
Abstract
Outgroup dehumanization figures centrally in historical intergroup violence, yet little is known about the factors that promote dehumanized perceptions of others. Drawing on research highlighting the importance of disgust-relevant social categorizations and disgust sensitivity to outgroup dehumanization, the authors hypothesized that feelings of disgust causally facilitate dehumanized social cognition. To test this hypothesis, participants (N = 94) were randomly assigned to receive inductions of disgusted, sad, or neutral emotions. We then assessed their implicit associations between animals and an arbitrary outgroup created with a minimal-groups procedure. Results showed that although all participants demonstrated dehumanizing biases, disgusted participants showed the strongest associations between the outgroup and animals (and the ingroup with humanity). Participants in the sad and neutral groups did not differ. Disgust thus appears to have the unique capacity to foster the social-cognitive dehumanization of outgroup members.
Outgroup dehumanization is arguably the most morally dangerous intergroup bias, having played a pivotal role in many wars and genocides throughout history. The infamous Nazi propaganda in Julius Streicher’s Der Stürmer depicted Jews as vermin, swine, and insects (Bytwerk, 1983). In the 1994 Rwandan genocide, the Radio Télévision Libre des Mille Collines broadcasted a call to kill the Tutsi “cockroaches,” claiming that they were a dirty race in need of extermination (Zimbardo, 2007). While the causes of these atrocities are complex, a notable commonality is the depiction of outgroups in a manner designed to evoke disgust. Disgust metaphors and elicitors figure prominently in genocidal narratives (Savage, 2007), with feelings of disgust routinely evoked to derogate enemies in times of war and to moralize the persecution of minorities (Taylor, 2007).
Why disgust? Does disgust have a unique capacity to facilitate the dehumanization processes witnessed in genocide? The consistent presence of disgust-provoking imagery in historical dehumanization suggests that disgust is especially effective at blurring the human–animal divide. To date, there have been no experimental investigations of disgust’s role in dehumanization processes. The focus of the current research was to investigate whether incidental activation of disgust facilitates dehumanized social cognition, and ultimately, the moral exclusion of outgroups.
Disgust: The Exclusionary Emotion
Disgust is widely regarded as a basic, innate emotion geared toward behavioral avoidance, exclusion, and rejection (Rozin, Haidt, & McCauley, 2008). Many aspects of disgust make it relevant to dehumanization, the most critical of which may be its close link to the distinction between humans and animals. As noted by Rozin et al. (2008), disgust elicitors are often animal-related. Animals associated with garbage, death, and disease (e.g., rodents and insects) and the “animalistic” human drives (e.g., sex, aggression, hunger, and waste elimination) are the stimuli most capable of eliciting disgust (Rozin et al., 2008). Once activated, feelings of disgust reliably evoke feelings of superiority over offending targets, who by virtue of their disgustingness, are expelled from the circle of moral regard (Pizarro, Detweiler-Bedell, & Bloom, 2006). As such, disgust guards the human–animal boundary in social cognition, playing the dual role of distancing ourselves from “lower” creatures and reaffirming our own humanity. When applied to intergroup contexts, disgust inserts a psychological boundary between us and them that humanizes us at the expense of the other.
A basic role for disgust in outgroup boundary perceptions is supported by research suggesting that foreign outgroups (Navarette & Fessler, 2006), and groups stereotyped as being low in both competence and warmth (Fiske, Cuddy, Glick, & Xu, 2002) are especially likely to elicit disgust. These competence and warmth dimensions resemble the trait dimensions that convey information about target “humanness” (Haslam, Bain, Douge, Lee, & Bastian, 2005). Disgust also plays a critical role in creating and exacerbating certain types of prejudice (Dasgupta, DeSteno, Williams, & Hunsinger, 2009; Terrizzi, Shook, & Ventis, 2010) and guiding moral evaluations of others (Schnall, Haidt, Clore, & Jordan, 2008). Finally, disgust short-circuits feelings of interpersonal closeness (Sternberg, 2003), providing the ideal conditions for moral disengagement. Disgust, the rejection emotion, may be uniquely equipped to enable dehumanized social cognition.
Previous Research on Dehumanization
Much of the dehumanization (or infrahumanization) research has focused on the preferential attribution of humanizing characteristics, such as complex emotions (e.g., Leyens et al., 2000) or traits such as warmth and intelligence (e.g., Haslam et al., 2005), to ingroups relative to outgroups. Other research has examined implicit associations between outgroups and animals (e.g., Capozza, Boccato, Andrighetto, & Falvo, 2009; Goff, Eberhardt, Williams, & Jackson, 2008; Saminaden, Loughnan, & Haslam, 2010; Vaes, Paladino, & Puvia, 2011; Viki et al., 2006). Although these studies document a widespread dehumanizing bias in judgments of social and ethnic outgroups, it is not clear whether outgroup status per se (e.g., Tajfel, Billig, Bundy, & Flament, 1971) is sufficient for activating animalistic forms of dehumanization (but see Demoulin et al., 2009; Kofta, Baran, & Tarnowska, in press; Mirosławska & Kofta, 2007, for minimal-groups findings using emotion-focused infrahumanization measures).
With respect to the role of disgust in dehumanization, there is evidence that feelings of disgust may weaken or block perceptions of target humanity. In two social-neuroscience investigations, Harris and Fiske (2006, 2007) found that members of certain disgust-eliciting outgroups fail to be processed as fully human, social entities. Correlates of dispositional disgust sensitivity resemble these findings. Hodson and Costello (2007) reported positive correlations between interpersonal disgust sensitivity and a tendency to dehumanize immigrants with respect to “uniquely human” personality traits.
The above findings empirically document an association between disgust and dehumanization, however, the causal status of disgust in outgroup dehumanization has not been investigated experimentally. There is also no confirmation that directly animalistic dehumanization processes occur in the absence of prior attitudes, making the minimal conditions necessary for dehumanization unclear. The current research sought to address that gap in the literature by examining whether disgust can cause people to associate an arbitrary outgroup with animals.
Overview of the Current Study
This study examined the effect of disgust on implicit associations between outgroups and animals, as measured with a modified version of the Implicit Association Test (IAT; Greenwald, McGhee, & Schwartz, 1998) and using a minimal-groups paradigm (e.g., Tajfel et al., 1971). To test the disgust-dehumanization hypothesis, we primed feelings of disgust, sad, or neutral emotions. We predicted that incidental feelings of disgust would produce implicit dehumanizing biases, such that participants in the disgust condition would associate outgroup members with animality and ingroup members with humanity. In contrast, sad and neutral emotions were not expected to produce or augment dehumanizing biases.
Method
Participants
Participants were 102 undergraduate volunteers at a medium-sized Canadian university who participated in exchange for partial course credit. Approximately 80% were Canadian-born of European ancestry. Four participants were excluded for not following instructions during the emotion induction phase and one was excluded due to a preexisting hand injury. Data were excluded for three participants whose IAT scores exceeded two standard deviations from their group means (e.g., Cardinal & Aitken, 2006; Weinberg & Abramowitz, 2002) 1 . This left a final sample of 94 participants (70 female, 24 male; M age = 19.45, SD = 3.34).
Materials and Procedure
This study adapted the procedure used by DeSteno, Dasgupta, Bartlett, and Cajdric (2004) in their study of anger effects on IAT responses. Participants were tested in small groups of up to six people and all materials were presented by computer.
Minimal group assignment
Each session began with a bogus test asking participants to estimate the frequency of various neutral occurrences (e.g., “Number of pages in an average fiction novel?”). A results screen then indicated that the test sought to identify “overestimators” and “underestimators,” and displayed the participant’s test result, which was “overestimator” in every case. In this way, all participants were induced to regard overestimators as an ingroup and underestimators as an outgroup.
Group membership activation
Next, participants were shown photographs of their ingroup and outgroup members, that is, other “overestimators” and “underestimators.” A total of 12 color photographs (six ingroup, six outgroup) were selected from the Productive Aging Lab Face Database (Minear & Park, 2004), each depicting the frontal facial view of a Caucasian adult, aged 18–29, with equal gender representation. The backgrounds were color coded so that one group appeared before a grey backdrop and the other appeared before a white backdrop. A small caption explicitly indicated group membership. Group membership was counterbalanced so that the ingroup photographs were the outgroup for approximately half of participants. The photographs were presented one at a time in a random order, with ingroup members presented first, followed by outgroup members. This task was designed to activate group membership and to familiarize the participants with the stimuli used in the IAT portion of the study.
Dehumanization IAT and emotion induction
The stimuli used to capture the concepts of humanity and animality in the IAT task were 12 words used by Viki et al. (2006): person, humanity, people, man, woman, civilian, pet, mongrel, critter, creature, feral, and wildlife. We conducted a pilot study with 82 English-fluent U.S. participants (46.3% female, Mage = 31.93, SD = 11.26) recruited through Amazon’s Mechanical Turk Website (http://www.mturk.com) to ensure that the words both, (a) differed in their human/animal properties, and (b) were equated in valence. Participants rated the human and animal words (along with 10 filler items to disguise the purpose of the ratings; e.g., friend, glutton) on two 7-point scales: one that ranged from 1 (uniquely human) to 7 (uniquely animal), and a second scale that ranged from 1 (extremely negative) to 7 (extremely positive). These ratings were subjected to a multivariate analysis of variance (MANOVA) with word type as a within-subjects factor and English as a second language as a between-subjects factor. As expected, the human word ratings differed from the animal word ratings across the two measures, Wilks’ λ = .314, F(2, 79) = 86.43, p < .001, ηp2 = .69. English as a second language did not interact with the word ratings, F(2, 79) = 2.15, p = .12, ηp2 = .05. Most importantly, the human words (M = 1.16, SE = 0.35) differed significantly from the animal words (M = 6.18, SE = 0.25) on the uniquely human/uniquely animal dimension, F(1, 80) = 171.58, p < .001, ηp2 = .68. However, as expected, the human words (M = 4.57, SE = 0.35) did not differ from the animal words (M = 4.12, SE = 0.28) on the evaluative dimension, F(1, 80) = 1.24, p = .27, ηp2 = .02. Thus the word stimuli adequately capture dehumanization-relevant information without confounding evaluative meaning.
The dehumanization IAT included the standard set of seven IAT trial blocks. Depending on the particular block, participants categorized minimal ingroup and minimal outgroup photographs as “us” versus “them,” words as “human” versus “animal,” or simultaneously categorized both words and photographs according to their properties. The order of the combined test blocks was counterbalanced such that approximately half of the participants received the “Us + Human, Them + Animal” response key pairing first, and the other half received it second.
The initial emotion induction occurred immediately before the critical trials in the first IAT combined test block, where participants were randomly assigned to receive one of three emotion inductions: disgust, sadness, or neutral. The induction consisted of viewing three photographs from the International Affective Picture System (IAPS; Lang, Bradley, & Cuthbert, 2005), a widely used stimuli set that has been standardized according to its emotional content (Mikels et al., 2005). Similar procedures have been used by others to induce emotional states of disgust and sadness (e.g., Moretti & di Pellegrino, 2010). The disgust photographs portrayed an open-heart surgery, a dirty toilet, and a cockroach on a plate of food. The sadness photographs portrayed a young boy crying, a wounded soldier crying, and an elderly man at the bedside of his hospitalized wife. The neutral photographs portrayed everyday household objects, including a stool, a lamp, and a fan. IAPS codes were: 7380, 9320, 3250, 2900, 2205, 9421, 7020, 7025, 7175. We supplemented this pictorial emotion induction with an emotional writing task designed by DeSteno et al. (2004). Participants wrote for 4 minutes about an autobiographical event that made them feel very disgusted, very sad, or emotionally neutral, depending on condition. 2 To ensure that the induction lasted the duration of the IAT, participants continued writing about their recalled event for an additional 2 minutes before the second combined test block.
Data reduction followed the recommendations of Greenwald, Nosek, and Banaji (2003). All analyses were performed directly on the IAT D scores, which were coded so that positive values indicate an implicit dehumanizing bias (i.e., faster judgment of stimuli in trials with the “Us + Human, Them + Animal” response key pairing, relative to trials with the “Us + Animal, Them + Human” pairing).
Mood questionnaire
A mood questionnaire administered at the end of each session asked participants to rate their mood on a series of 5-point adjective rating scales, ranging from “very slightly or not at all” to “extremely.” Responses to three items, disgusted, repulsed, and revolted, were combined to create a composite disgust scale (α = .77). Responses to the single item sad provided the sadness rating.
Results
Manipulation Check: Emotion Inductions
Planned orthogonal comparisons were used to determine whether the emotion inductions were successful in producing incidental feelings of disgust and sadness. As expected, participants in the disgust induction condition reported more feelings of disgust (M = 1.65, SD = 0.82) than those in the sad (M = 1.16, SD = 0.32) and neutral (M = 1.19, SD = 0.48) conditions, t(91) = 3.74, p < .001, d = 0.78. In contrast, the sad and neutral conditions did not differ in their disgust ratings, t(91) = 0.22, p = .83, d = 0.05.
Similarly, as expected, participants in the sad condition reported more sad emotions (M = 2.06, SD = 1.22) than those in the disgust (M = 1.31, SD = 0.59) and neutral (M = 1.47, SD = 0.94) conditions, t(91) = 3.25, p = .002, d = 0.68. In comparison, the disgust and neutral conditions did not differ in their sadness ratings, t(91) = 0.64, p = .53, d = 0.13.
Dehumanization IAT Analyses
Descriptive statistics for the dehumanization IAT are presented in Table 1. Ninety-five percent confidence intervals (95% CIs) for the IAT D scores of each group were used to determine whether there was evidence of implicit dehumanizing biases. These results are also displayed in Table 1. As predicted, the D scores in the disgust condition were significantly greater than 0, indicating that disgusted participants were significantly faster in trials with the “Them + Animal” response key pairing than the “Them + Human” pairing. The D scores of the sad and neutral control conditions were also significantly greater than 0.
Dehumanization IAT response latencies and implicit bias scores by condition.
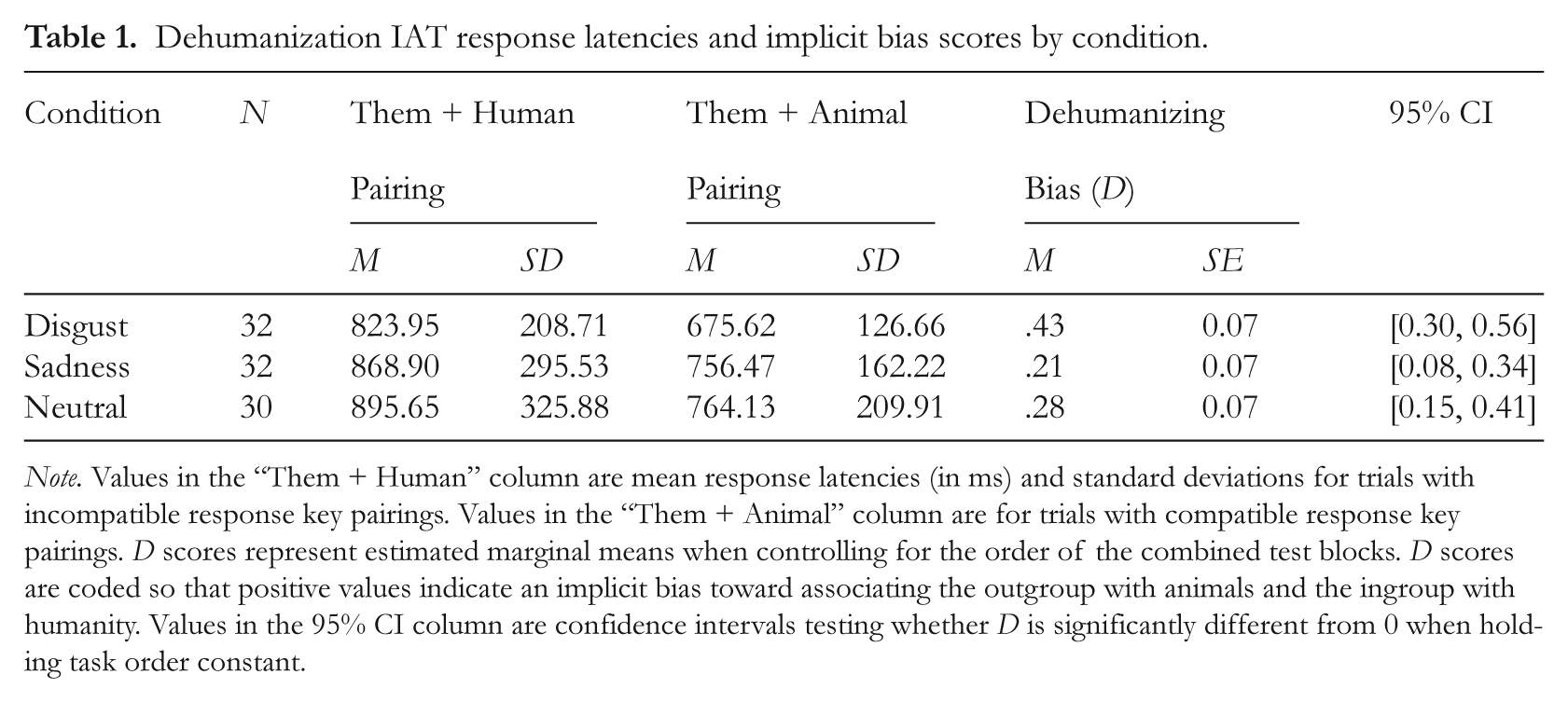
Note. Values in the “Them + Human” column are mean response latencies (in ms) and standard deviations for trials with incompatible response key pairings. Values in the “Them + Animal” column are for trials with compatible response key pairings. D scores represent estimated marginal means when controlling for the order of the combined test blocks. D scores are coded so that positive values indicate an implicit bias toward associating the outgroup with animals and the ingroup with humanity. Values in the 95% CI column are confidence intervals testing whether D is significantly different from 0 when holding task order constant.
Planned orthogonal comparisons tested whether participants in the disgust condition demonstrated greater dehumanizing biases than those in the sad and neutral conditions when controlling for the order of the combined test blocks. As expected, the IAT D scores were significantly greater in the disgust condition than in the sad and neutral control conditions, t(90) = 2.32, p = .023, d = 0.49. In contrast, the sad and neutral conditions did not differ in the magnitude of their dehumanizing biases, t(90) = 0.74, p = .46, d = 0.16. Figure 1 displays this pattern of means, with the strongest dehumanizing biases occurring in the disgust condition.
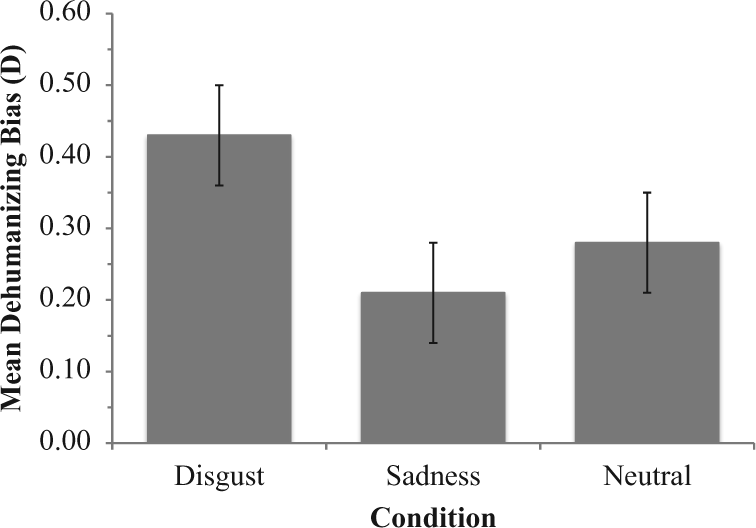
Mean IAT dehumanizing bias as a function of mood induction condition when holding block order constant.
Discussion
This study examined the influence of incidental disgust on outgroup dehumanization using a modified version of the Implicit Association Test (IAT; Greenwald et al., 1998) and a minimal-groups paradigm. In line with previous findings (e.g., Viki et al., 2006), all participants demonstrated a dehumanizing bias whereby outgroup members were more strongly associated with animals than were ingroup members. Feelings of disgust, however, significantly potentiated this dangerous cognitive bias. This effect was distinct from sadness, which showed little influence on dehumanization, suggesting that the observed effect of disgust on dehumanization is not due to negative valence priming. Our results therefore show both a general tendency to dehumanize members of arbitrary, minimal outgroups, and potentiation of that tendency via incidental disgust.
Although preliminary, these findings provide the first direct evidence that feelings of disgust can shift the human–animal boundary in social cognition, thus empirically linking it to the psychological machinery of outgroup dehumanization. Given that the mechanisms behind dehumanization have been relatively unexplored, we hope that our findings will encourage further investigations of this important topic. Deeper understanding of how disgust promotes dehumanization might lead to novel interventions for defusing intergroup violence (Hodson & Costello, 2007).
The current research is also one of the first to evidence dehumanizing associations using minimally created groups. Interestingly, although the dehumanizing biases were greatest in disgusted participants, all participants found it easier to associate outgroup members with animals, relative to ingroup members. This is notable because the minimal-groups paradigm ensured that they had no prior attitudes toward the group members. Interpersonal similarity, merely with respect to a previously unknown and unvalued tendency (i.e., “overestimation”), was sufficient to initiate an implicit dehumanization process. These results are consistent with those of Demoulin et al. (2009), who found evidence of explicit infrahumanization toward a minimal outgroup when the groups were formed by meaningful criteria. Thus in addition to documenting a causal role for disgust, this study adds implicit dehumanization to the array of intergroup biases that are activated by the most minimal—but meaningful—conditions of interpersonal similarity. Given that Demoulin et al. (2009) found that group meaningfulness was necessary for outgroup infrahumanization to emerge, it is possible that disgust works by imbuing greater meaningfulness to ingroup–outgroup distinctions (e.g., via the threat of disease). This emotional charging of group categorization may be a key component of the mechanism through which disgust influences dehumanization processes.
It is important to note that our findings are limited to IAT effects, which unfortunately do not distinguish between “outgroup dehumanizing” and “ingroup humanizing” biases. This is an inherent limitation of the IAT methodology. Since ingroup enhancement and outgroup derogation motives are not necessarily equivalent (Brewer, 1999), either or both may be involved in the effect found here. It is possible that disgust does not strengthen associations between outgroups and animals, but instead creates a stronger association between the ingroup and humanity. This would suggest that disgust reaffirms people’s unique humanity and extends it to similar others, at the expense of outgroup members. Additional studies using methods other than the IAT are necessary to resolve whether disgust promotes enhanced ingroup humanization, outgroup dehumanization, or both.
A second limitation is that the sadness and disgust manipulation check ratings were lower than expected, despite being significant in the appropriate directions and having large effect sizes. This was disappointing because the induction techniques have been successful at inducing moderately strong emotional states in previous research (e.g., Baumann & DeSteno, 2010; DeSteno et al., 2004; Moretti & di Pellegrino, 2010). We believe that the low ratings are likely due to a combination of two factors. First, an unfortunate choice of manipulation check scale anchors (1 corresponded to both “not at all” and “very little”) likely shifted scores to the low ends of the scales. Secondly, the ratings were completed at the end of the experiment when the effects of the emotion inductions were likely greatly diminished. These two factors may explain the low ratings. At the same time, it is also possible that the low ratings indicate that the manipulation failed to produce emotional states (i.e., a lack of “hot,” affective priming), but instead primed semantic associations involving disgust and sadness (“cold,” cognitive priming). 3 If this is correct, our findings carry a slightly different interpretation: exposure to concepts involving disgust facilitates dehumanizing associations in social-cognition without the recruitment of emotional architecture. Although the affective route seems more intuitively compelling, conceptual priming may be at least part of the story, if not the whole story, here. Unfortunately, it is often hard to disentangle these two processes, and even widely used implicit measures—such as the affective priming procedure (AMP; Payne, Cheng, Govorun, & Stewart, 2005)—have been subject to recent debate over the affective or conceptual nature of their effects (Blaison, Imhoff, Hühnel, Hess, & Banse, 2012). It would nonetheless be valuable to conduct a follow-up study with a more clearly conceptual manipulation (e.g., a sentence-unscrambling task) to examine whether semantic primes produce similar results.
It is also unclear whether the effects of disgust on dehumanizing associations are distinguishable from that of all other negative emotions. We evaluated sadness as a negative affect control, yet some might argue that a more stringent control would be anger or contempt. However, the effects of anger and contempt may prove very difficult to distinguish from that of disgust. All three are highly moralized other-condemning emotions (Haidt, 2003). Induced feelings of anger have also been shown to increase heuristic-based processing in social perception (Bodenhausen, Shepard, & Kramer, 1994), strengthen biases toward dispositional attributions of behavior (Keltner, Ellsworth, & Edwards, 1993), and cause implicit negativity toward outgroups (DeSteno et al., 2004). Consequently, we concede that anger and contempt may elicit dehumanization effects that are indistinguishable from that of disgust. On the other hand, anger and disgust are motivationally distinct. Anger is strongly associated with cognitive and behavioral approach (e.g., rumination, reactive aggression) whereas disgust is strongly associated with the opposite (Haidt, 2003). Indeed, dispositional disgust sensitivity is negatively associated with aggression for this very reason (Pond et al., 2012). The contexts that elicit anger and contempt are also clearly distinguishable from those that elicit disgust (Haidt, 2003; Hutcherson, & Gross, 2011). Therefore, if other condemning emotions like anger and contempt prove relevant to dehumanization processes, their differences from disgust may influence the form that dehumanized cognition takes. Anger-evoking contexts may entail more agentic forms of dehumanization, while disgust-evoking contexts may entail more self-protective, exclusionary forms of dehumanization. Future studies should examine whether anger and contempt can induce implicit dehumanization, and determine how and when their dehumanizing effects differ from those of disgust.
Follow-up studies could also employ other types of emotion manipulations; for example, the use of smells, movies, or emotional music. This would be particularly helpful for ruling out the possibility that disgust facilitated dehumanization more strongly than sadness simply because the disgust photographs contained less human stimuli than the sadness photographs (see the Method section for a description of the stimuli). The photo presentation was only a small part of the disgust manipulation—and the disgust writings largely involved human body products and people doing disgusting things—so this explanation may not be likely. Still, it is desirable to evaluate this and other possible alternative explanations for the effects seen here.
Conclusion
Tolerance and inclusive compassion are increasingly important in the ethnically and socially diverse communities of the 21st century. Accordingly, an important goal of psychological research is to identify social-cognitive biases that impede positive regard toward outgroups and the causal mechanisms that underlie those biases. The current research highlights a previously unarticulated effect of disgust on the most morally dangerous of intergroup biases: dehumanized perceptions of outgroup members. Further exploration of disgust’s function in shifting the human–animal boundary in social cognition will enrich our understanding of dehumanization and its all too commonly tragic role in intergroup violence.
Footnotes
Acknowledgements
The authors thank Mark Schaller for his comments on an earlier draft of this article.