
Abstract
Early acquisition of a second language influences the development of language abilities and cognitive functions. In the present study, we used functional Magnetic Resonance Imaging (fMRI) to investigate the impact of early bilingualism on the organization of the cortical language network during sentence production. Two groups of adult multilinguals, proficient in three languages, were tested on a narrative task; early multilinguals acquired the second language before the age of three years, late multilinguals after the age of nine. All participants learned a third language after nine years of age. Comparison of the two groups revealed substantial differences in language-related brain activity for early as well as late acquired languages. Most importantly, early multilinguals preferentially activated a fronto-striatal network in the left hemisphere, whereas the left posterior superior temporal gyrus (pSTG) was activated to a lesser degree than in late multilinguals. The same brain regions were highlighted in previous studies when a non-target language had to be controlled. Hence the engagement of language control in adult early multilinguals appears to be influenced by the specific learning and acquisition conditions during early childhood. Remarkably, our results reveal that the functional control of early and subsequently later acquired languages is similarly affected, suggesting that language experience has a pervasive influence into adulthood. As such, our findings extend the current understanding of control functions in multilinguals.
Keywords
Introduction
Acquisition of a second language (L2) early in life has many important consequences. It has not only been proposed to promote flexible learning of multiple, new speech items in bilingual infants (Kovacs & Mehler, 2009), but is also supposed to advance cognitive control and selection abilities in children and adults (Bialystok, 1999; Bialystok et al., 2005; Luk, de Sa, & Bialystok, 2011). Interestingly, whilst speaking, reading or listening for comprehension, bilinguals resort to mechanisms of control to avoid interference (Festman, Rodriguez-Fornells, & Munte, 2010; for a review, see Kroll, Bobb, Misra, & Guo, 2008). Thus, there is currently a need of understanding language control in multilinguals and its connection to age of acquisition (AoA). Case reports relating to bilingual aphasics have localized the brain regions that are engaged in language control and point to the importance of this facility in a communicative context. For instance, lesions in the prefrontal cortex or the anterior cingulate gyrus can result in unintentional language switching (Fabbro, Skrap, & Aglioti, 2000). Pathological mixing has been related to dysfunction of the temporo-parietal junction (Abutalebi & Green, 2007) and of the basal ganglia (Abutalebi, Miozzo, & Cappa, 2000; Marien, Abutalebi, Engelborghs, & De Deyn, 2005), and fixation to one language has been observed after lesions to the latter (Aglioti, Beltramello, Girardi, & Fabbro, 1996). Further, studies involving either intraoperative stimulation to induce language switching (Kho et al., 2007; Moritz-Gasser & Duffau, 2009), the recording of Event-related Potentials (ERPs) (Khateb et al., 2007; Rodriguez-Fornells et al., 2005) or functional magnetic resonance imaging (fMRI) studies (Abutalebi et al., 2008; Crinion et al., 2006; Hernandez, Martinez, & Kohnert, 2000; Hernandez, Dapretto, Mazziotta, & Bookheimer, 2001; Rodriguez-Fornells et al., 2005) have corroborated patient data relating to the brain areas implicated in bilingual control.
An early AoA has been reported to influence sentence production in prefrontal brain regions (Bloch et al., 2009; Kim, Relkin, Lee, & Hirsch, 1997; Mahendra, Plante, Magloire, Milman, & Trouard, 2003; for a review see Wattendorf & Festman, 2008). However, these published data leave unanswered the question of whether learning of a second language early in life influences other brain areas involved in language control. In addition, the impact of early AoA on the processing of a later learned language – indications for which have been disclosed by following an emergentist approach (Hernandez & Li, 2007; Hernandez, Li, & MacWhinney, 2005) – has never been investigated. In the present study, fMRI was used to evaluate the effect of early AoA on neuronal language representation in healthy multilinguals. We hypothesized that early AoA affects brain regions engaged in language control. Consequently, we anticipated that this effect, if generalized, would be observed during production of all languages of a multilingual repertoire. Language production at the sentence level was taken as an indicator for engagement of control, and functional activity during the use of the first (L1) and the second (L2) languages, as well during that of a third (L3), late-acquired language was obtained in early multilinguals (EMs) and contrasted with the single L1 and the two late acquired languages of late multilinguals (LMs).
Materials and methods
Inclusion criteria
From a larger cohort of multilinguals, we selected participants who have been separated on whether they learned a second language early in life, namely second language (L2) acquisition before the age of three years (EMs) and after the age of nine years (LMs). Participants in the study had learned a third language (L3) after the age of 9 years, but none between 3 and 9 years (for details pertaining to the learned languages, see Table 1). A further inclusion criterion was high proficiency in all three languages (see Table 2). Each individual was closely questioned on its biography of language learning (Franceschini 2002) and was required to complete a detailed proficiency questionnaire, which was developed based on the Common European Framework of Reference (CEFR; North, 2000). Language biographies furnished additional information on the language-learning process. The proficiency questionnaire afforded a standard for the comarison of proficiency levels across a very broad range of languages (see also Costa & Santesteban, 2004; Costa, Santesteban, & Ivanova, 2006) and comprised the following skills: comprehension, reading, writing, spoken production and spoken interaction. This approach has been validated by other investigators: it yields consistently high, overall correlations between the results of self-assessment and ratings that are based on external criteria for foreign languages (Oscarsson, 1984, 1997). Finally, eight EMs (four males) and 8 LMs (four males) satisfied the criteria for inclusion in this study.
Languages learned by early and late multilinguals.

Acquired informally only;
acquired informally with formal instruction later at school;
acquired at school; bold: first formally instructed language at school; F: female subject; M: male subject.
Proficiency levels of early and late multilinguals.

Proficiency levels of early and late multilinguals in L1, L2 and L3 according to the Common European Framework of References (CEFR) for Languages (A – basic user; B – independent user; C – proficient user) according to a six-level scale (in ascending order including the comparable Cambridge (English for Speakers of Other Languages, ESOL) Examination: A1 – Breakthrough; A2 – Waystage (Key English Test, KET); B1 – Threshold (Preliminary English Test, PET); B2 – Vantage (First Certificate in English, FCE); C1 – Effective Proficiency (Certificate in Advanced English, CAE); C2 – Mastery (Certificate of Proficiency in English, CPE).
Participants
The multilinguals included in this study (EMs and LMs) were right-handed (lateralization index >90%, according to the Edinburgh Handedness Inventory) and without a history of neurological disorders. They were between 22 and 35 years of age and had similar educational backgrounds (minimal requirement: undergraduate level).
The study was approved by the Ethics Committee of the University Hospital of Basel (Switzerland) and was undertaken with the informed written consent of each participant.
Tasks
The test condition for each of the three languages involved two tasks, which were presented in the scanner using a block design (Bloch et al., 2009): a silent sentence production task adapted from Kim et al. (1997) and a baseline task (finger tapping). During the language task, images of a sun at three different positions, symbolizing morning, noon and evening (see Figure 1) were presented to the participants, who were required to covertly formulate respective activities of the previous day in a predefined language. All participants practised the task in all three languages aloud before the scanning session. They received extensive feedback; in particular, we explained to the participants naïve to imaging techniques that the quality of the data depended upon their co-operation during the task. To remain in a monolingual mode (Grosjean, 2001), that is, to suppress all other languages except the one currently tested, the participants were asked, prior to each test, to choose an imagined interlocutor, a person to whom they would speak only in this language. The display of a cross indicated the baseline task during which the subjects responded with a single finger tap of the right hand to silent periods of the scanning sound (every 6 s). The main purpose of this condition was to sustain attention rather than to execute the motor act. For subtractive designs, such auditory conditions are superior to an uncontrolled resting state, since they engage fewer linguistic processes (Binder et al., 1999). Visual cues for each task (sun versus cross) were displayed for periods of 30 seconds and repeated 9 times. In the break after each session, participants were asked to choose a new interlocutor and to prepare for the expression of a different language. Thus, this task entailed language switching only between, not within, tasks. After the scanning session, each participant was asked to report on his/her performance during the scanning session. No specific difficulties in performing the task in any of the languages were revealed.
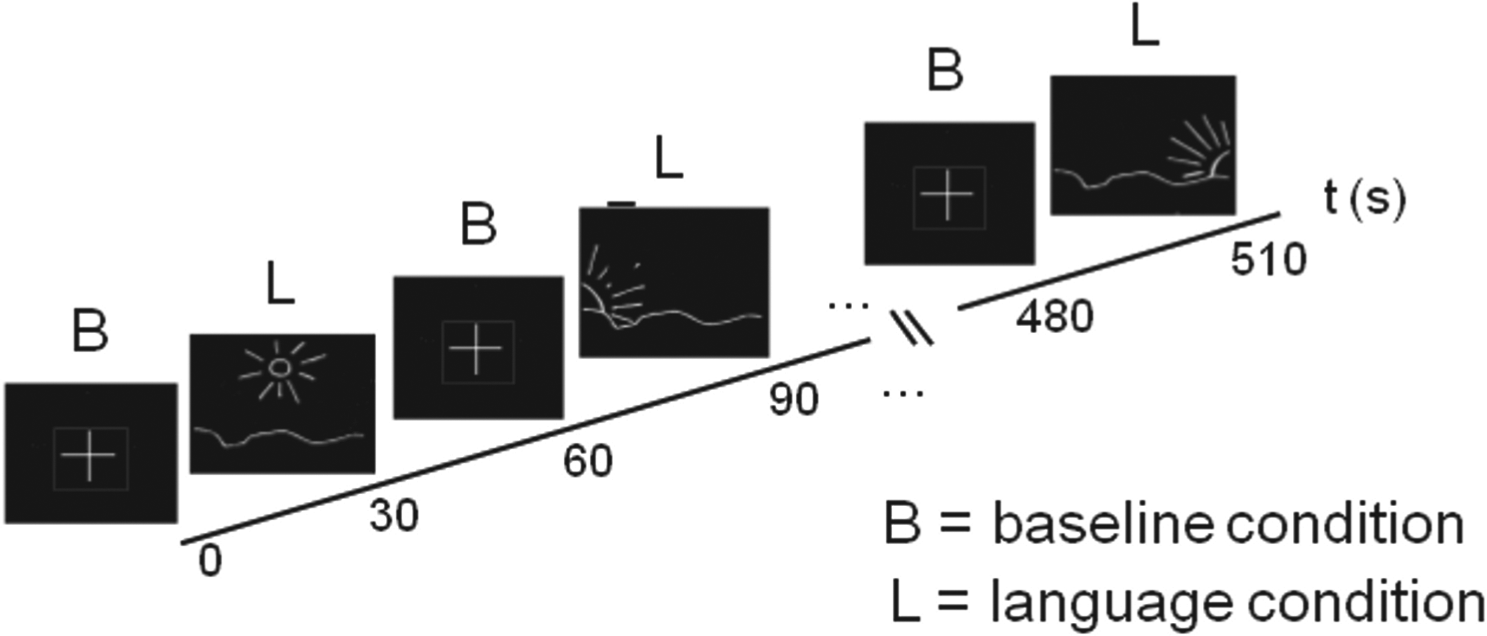
Experimental set-up for fMRI. The stimuli for the language condition are represented by images of a sun at different positions, symbolizing morning, noon and evening; they serve as signals to the participants to relate the activities of the previous day. The auditory baseline condition was represented by a simple cross. To prevent the participants from preparing in advance for the language condition, the order of visual language stimuli was randomized across different runs. Each symbol was projected for 30 s and repeated 9 times.
Taking into account that variability in language-related activity between scanning sessions may confound the study of languages in bilinguals (Mahendra et al., 2003), sessions were repeated for each participant on different days. This measure was preferred over increasing of the group size. The order of languages during the sessions was randomized.
MRI data acquisition and analysis
Imaging was performed on a 1.5 T Magnetom Vision MRI scanner (Siemens, Germany). Functional images were obtained using a T2*-weighted, echo planar imaging (EPI) sequence (repetition time: 6 s, echo time: 60 ms, flip angle: 90°) that covered the entire brain with 48 contiguous axial slices (resolution: 3 mm × 3 mm × 3 mm). Each language was tested in a separate session. For each tested language, 90 scans were collected, comprising 9 blocks per condition. Each block included 5 scans. After the completion of the fMRI sessions, a high-resolution, structural, T1-weighted image (resolution: 1 mm × 1 mm × 1 mm) was obtained using an MP-RAGE (magnetization prepared rapid acquisition gradient echo) sequence. The data were processed and analyzed using Statistical Parametric Mapping (SPM99) software (Friston et al., 1995). Functional images of each session were realigned to correct for movements, co-registered to the structural image, normalized to the standard brain template and smoothed by an 8-mm isotrophic Gaussian kernel. In EMs and in LMs, the group-specific language-related activity (main language effect) for L1, L2 and L3 was revealed by contrasting the language with the baseline condition in a fixed-effect model - necessitated by the small group size - which included both tested runs (p < 0.05, corrected, 5 voxels). To reveal the effect of early AoA on language processing, the comparison between EMs and LMs was made for languages that had been learned at the same age and to the same degree of proficiency (differential effect of EML1/LML1, EML2/LML1 and of EML3/LML3, p < 0.0001, uncorrected, 10 voxels). To account for inter-run variability, the main effects and differential effects reported in this study were masked (inclusively) by a conjunction of both tested runs at a statistical threshold of p < 0.001 (uncorrected) in the inter-group comparisons. This procedure ensured not only a significant activation of the revealed brain regions during both runs, but also an exclusion of those exhibiting significant differences between the two. Parameter estimates of Blood-Oxygen-Level-Dependent (BOLD) signals in EMs and LMs were extracted from the peaks of differential activation. The results of the analysis of selective activation for language and baseline stimuli are graphically depicted in Figure 3c. They reveal that our findings were not attributable to baseline differences. Activated brain regions were recorded as activation maxima together with the z-score and the coordinates in the Talairach and Tornoux space (Talairach & Tournoux, 1988).
Results
Proficiency self-evaluation test and language biography
The mean ‘global proficiency’ scores indicated that all subjects reached at least an advanced proficiency level of B2/C1 according to the classification of the CEFR for Languages (see Table 2). The highest level of proficiency was revealed for early-acquired languages. Languages acquired at a similar age were mastered to a comparable level of proficiency in both groups (EML1/LML1, EML2/LML1 and EML3/LML3). Moreover, in two sub-tests of the CEFR classification, namely, spoken interaction and spoken production, which are the most relevant for the performance of the narrative task tested during fMRI scanning, EMs and LMs attained at least effective proficiency (C2), if not mastery (C1). An evaluation of the language-learning biography confirmed the regular use of all three languages by each participant. It likewise confirmed that all subjects satisfied the group-specific selection criteria, which included the circumstance that no further languages had been learned between the ages of 3 and 9. Most of the EMs (6 out of 8 participants) acquired L1 and L2 simultaneously. The L1 was determined according to its greater importance (dominance) during the early-acquisition process. However, since heightened activity in EMs (as compared to LMs L1) was not only observed in the L1 but also in the L2, early language dominance appeared not to influence our results. In LMs, the languages learned late were classified as an earlier (L2) and later (L3) language. Language biographies also provided information on the mode of language acquirement, thereby permitting a distinction between spontaneous, unstructured (informal) learning and formal instruction at school (Table 1). In two participants the first language – notably Swiss German – was not formally taught and written at school. Indeed, in the German part of Switzerland, the language used for formal language instruction is German rather than Swiss German, particularly for the written part of the language. Whether and, if so, in which respects, dialects such as Swiss German are linguistically independent are still a matter of debate. Since only three Swiss German-speaking subjects participated in the present study, a significant impact on the fMRI-data seems unlikely.
FMRI investigation: General pattern of language-related activity in EMs and LMs
In EMs brain activity was investigated during the expression of the two languages that had been acquired early (L1, L2) and the one language that had been acquired late (L3). In LMs, brain activity was investigated during the expression of the one language that had been acquired early (L1) and the two languages that had been acquired late (L2, L3). By contrasting sentence production with the auditory baseline condition, cortical areas involved in the language task were revealed (Table 3 and Figure 2): in both groups and for all languages, left-lateralized neuronal activity was observed in the dorsolateral premotor (BA 6) and supplementary motor areas (medial BA 6), in Broca’s area (BA 44/45), in the superior temporal sulcus (BA 21/22), and in the inferior parietal lobe (BA 40/7). Bilateral activity was present in the supplementary motor areas (medial BA 6), in the posterior cingulate gyrus (BA 23/30/31) and in the visual cortex (BA 17, 18, 19). Cerebellar activity was consistently registered in the right hemisphere. Activity in the corpus striatum andthe inferior frontal gyrus (BA 47) was observed in all languages of both groups, excepted in LMs for L1.
Task–related L1, L2 and L3 activation in early and late multilinguals.
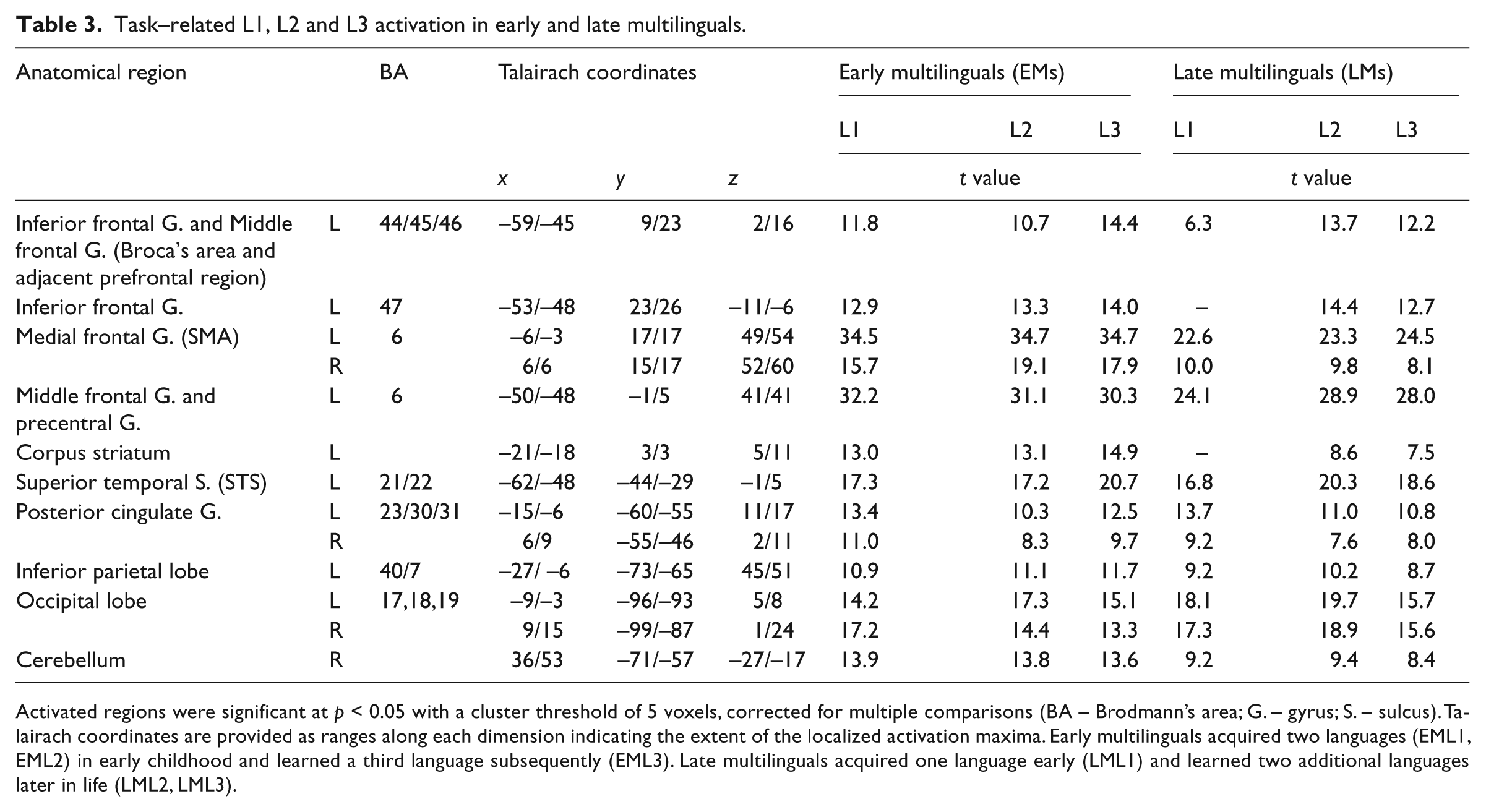
Activated regions were significant at p < 0.05 with a cluster threshold of 5 voxels, corrected for multiple comparisons (BA – Brodmann’s area; G. – gyrus; S. – sulcus). Talairach coordinates are provided as ranges along each dimension indicating the extent of the localized activation maxima. Early multilinguals acquired two languages (EML1, EML2) in early childhood and learned a third language subsequently (EML3). Late multilinguals acquired one language early (LML1) and learned two additional languages later in life (LML2, LML3).
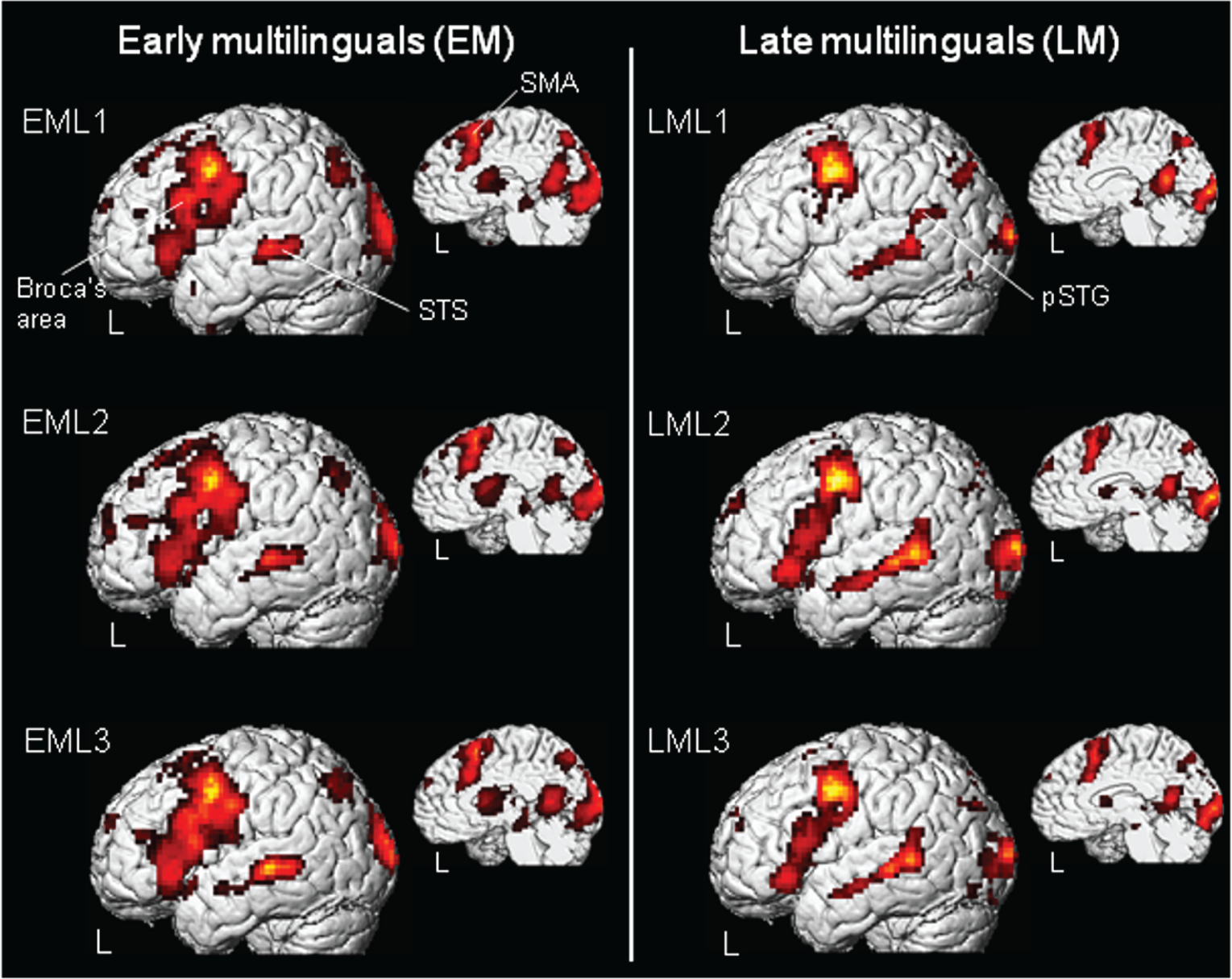
Group average language-related activity in EMs, left panel and LMs right panel multilinguals during processing of early-acquired languages (EML1/L2, LML1) and of late-learned languages (EML3, LML2, LML3). Activated brain regions were rendered on a three-dimensional brain showing lateral and medial views of the left hemisphere. In EMs and LMs, left-lateralized activation was revealed in Broca’s area (BA 44/45), the supplementary motor area (SMA, BA 6), the superior temporal sulcus (STS, BA 21/22) and in the posterior, superior temporal gyrus (pSTG, BA 22) (p < 0.05, corrected for multiple comparisons; based on the data gleaned from both test-runs).
FMRI investigation: Comparison of language-related activity between EMs and LMs
Group comparisons (Figure 3, (a)–(c) and Table 4) revealed significantly higher neuronal activity for EMs in both the frontal and subcortical regions of primarily the left hemisphere. This phenomenon was evident for languages that had been acquired both early and late, as illustrated for EML1/LML1 and EML3/LML3 in Figure 3, a & b (left panel). The regions of heightened activity included Broca’s area (BA 44/45, 44/9 and 44/6) and, to a lesser extent, its right homologue (BA 44/45 and 44/9), the left and right supplementary motor areas (SMAs, BA 6), the left anterior cingulate gyrus (BA 32), and the left striatum. In EMs, heightened activity in the left dorsolateral, prefrontal cortex (BA 46) was observed only for the languages that had been acquired early (L1 and L2); heightened activity in the left orbitofrontal BA 47 occured only for L1. Increased activity was registered in the higher visual-association areas of the cuneus (BA 19) of both hemispheres during the expression of all three languages. An increased response was observed also in the left fusiform gyrus (BA 19/37), but only during the processing of the early acquired languages (L1 and L2).

Comparison of the brain regions that were activated in EMs and LMS during the processing of early (EML1 versus LML1) and late (EML3 versus LML3) languages (p < 0.0001, uncorrected; based on the data gleaned from of both tested runs). (a) Group differences for early acquired languages (EML1 versus LML1). In EMs (left panel) heightened language-related activity was revealed in the left fusiform gyrus (BA 37/19), in the left inferior frontal gyrus (BA 44/45, Broca’s area), in the left middle frontal gyrus (BA 46), in the left corpus striatum and in the bilateral supplementary motor area (SMA, BA 6). Note that the superior temporal gyrus/sulcus (BA 21/22) has been revealed due to baseline differences and will therefore not be discussed. In LMs, only the left posterior, superior temporal gyrus (pSTG, BA 22) was highlighted. (b) Group differences for late-acquired languages (EML3 versus LML3). Note that the activation pattern is similar to that in (a) (EML1 versus LML1), with the exception that no group-specific activity was observed in either the left fusiform gyrus or the left BA 46. (c) Parameter estimates extracted from peaks of differential group activation reveal selective activation for language and for baseline tasks. Language-related differences between EMs and LMs in lefe frontal and lefe temporal brain regions are illustrated.
Differential language-related activation in early and late multilinguals.
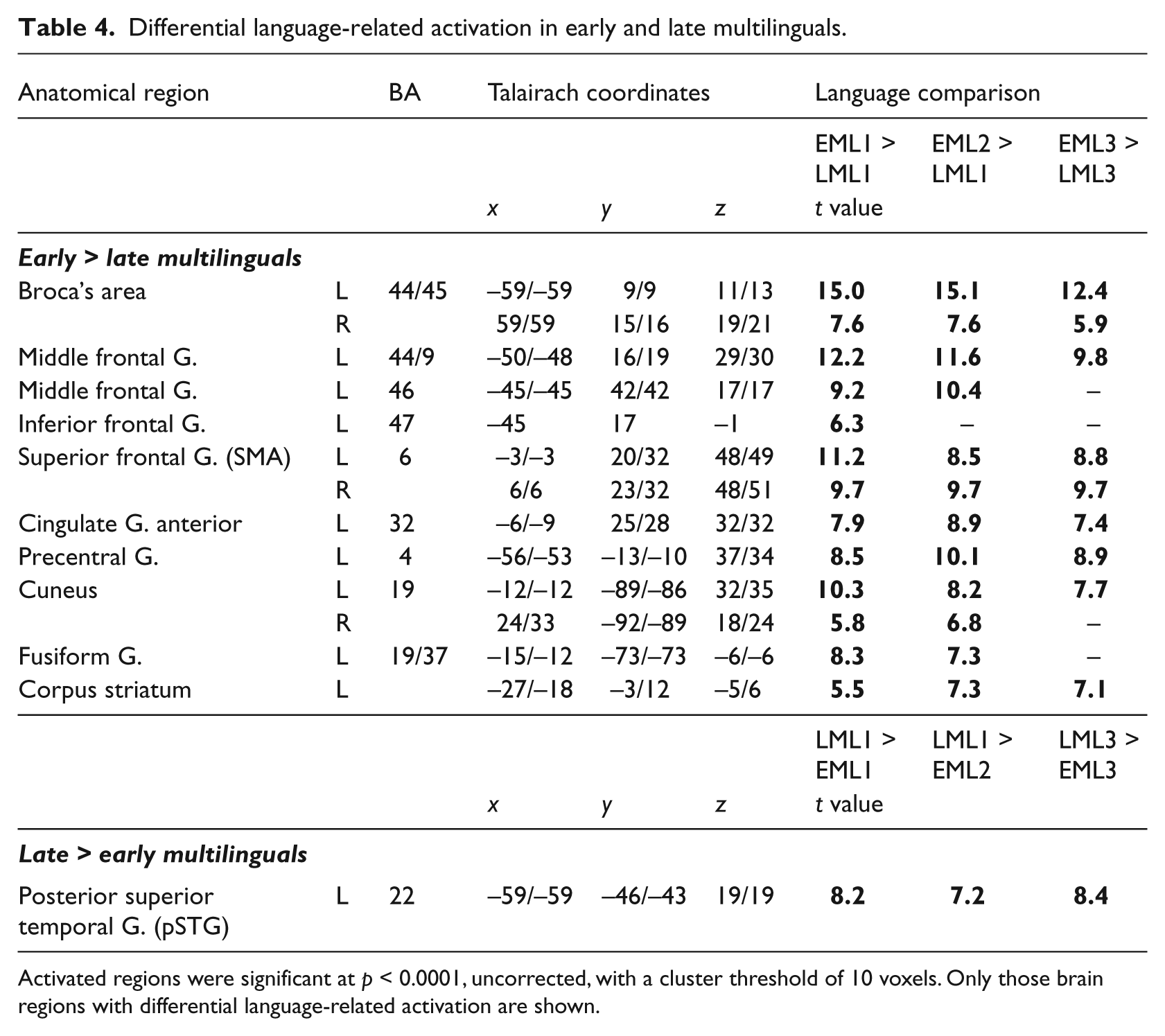
Activated regions were significant at p < 0.0001, uncorrected, with a cluster threshold of 10 voxels. Only those brain regions with differential language-related activation are shown.
LMs, in contrast, exhibited only minor increases in language-related activity compared with EMs. Only in the posterior portion of the left superior, temporal gyrus (pSTG, BA 22) did neuronal activity exceed that in EMs for all comparisons, as illustrated for the LML1/EML1 and LML3/EML3 comparisons in Figure 3, a & b, right panel. Parameter estimates of the BOLD signal change in the left Broca’s area and prefrontal cortex (BA 44/9 and BA 46) and in the posterior superior temporal gyrus (pSTG (BA 22)) (Figure 3c) revealed selective activation for the language and the auditory baseline condition, thereby demonstrating the result to be unbiased by neuronal activityduring the baseline condition.
Discussion
In both, EMs and LMs, brain areas that are commonly involved in sentence processing were activated. However further analysis of the data highlighted the specific differences between the two groups, which point to differences in the involvement of control functions. Interestingly, the AoA effect had an impact on subsequently-learned languages irrespective of whether these had been acquired early or late, which is in the line with the emergentist theory (see below).
EMs and LMs: Activated brain regions during a narrative task
In multilinguals, language control and selection mechanisms are particularly necessary during sentence processing (Festman Rodriguez-Fornells & Münte, 2010). The same brain areas that have been implicated in bilingual language control on the basis of clinical observations in neurological patients and imaging studies (Abutalebi & Green, 2007) were activated in EMs and LMs during the performance of a narrative task. In both EMs and LMs, and for each of the three tested languages, heightened activity was observed in the left prefrontal cortex, the left and right anterior cingulate gyrus, the left pSTG, the left striatum (except for L1 in LMs) and the bilateral viaual cortex. For the most part, these findings are in line with previous studies that have adressed visually cued sentence production in multilinguals (Awad, Warren, Scott, Turkheimer, & Wise, 2007; Golestaniet al., 2006; Mahendra et al., 2003; Sassa et al., 2007). Yet, in these previous studies, striatal activity was demonstrated in only a few subjects (Mahendra et al., 2003).
EMs: Higher neuronal activity in prefrontal and subcortical areas
A comparison of language processing in EMs and LMs revealed an effect of early bilingualism on the activity of the inferior frontal gyrus (mainly Broca’s area, see Table 4). This finding confirms the results of previous studies (Bloch et al., 2009; Kim et al., 1997; Mahendra et al., 2003; Wartenburger et al., 2003). Remarkably, however, heightened activity was observed also in brain regions that have not been hitherto implicated in the effects of early bilingualism, namely, the right homologue of Broca’s area, the bilateral supplementary motor area (BA 6), the left striatum, the left anterior cingulate gyrus and the left precentral gyrus. In studies on bilinguals, this network is engaged when the target language has to be selected (e.g. in picture-word interference and switching tasks). During picture-naming under conditions in which phonological interference from the non-target language pertains, the reaction times were longer and the neuronal activity was higher in early bilinguals than in monolinguals. The brain regions implicated were the left middle frontal gyrus, the SMA and the anterior cingulate gyrus (Rodriguez-Fornells et al., 2005). Furthermore, an fMRI analysis of cerebral activity in Spanish-English bilinguals during mixed-language naming compared to naming in only one language has revealed higher activity in the left dorsolateral prefrontal regions (Hernandez, Martinez & Kohnert, 2000; Hernandez et al., 2001). Moreover, tasks that predominately challenge the parallel activation of both languages, such as semantic priming in the non-target language (Crinion et al., 2006) and translation (Lehtonen et al., 2005; Price, Green, & von Studnitz, 1999), lead to heightened activity in prefrontal regions and the striatum. These findings indicate that subcortical regions are also involved in the complex mechanisms subserving the selection of languages. Interestingly, the aforementioned studies indicate that the control mechanisms were engaged during tasks in which they were actively induced, for example, when two languages had to be alternatively enhanced and suppressed (switched). Concerning our own ‘free’ narrative task, it has to be pointed out that the multilingual participants were presumed to remain in a monolingual mode (Grosjean, 2001), since languages were tested one after the other and not intermingled. A further advantage of the continuous sentence production is that it involves no additional ‘external’ control tasks, such as the search for an appropriate lexical item (e.g. verbal fluency when searching for words beginning with a certain letter, or picture naming when searching for a suitable target word). Thus, using our production task, we were able to describe the effect of early AoA on brain regions that subserve control as being inherent to natural language production. Since EMs were confronted with the management of two languages early in life, our findings implicate functions that are associated with this ‘early acquisition modus’, at least so far as these concern the processing mechanisms underlying language selection (de Bot, 1992; Green, 1998). Hereby we suggest an extension of the monolingual-bilingual mode dichotomy that has been proposed by Grosjean (2001).
In EMs, early developed functions can be recruited, albeit with limitations, for late-learned languages (see Table 4 and Figure 3). In fact, in this group, the heightened activity in the left middle frontal gyrus (BA 46) and in the left fusiform gyrus (BA 19/37) occurred only for languages that had been acquired early, not for those that had been learned late. Neuronal activity in both cortical regions is known to increase during retrieval of contextual information that is associated with verbal stimuli (Dobbins, Foley, Schacter, & Wagner, 2002; Lepage, Habib, Cormier, Houle, & McIntosh, 2000). When languages are acquired later in life, earlier established functions perhaps are not always recruited, especially in those that interact with more general concepts. In EMs, heightened activity occured also in the precentral gyrus (BA 4). In this region, neuronal processes are related to articulatory levels of language processing (Wildgruber, Ackermann, & Grodd, 2001), indicating that early bilingualism induces also a modulation of processes that are implicated in the motor control of speech.
LMs: Higher neuronal activity in the pSTG
In LMs, higher activity than in EMs was registered in the pSTG for all three languages (see Table 4 and Figure 3). Consistent with the finding that this cortical region is activated not only during verbal-retrieval and speech-perception tasks but also during verbal-fluency ones (Buchsbaum, Hickok, & Humphries, 2001; Price et al., 1996; Wise et al., 2001), it has been proposed that the transient phonological storage of sound-related material in portions of the pSTG serves as an interface between speech perception, lexical recall and speech production (Buchsbaum et al., 2001; Wise et al., 2001). It thus appears that LMs show enhanced control of transient phonological representations. Since EMs seem to develop distinct phonological representations early in life (Bosch & Sebastian-Galles, 1997), the control of phonological recall processes may be less important in these participants.
Processing of early-and later-acquired languages in an emergentist framework
The present study reveals an effect of early age of acquisition (AoA) on brain language representation. This finding accords with those of published studies that have investigated a comparable ‘natural’ language production task in multilinguals (Bloch et al., 2009; Kim et al., 1997; Mahendra et al., 2003). These previous studies described the AoA effect as spatially less separated (Kim et al., 1997), quantitatively higher (Mahendra et al., 2003) and statistically less variable (Bloch et al., 2009) activation of the EMs’ two early acquired languages (L1 and L2) as compared to the LMs’ early (L1) and late (L2) acquired languages. Here, we compared not only L1 and L2, but also a third, later learned language (L3) between the two groups. The analysis pertaining to L3 revealed similar results to those for L1 and L2, which points to a pervasive effect of early AoA on brain functions.
An explanation of this finding may be sought in the emergentist theory of multilingualism. The emergentist theory (see, for example Hernandez & Li, 2007) holds that only early bilinguals learn both languages in parallel, with each language slowly developing into separate cortical processing maps, which are turned into consolidated language modules. The existence of such consolidated language modules implicates selection-mechanisms which lead to the development of a more efficient language processor (O’Grady, Kwak, Lee & Lee, 2011). It may explain why early experience of two languages has an impact on processing of any later learned language, as revealed in our study. The effects observed in EMs may result from the type of early predominant sensori-motor learning (sensori-motor loops involving the basal ganglia and the prefrontal cortex). Since LMs cannot rely on these early established modes of consolidated language processing, other brain areas are recruited to support their less efficient processor. In the tested task this was the pSTG, which is primarily involved in phonological working memory.
Other variables: Age limit, language typology and proficiency
In this study, we focussed on the AoA of second language as a factor for distinguishing between the language systems of EMs and LMs. Considering that bilinguals can master basic language skills in two languages already at the age of 3 years, which implies that the underlying cognitive functions are then mature (Meisel, 2008), we included only participants who started to use L2 before or at this age. Since the participants had not acquired any other language between the ages of 3 and 9 years, we were sure of capturing the influence of AoA on language functions during early childhood. Owing to the stringent selection criteria, the number of participants in each group was small (n=8).
Other parameters pertaining to language and its learning, such as language typology and proficiency, could also influence its neuronal representations in multilinguals. In our study, a broad range of languages (Table 1) was included. Although language type has been shown to have an impact on reading strategies (Paulesu et al., 2000; Tan, Feng, Fox, & Gao, 2001) and the processing of speech sounds (Klein, Zatorre, Milner, & Zhao, 2001), no effect on language production tasks has ever been noted (Bloch et al., 2009). Proficiency has also been a factor whose influence has been demonstrated in bilinguals. For instance, during word fluency (Vingerhoets et al., 2003), semantic and/or syntactic judgement (Wartenburger et al., 2003) and mixed language naming (Hernandez et al., 2000, 2001), lower proficiency for L2 is associated with greater prefrontal control, viz., with higher activation in the inferior frontal gyrus (BA 47) and in the middle frontal gyrus (BA 46) (De Bleser et al., 2003; Hernandez & Meschyan, 2006). Furthermore, behavioural data disclose a higher incidence of speech production errors – slip-of-the-tongue effects – in individuals who are less proficient in L2 than in L1 (Poulisse & Bongaerts, 1994). And during language switching, task-naming latencies – so-called switching costs – have been shown to be asymmetrical for the two languages of a bilingual when proficiency in one of these is poor (Costa et al., 2006). When designing our own study, we took these findings into consideration by including a language proficiency test (see Methods). Since the acquisition/learning conditions and proficiency (notably during spoken interaction and spoken production, see Table 2), were comparable for early (L1)- and late (L3)-learned languages, we are able to exclude an impact of these factors on our results. However, since a distinction was not made between high and very high levels of proficiency, an effect on our results of a very high degree of balanced bilingualism (viz, specific language control mechanisms for both languages that are independent of AoA, as reported by Costa et al., 2006) cannot be entirely excluded and needs to be further explored.
The current study represents a further step towards distinguishing the various factors that are involved in the development and use of language control in multilinguals. We propose that the early learning of two languages has a pervasive effect on a neuronal network that is presumed to regulate language control in bilinguals at different processing levels, which include even subcortical structures. Our findings might be relevant in understanding the patterns of recovery in bi-and multilingual aphasics or other behavioural changes in healthy multilinguals. On this note, the data presented in this study will promote further explorations of the AoA - effects and its influence on diverse aspects of control functions in multilinguals.
Footnotes
Acknowledgements
The authors would like to thank Dr. Klaus Fiedler and Prof. Joachim Wattendorff for their expert suggestions for improving this article and Dr. Martin Rausch for implementing the EPI-sequence.
Funding
The study was supported by the “Erneuerungsfonds” of the University of Basel, by the Institute of Anatomy and the Romance Seminar of the University of Basel and by the Institute of Radiology and the Neurosurgical University Clinic of the University Hospital of Basel. Julia Festman was supported by the Deutsche Forschungsgemeinschaft (DFG, FE 941/1-1).