaceae increased (Guo et al., 2022).
Prolonged exposure to L. helveticus CCFM1076 but not L. acidophilus La28 and L. acidophilus JCM 1132 led to a notable rise in the level of 5-HT within the prefrontal cortex of rats that had been prenatally exposed to VPA. Moreover, following this treatment, there was an observed increase in the BUT concentration and the turnover of 5-HT within the intestine. This study suggests that the positive effect of L. helveticus strain CCFM1076 on behavior is likely due to the regulatory effect of BUT on 5-HT (Figure 2(c)) (Q. Kong et al., 2021).
Bifidobacterium longum BB536 has the potential to enhance the production of bacterial flagella, resulting in increased expression of C-type lectin in the bloodstream via the activation of Toll-like receptor 5. The antibacterial properties of C-type lectin enable it to promote a balanced gut microbiota which may be considered a beneficial effect of the probiotic (Figure 2(d)) (Abujamel et al., 2022).
Conclusion
Overall, several studies highlight the fundamental role of the gut-brain axis in neurodevelopment and behavior. This notion has led to an increased interest in how probiotics might help with improving ASD symptomatology by using genetic and environmental animal models of ASD. These models have revealed insights into pathways involved in the gut-brain axis including microbial populations, SCFAs, neurotransmitters and immune responses that collectively contribute to ASD. Preclinical studies have demonstrated the potential of probiotics, such as Lactobacillus strains, to modulate these links and ameliorate ASD-like behaviors, offering a beneficial impact for future therapeutic interventions. For instance, Lactobacillus paracasei reduces oxidative stress in PPA-induced hamster models, Lactobacillus reuteri improves social behavior in Shank3B-/- mice through the oxytocinergic system, and Lactiplantibacillus plantarum ameliorates behavioral symptoms in poly(I:C)-exposed mice. Notably, animal models treated with probiotics exhibit restored Bacteroidetes/Firmicutes ratios. Moreover, differences in gut microbiota abundance were observed between sexes.
In parallel and complementarily to all the preclinical findings discussed in this review, it is worth to briefly describe the most recent advances in the use of probiotics for ASD in humans. In a recent randomized controlled trial, a 6-month multi-strain probiotic therapy Vivomixx administered to 46 children with ASD, induced a decrease of power in frontopolar regions in beta and gamma bands, and increased coherence in the same bands together with a shift in frontal asymmetry. Interestingly, electroencephalography measures were significantly correlated with clinical measures such as “writing skills,” suggesting a link between probiotics supplementation and changes in brain activity (Billeci et al., 2023). A different placebo-controlled study investigated the efficacy of L. reuteri once daily for 28 days in 15 autistic participants and showed improvements in both adaptive behavior (measured by the Vineland-3 scale) and social preference as measured by eye tracking (Schmitt et al., 2023). L. reuteri was also tested in a different cohort of children with ASD, showing to induce significant improvements in social functioning but no effects on overall autism severity or on repetitive behavior (Mazzone et al., 2024).
In future research, investigating mechanisms, gender-specific responses, optimal strains, and advanced techniques could further clarify the path toward effective probiotic therapy for children with ASD. Specifically, it is important to study individual differences within the ASD population in order to better account for etiological and phenotypic heterogeneity. In addition, great emphasis must be placed on mechanistic processes and longitudinal developmental trajectories rather than endpoints. Future research should also focus on protective factors versus risk factors associated with ASD by studying the individuals at risk for ASD (e.g. infant siblings, individuals with genetic risk variants associated with ASD, and premature infants) who do not meet criteria for an ASD diagnosis. Identifying ASD protective factors, including variations in gut microbiota, will provide important information for novel gut-targeting prevention and/or invention approaches. Finally, it is important to emphasize the need for more clinical research to extrapolate from the promising preclinical findings and ensure their translation with the ultimate goal of improving the management of ASD in children.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.