
Abstract
Objective:
In this study, we investigated the effects of hydrogen sulfide (H2S) and sonic hedgehog (SHH) on the proliferation, autophagy, and apoptosis of human microvascular endothelial cells (HCMEC/D3). We also explored the regulatory relationship between cystathionine-β-synthase (CBS) and the SHH pathway.
Method:
Human microglia cells (HMC3) were stimulated under hypoxia to secrete H2S and SHH proteins, which were then co-cultured with HCMEC/D3 cells. The relationship between H2S and SHH was investigated by inhibiting the CBS or SHH pathways. Vascular endothelial growth factor (VEGF) levels in the culture medium were detected using ELISA, while H2S levels were detected via a micro method. The mRNA levels of hypoxia-inducible factor 1α (HIF-1 α), VEGF, Beclin-1, light chain 3 (LC3), and caspase-3 were detected using RT-PCR. Protein levels of CBS, SHH, extracellular regulated kinase 1/2 (ERK1/2), phospho-ERK1/2 (P-ERK1/2), Beclin-1, LC3, and Caspase-3 were determined by western blot.
Results:
The results indicated that HIF-1α expression increased in HMC3 under hypoxic conditions but did not change when either the SHH signaling pathway or CBS was suppressed. H2S secretion of HMC3 increased during hypoxia, with both CBS and SHH proteins being up-regulated. The inhibition of CBS resulted in decreased levels of H2S and SHH in HMC3. whereas SHH pathway is inhibited, the H2S secretion level is not affected. H2S and SHH proteins were found to enhance the expression of VEGF, P-EKR1/2, Beclin-1, and LC3, while decreasing the expression of caspase-3 in HCMEC/D3 cells.
Conclusion:
H2S secretion of HMC3 promotes the proliferation and regeneration of HCMEC/D3 by regulating SHH protein and alleviating hypoxic injury.
Introduction
Cardiovascular and cerebrovascular diseases (CCVDs) encompass pathological conditions affecting the vascular systems of the brain and heart. With changes in dietary patterns, there is a higher prevalence of CCVDs, and these conditions are affecting increasingly younger populations. Among these, ischemic heart disease and ischemic stroke are the leading causes of mortality globally, accounting for 16% and 11% of deaths, respectively. 1
When an ischemic cerebral infarction occurs, the surrounding cells experience hypoxia and may undergo necrosis. The primary therapeutic objective is to restore the transport of blood and nutrients.2,3 Development of collateral circulation plays a crucial role in delivering oxygen and nutrients to the infarcted tissue, thus facilitating recovery from ischemic stroke.4,5 This process involves the proliferation of vascular endothelial cells and the formation of new blood vessels, regulated by various factors such as blood pressure, protein molecules, cytokines, and gas molecules.6–9
Hydrogen sulfide (H2S), an endogenous gas signaling molecule, is frequently present in a variety of mammals and plays a significant role in both the cardiovascular and nervous systems.10–12 In mammals, H2S is primarily synthesized by cystathionine γ-lyase (CSE), CBS, 3-mercaptopyruvate sulfurtransferase (3-MST), and cysteine aminotransferase (CAT). 13 Within the central nervous system (CNS), l-cysteine is catalyzed by CBS, which is predominantly expressed in HMC3 microglia cells, to produce endogenous H2S. When brain tissue is deprived of oxygen, HMC3 is activated to secrete H2S.14,15 H2S reduces secondary injury induced by cerebral ischaemia-reperfusion through mechanisms such as anti-oxidative stress, suppression of inflammatory responses, inhibition of apoptosis, attenuation of cerebrovascular endothelial cell damage, and modulation of autophagy, thereby playing a vital role in other cerebral ischemic injury events.16,17 Research indicates that H2S can enhance the phosphorylation of protein kinase B (AKT) and ERK, elevate VEGF and ang1 levels, induce autophagy in damaged cells, and contribute to cellular homeostasis.18,19 The Hedgehog (Hh) family of secreted proteins plays an important role in cell growth, survival, and fate determination. SHH is the most widely expressed hedgehog protein. It has a critical role in the development of neural and vascular systems. When SHH binds to the transmembrane receptor patched-1 (PTCH1), it interrupts the inhibitory effect of PTCH1 on the transmembrane protein smoothened (SMO), leading to the activation of Gli transcription factors (Gli1-3), and the initiation of a downstream signaling cascade. 20 This process is known as the ‘canonical’ SHH signaling pathway. Recent research has also identified two distinct classes of ‘non-canonical’ SHH signaling pathways that operate independently of Gli-mediated transcription. 21
The SHH signal transduction cascade is crucial for the development of most tissues and organs, as well as for vasculogenesis. The SHH signaling pathway plays a significant role in the recovery from hypoxic brain injury, as it reduces protein oxidation, lipid peroxidation, and apoptosis induced by focal ischemia/reperfusion.22,23 Additionally, SHH promotes the establishment of collateral circulation vessels. 24 A recent study has demonstrated that application of the SHH protein enhances VEGF expression, improves microvascular density, and promotes angiogenesis. 25 Moreover, the SHH signaling pathway is involved in autophagy, contributes to cell homeostasis, and plays a significant role in cerebral ischemic injury.13,26–28
Although it has been proven that H2S and SHH can regulate the proliferation of vascular endothelial cells and autophagy, the influence between H2S and SHH remains unclear. We hypothesized that H2S promoted the proliferation and autophagy of hypoxic-injured vascular endothelial cells through SHH signaling pathway, while inhibiting apoptosis and alleviating hypoxic injury.
Materials and methods
Cell culture
HMC3 and HCMEC/D3 were purchased from Procell and Cellverse, respectively. Both cell types were cultured in DMEM (Gibco, Paisley, Scotland, UK) supplemented with 10% fetal bovine serum (Sigma, Uruguayan origin, S. Africa) and maintained at 37°C in a humidified atmosphere with 5% Carbon dioxide (CO2).
Co-culture and hypoxia
Human brain HCMEC/D3 and human HMC3 were inoculated into the upper and lower compartments of Transwell coculture systems (NEST, Wuxi, China) at a density of 2 × 105/mL. The experimental groups included a control group, a hypoxia group, a hypoxia + cyclopamine group, and a hypoxia + aminooxyacetic acid (AOAA) group. For hypoxic culture conditions, the cell culture plates were placed in an anaerobic tank (Mitsubishi Gas Chemical Company, Inc. Japan), along with an anaerobic production bag and an oxygen indicator. In hypoxic treatment, the indicator is pink, and the oxygen(O2) concentration in the anaerobic chamber is less than 0.1%. After reoxygenation, the culture medium was replaced with fresh medium, and cells were cultured for an additional 24 h. The hypoxia + cyclopamine group was cultured with fresh medium containing 10 μM cyclopamine (C860641, Macklin) for 24 h. The hypoxia + AOAA group was cultured with fresh medium containing 5 μM AOAA, an inhibitor of CBS activity through covalent binding and formation of an external aldimine with an oxime bond) (O805087, Macklin) for 24 h. To put it simply, 8 h hypoxia treatment was performed after co-culture, the medium was replaced after the end of hypoxia, and then routine culture was continued for 24 h to collect media and cells.
Cyclopamin inhibits SHH signaling pathway, and Cyclopamin inhibits SHH signaling pathway by targeting Smoothened (Smo) receptors, inhibiting SHH protein function. AOAA is a CBS inhibitor. AOAA covalently binds to CBS to form external aldehydamine with oxime bond, which inhibits CBS activity and thus inhibits H2S secretion.
H2S and VEGF content detection
H2S concentration in the cell media was detected using an H2S content detection kit (mlbio, Shanghai, China) after 24 h of reoxygenation. The measurements were conducted according to the manufacturer's instructions. The concentration of H2S (nmol/mL) was calculated using the formula:
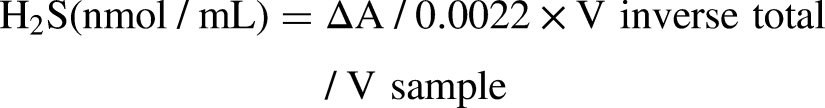
Angiogenesis assay
HMC3 media treated with anoxia for 8 h and inhibitor for 24 h were collected, and HCMEC/D3 in each group was re-suspended with these media. HCMEC/D3 were seeded at a density of 5 × 104 cells per well in a 48-well plate precoated with 50 µL of Matrix-Gel per well (Beyotime, Shanghai, China). Tube formation was quantified 4.5 h after various treatments by analyzing the sprouting tube-like structures in three randomly selected fields at 10 × magnification using an inverted phase-contrast microscope.
RT-PCR
Human HMC3 and HCMEC/D3 were collected after 24 h of reoxygenation for RT-PCR detection. RNAiso Plus (Takara, Beijing, China) was used to extract RNA from both cell types, and RNA concentrations were measured spectrophotometrically. Reverse transcription into cDNA was performed using 500 ng of isolated RNA with a reverse transcription kit (Takara, Beijing, China). VEGF, HIF-1α, Caspase 3, Beclin1, LC3, and glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA were detected using a LightCycler 96 (Roche, Basel, Switzerland).
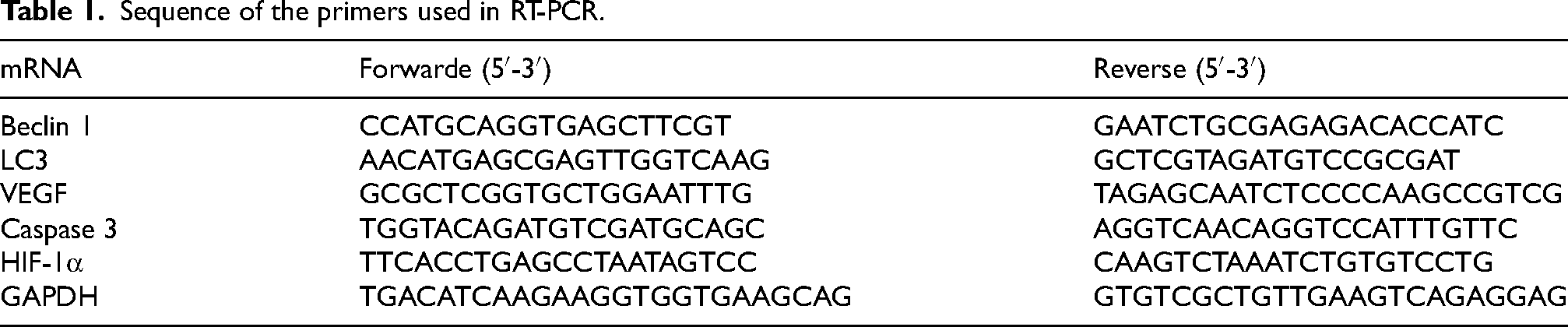
PCR reactions were performed in a 20-μL volume containing 10 μL of TB Green Premix Ex Taq II (Tli RNaseH Plus) (2X) (Takara, Beijing, China), 2 μL of template, 1 μL of specific primers (Table 1), and 6 μL of water. The thermal cycling profile included an initial denaturation step at 95 °C for 30 s, followed by 40 cycles of denaturation at 95 °C for 8 s, and annealing and extension at 60 °C for 30 s. The specificity of primers and amplification was demonstrated by subsequent analysis of melting points, which revealed single peaks for each target gene. Quantitative results were expressed using the Ct method, where the number of cycles (Ct) at which the fluorescence signal exceeded a defined threshold for GAPDH was subtracted from the Ct values of the target genes (ΔCt = Ct target - Ct GAPDH). Relative expression levels were determined as 2−ΔΔCt.
Sequence of the primers used in RT-PCR.
Western blot analysis
The RIPA Lysis Buffer (Beyotime, Shanghai, China) was used to separate proteins from cells, and their concentrations were determined using the BCA Protein Assay Kit (Beyotime, Shanghai, China). Protein samples (2 μg/μL) from boiled lysates were subjected to Sodium dodecyl sulfate(SDS) polyacrylamide gel electrophoresis (20 μg/lane) and blotted on hydrophobic immobilon-P Polyvinylidene fluoride(PVDF) transfer membranes (Millipore Corporation, Billerica, MA, USA). To prevent unspecific binding, membranes were blocked with protein-free rapid blocking buffer (Yamei, Shanghai, China) for 120 min at room temperature. Subsequently, membranes were washed five times with TBS containing 0.1% Tween 20 (TBS-T), each wash lasting 5 min, followed by overnight incubation with primary antibodies. Mouse Caspase 3/p17/p19 Monoclonal antibody (66470-2-Ig), Mouse Beclin 1 Monoclonal antibody (66665-1-Ig), Rabbit LC3 polyclonal antibody (14600-1-AP), Rabbit SHH Polyclonal antibody (20697-1-AP), Mouse CBS Monoclonal antibody (67861-1-Ig) Rabbit ERK1/2 polyclonal antibody (11257-1-AP); Rabbit Phospho-ERK1/2 (Thr202/Tyr204) polyclonal antibody (28733-1-AP) (Proteintech Group, Wuhan, China); and Rabbit anti-glyceraldehyde-3-phosphate dehydrogenase(GAPDH) (bs-2188R) (Bioss, Beijing, China). Primary antibodies were diluted 1:1000, with the exception of the anti-GAPDH antibody, which was diluted 1:2000. Dilutions were performed with tris buffered saline Tween 20 (TBS-T)TBS-T containing 5% milk powder. Subsequently, membranes were washed five times (each time for 5 minutes) with TBS containing 0.1% Tween 20 (TBS-T). Later, membranes were also washed with TBS-T (5 times, each time for 5 minutes), followed by incubation with anti-rabbit IgG (A0208) (Proteintech Group, Wuhan, China) or anti-mouse ImmunoglobulinG(IgG) (SA00001-1) (Proteintech Group, Wuhan, China) diluted 1:20000 in TBS-T (containing 5% milk powder). Membranes were washed again five times with TBS-T for 5 min each. Finally, blots were developed using enhanced chemiluminescence (ECL) with the Omni-ECL™ ultra-sensitive chemiluminescence test kit (YaMei, Shanghai, China), and Hyperfilm was exposed in a gel imager (Thermo Fisher, Waltham, Massachusetts, USA). Imaging results are analyzed using Image J (National Institutes of Health, Bethesda, Maryland, USA) and referred to GAPDH in corresponding samples.
Data and statistical analyses
Statistical analyses were conducted using GraphPad Prism 9. For datasets involving RT-PCR, Western blotting, H2S concentration, and ELISA, a one-way ANOVA with multiple comparisons test was employed. P < 0.05 was considered statistically significant.
Results
Hypoxia stimulates an increase in H2S secretion by HMC3
To investigate the stimulatory effect of hypoxia on H2S secretion by HMC3 cells, we treated HMC3 cells to varying durations of hypoxic conditions and subsequently measured the H2S concentration in the culture medium. The results demonstrated a significant increase in H2S content in the medium after 8 h of hypoxia compared to baseline (P < 0.01) (Figure 1).
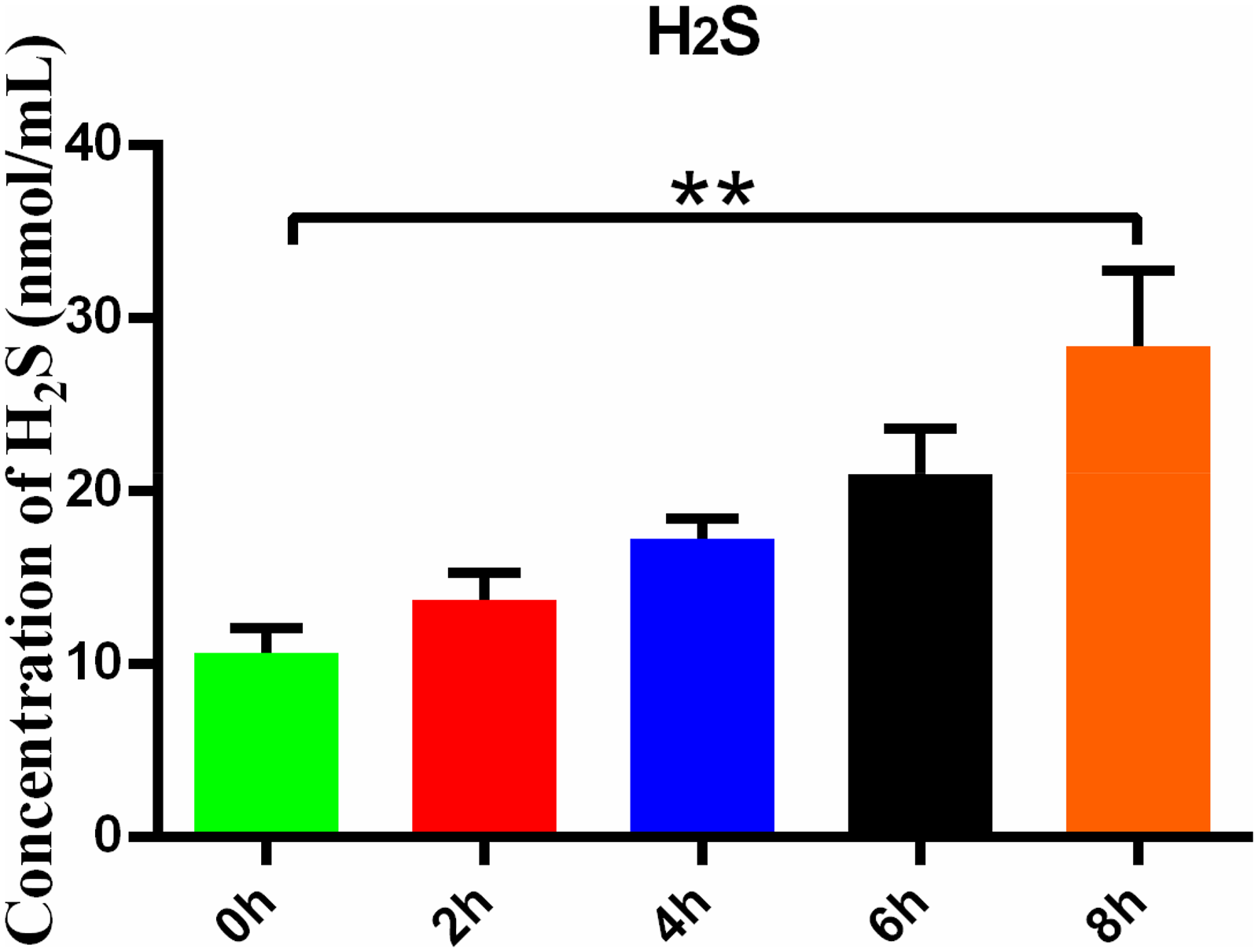
Hypoxia stimulates the increase of H2S secretion by HMC3. H2S content in HMC3 medium was treated with different hypoxia durations. Data are represented as mean ± SD. **P < 0.01, compared with the 0-h group.
Hypoxia activates HIF-1α and promotes H2S secretion
Our findings revealed that hypoxia significantly upregulated the expression of HIF-1α mRNA in HMC3 cells compared with the control group (P < 0.05). However, there were no significant differences in HIF-1α mRNA expression levels between the hypoxia group and the groups treated with hypoxia + cyclopamine group (P > 0.05). Meanwhile, there was no significant difference in the expression of HIF-1α mRNA between hypoxia + AOAA group and hypoxia group (P > 0.05) (Figure 2A). Additionally, the H2S content in the medium of HMC3 cells exposed to hypoxia was significantly higher compared to the control group (P < 0.005). The H2S content in hypoxia + cyclopamine group was small decreased, but there was no significant change in H2S content between the hypoxia + cyclopamine group and the hypoxia alone group (P > 0.05). Compared with the hypoxia group, the H2S content in the hypoxia + AOAA group was significantly reduced (P < 0.01) (Figure 2B). These results indicate that hypoxia-induced activation of HIF-1α in HMC3 cells. Hypoxia promotes H2S secretion in HMC3 cells, and inhibition of CBS reduces H2S secretion by HMC3 cells under hypoxic conditions.
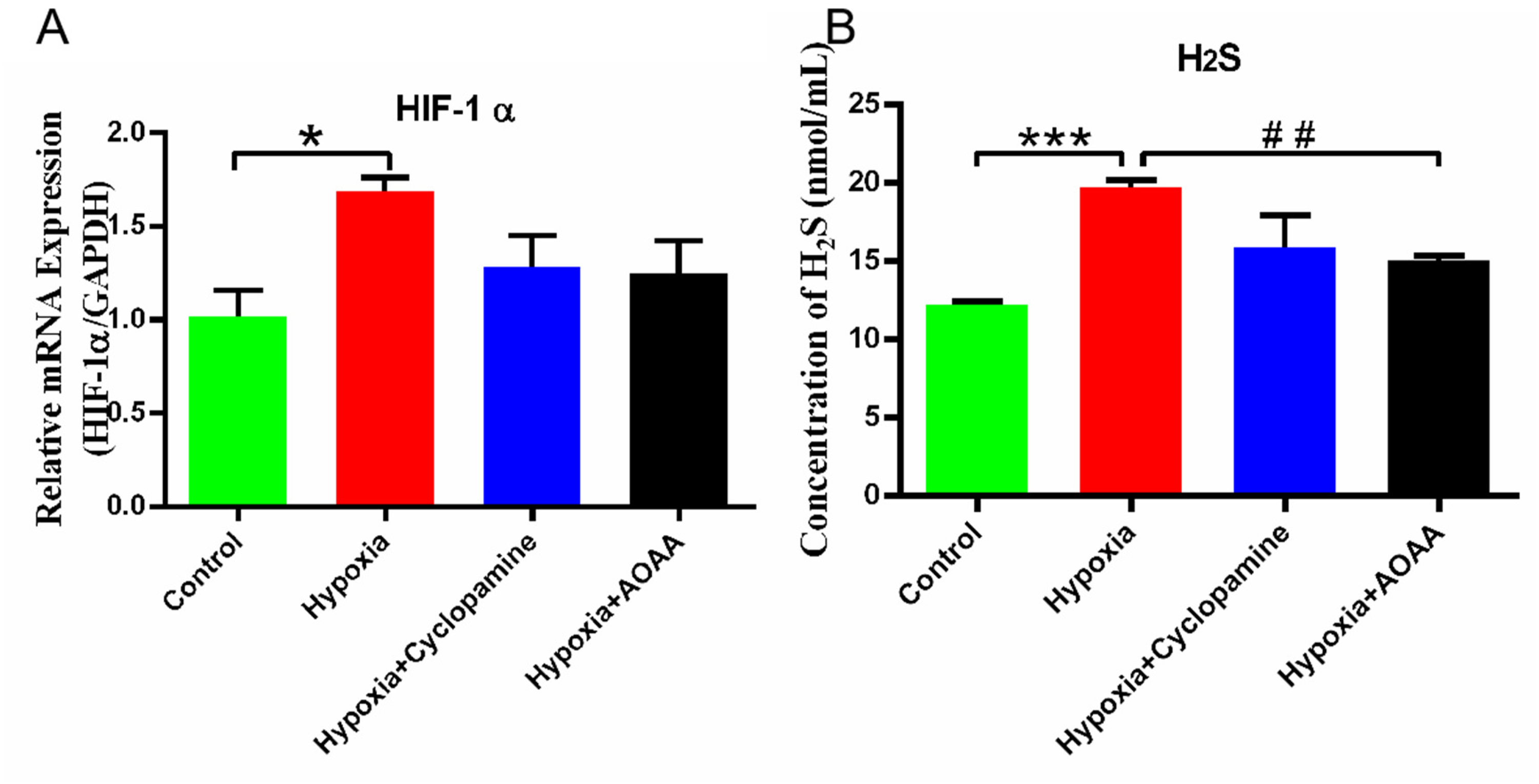
Effects of hypoxic stimulation on HMC3. (A) HIF-1α mRNA expression in HMC3. (B) H2S content in medium. Data are represented as mean ± SD. *P < 0.05. ***P < 0.005, compared with the control group. ##P < 0.01, compared with the hypoxia group.
H2S levels affect SHH secretion
Our findings confirm that CBS regulates the secretion of H2S by HMC3. In addition, we investigated the impact of H2S levels on the SHH signaling pathway. By treating HMC3 with hypoxia, we explored the relationship between the SHH pathway and H2S synthesis by suppressing either the CBS or SHH signaling pathway.
Compared with the control group, the expression of CBS protein in control + cyclopamine group and control + AOAA group had no significant change (P > 0.05), but the expression of CBS protein in hypoxia group was significantly increased (P < 0.01). Compared with hypoxia group, the expression of CBS protein in hypoxia + cyclopamine group had no significant change (P > 0.05). However, CBS protein levels significantly decreased in the hypoxia + AOAA group compared to the hypoxia group (P < 0.01). Similarly, compared with the control group, the expression of SHH protein in control + cyclopamine group and control + AOAA group had no significant change (P > 0.05), but the expression of SHH protein in hypoxia group was significantly increased (P < 0.05). There was no significant change in SHH protein level in the hypoxia + cyclopamine group compared to the hypoxia group (P > 0.05). Conversely, the level of SHH protein in the hypoxia + AOAA group decreased significantly, compared with the hypoxia group (P < 0.05) (Figure 3). Our findings reveal that under hypoxic conditions, inhibition of CBS leads to decreased protein levels of both CBS and SHH. This suggests that CBS in HMC3 may impact SHH protein synthesis in HMC3 by modulating H2S levels, ultimately affecting the SHH signaling pathway. Conversely, when hypoxia occurred and the SHH signaling pathway was inhibited, the level of CBS protein remained unchanged, suggesting that SHH did not impact CBS protein synthesis.
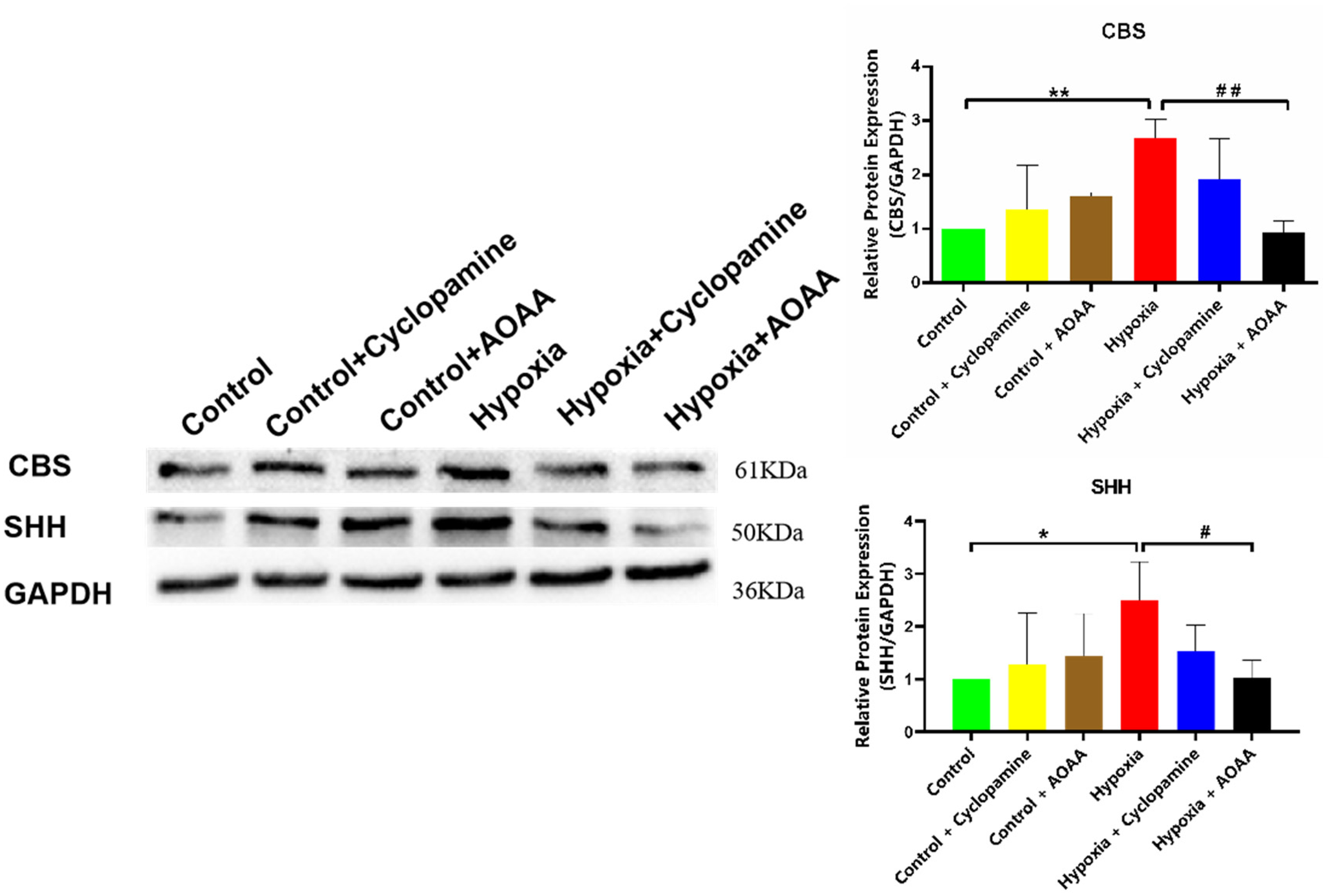
Inhibition of CBS or SHH signaling pathways on CBS and SHH proteins. Data are represented as mean ± SD. *P < 0.05, **P < 0.01,compared with the control group.
H2S promotes tube formation in HCMEC/D3 via SHH signaling pathway
In order to investigate the effect of H2S and SHH pathway on blood vessel formation in HCMEC/D3, blood vessel formation experiments were conducted. The results of mesh formation number showed that compared with the control group, the number of mesh per mm2 in hypoxia group was significantly increased (P < 0.005). Compared with hypoxia group, the number of mesh per mm2 in hypoxia + Cyclopamine group was significantly reduced (P < 0.01). Compared with the hypoxia group, the number of mesh per mm2 in the hypoxia + AOAA group was significantly decreased (P < 0.01)(Figure 4B). The results showed that compared with the control group, the length of the main trunk in the hypoxia group was significantly increased (P < 0.005). Compared with hypoxia group, the main length of hypoxia + Cyclopamine group was significantly reduced (P < 0.01). Compared with the hypoxia group, the length of the main trunk in the hypoxia + AOAA group was significantly decreased (P < 0.01) (Figure 4C). The results showed that compared with the control group, the branch length of hypoxia group was significantly increased (P < 0.005). Compared with hypoxia group, the branch length of hypoxia + Cyclopamine group was significantly reduced (P < 0.01). Compared with hypoxia group, the branch length of hypoxia + AOAA group was significantly decreased (P < 0.01) (Figure 4D). These results indicate that H2S promotes the proliferation and migration of HCMEC/D3 cells to form blood vessels through SHH signaling pathway.
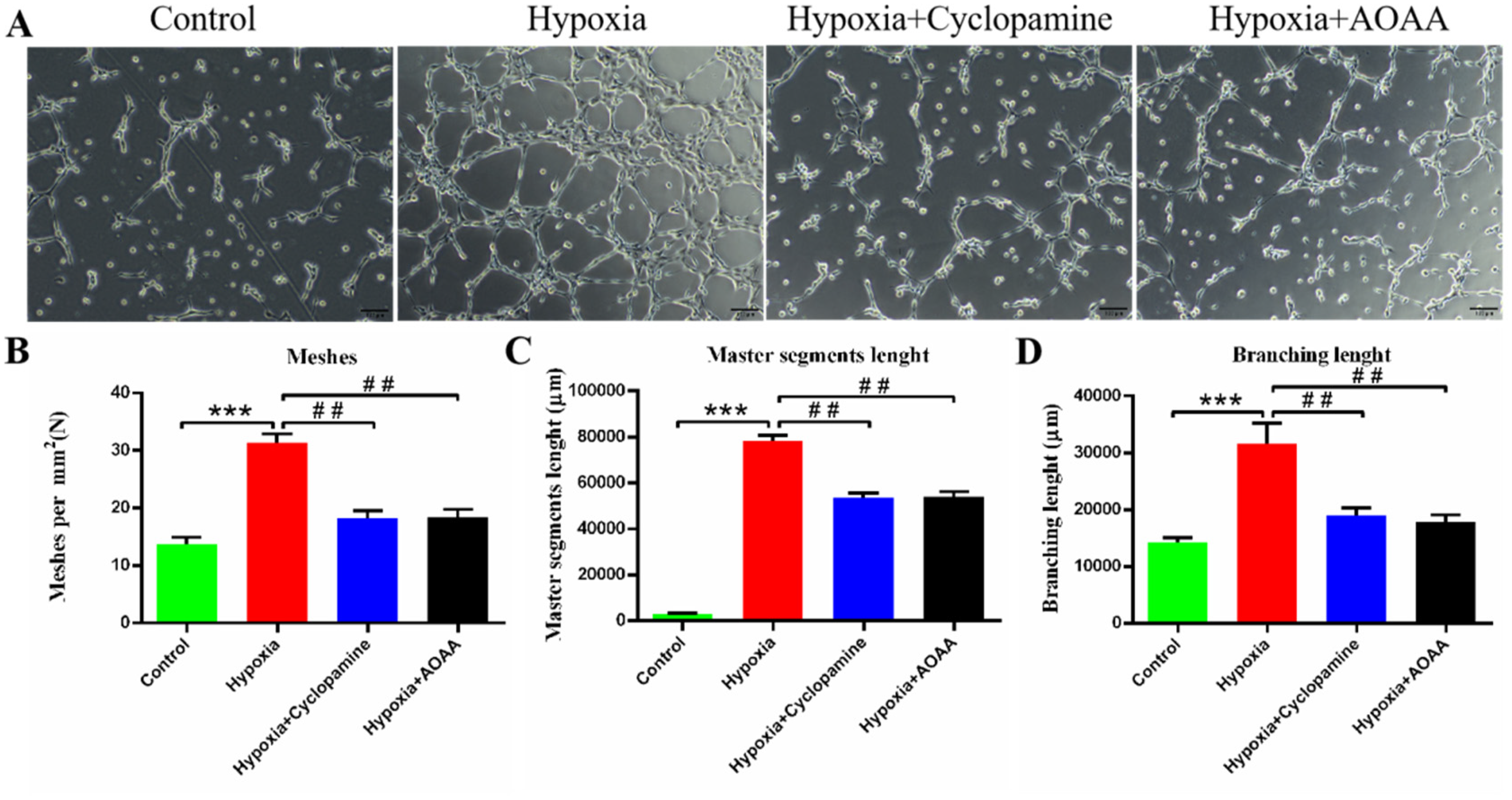
Effects of H2S and SHH signaling pathways on HCMEC/D3 angiogenesis. (A) 4.5 h HCMEC/D3 cells were cultured in each group for angiogenesis experiment. (B) The number of blood vessel grids formed per mm2. (C) Trunk Length. (D) Branch Length. Data are represented as mean ± SD. ***P < 0.005, compared with the control group. ## P < 0.01, compared with the hypoxia group.
H2S and SHH proteins promote VEGF secretion and ERK phosphorylation in HCMEC/D3
The previous results demonstrated that hypoxia leads to increased secretion of H2S and SHH proteins in HMC3 cells. To investigate how H2S affects the proliferation of human HCMEC/D3 cells via SHH signaling pathway, we conducted a co-culture of these two cell types to observe changes in human microvascular cells.VEGF and ERK expression levels were detected in HCMEC/D3. When comparing the content of VEGF in the medium, we discovered that the VEGF levels in the hypoxia group were significantly higher than those in the control group (P < 0.01). Compared with the hypoxia group, VEGF levels in hypoxia + cyclopamine group and hypoxia + AOAA group were decreased in medium (P < 0.05, P < 0.005). (Figure 5A). Compared with the control group, VEGF mRNA in the hypoxia group was significantly increased (P < 0.05). At the same time, VEGF mRNA levels in the hypoxia + cyclopamine group were significantly decreased compared to the hypoxic group (P < 0.05). VEGF mRNA levels were significantly decreased in the hypoxia + AOAA group compared to the hypoxic group (P < 0.05) (Figure 5B).
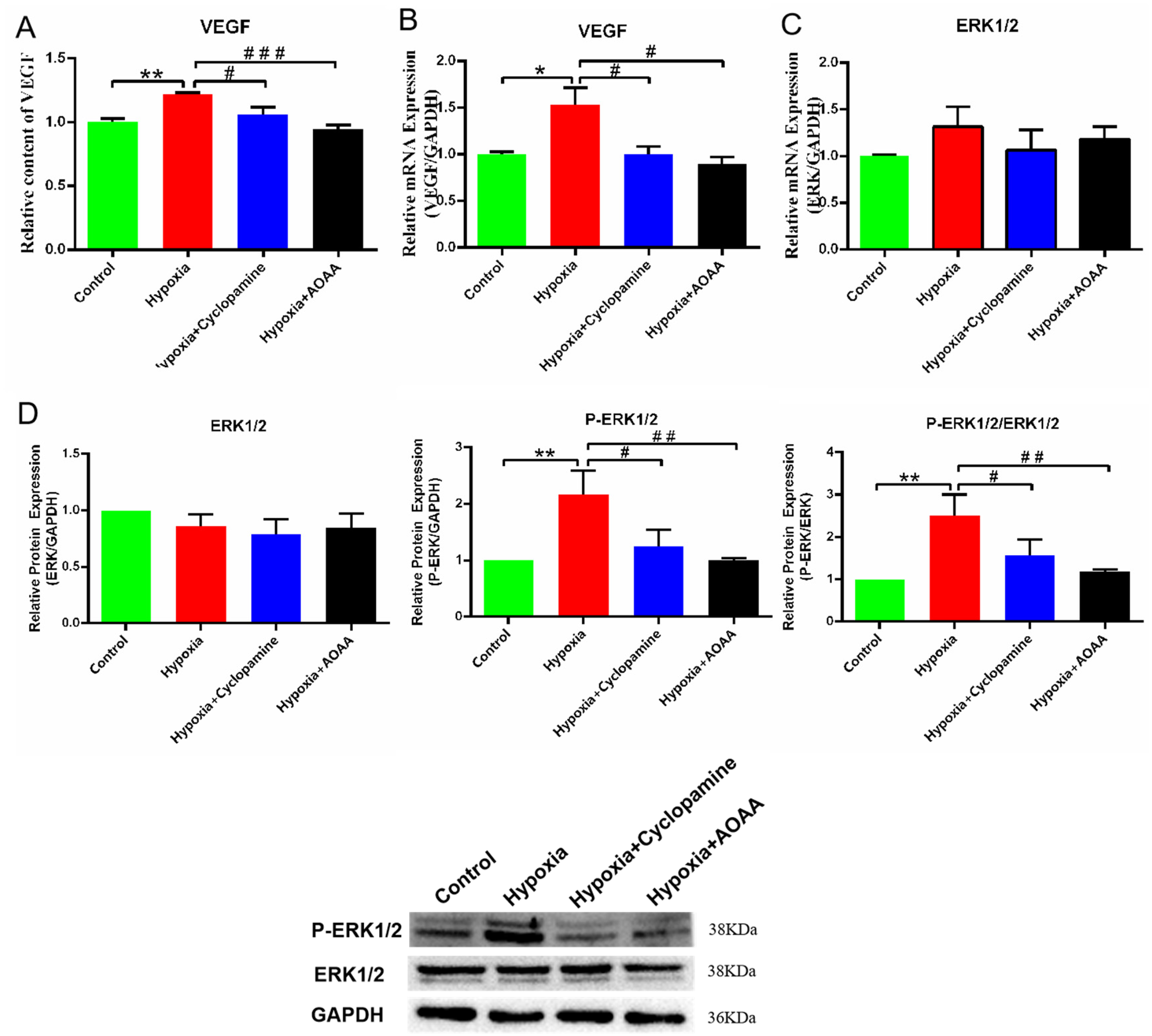
Expression level of VEGF and ERK in HCMEC/D3. (A) VEGF levels in medium. (B) VEGF mRNA level. (C) ERK1/2 mRNA level. (D) Levels of phospho-ERK1/2 and ERK1/2 by Western blot analyses. Data are represented as mean ± SD. *P < 0.05. **P < 0.01, compared with the control group. # P < 0.05. ## P < 0.01. ### P < 0.005, compared with the hypoxia group.
According to the results of the mRNA and protein analysis, there was no significant change in ERK1/2 levels across all groups (P > 0.05) (Figure 5C、5D). However, the ratio of P-ERK1/2 to ERK1/2 was significantly increased in the hypoxia group compared to the control group (P < 0.01). Compared with the hypoxia group, the ratio of P-ERK1/2 to ERK1/2 in both the hypoxia + cyclopamine group (P < 0.05) and the hypoxia+ AOAA group (P < 0.01) decreased (Figure 5D). These results indicate that H2S enhance VEGF secretion and ERK1/2 phosphorylation through SHH signaling pathway, thereby promoting the proliferation of HCMEC/D3 cells.
H2S promotes autophagy and alleviates apoptosis by regulating the SHH pathway
Subsequently, we investigated the effects of H2S and SHH proteins on autophagy apoptosis in hypoxic-damaged HCMEC/D3 cells. Autophagy and apoptosis-related genes were detected in the co-cultured HCMEC/D3 cells. In this study, RT-PCR results revealed the Beclin-1 mRNA level in the hypoxia group was significantly increased compared to the control group (P < 0.05). However, Beclin-1 mRNA levels in the hypoxia + cyclopamine group (P < 0.01) and the hypoxia + AOAA group (P < 0.01) were significantly lower than those in the hypoxia group. Meanwhile, the changes in LC3 mRNA in each group were consistent with Beclin-1 mRNA.
Additionally, undifferentiated in the level of caspase- 3 transcription in the hypoxic group compared to the control group (P > 0.05). Compared with the hypoxia group, the hypoxia + AOAA group had a increase (P < 0.01) (Figure 6A). Western blot results showed that Beclin-1 protein levels were significantly increased in HCMEC/D3 in the hypoxia group compared to the control group (P < 0.05). Interestingly, the Beclin-1 protein levels were significantly reduced in the hypoxia + cyclopamine and hypoxia + AOAA groups compared to the hypoxia group (P < 0.01, P < 0.05). Furthermore, the LC3II/LC3I protein in HCMEC/D3 was significantly up-regulated in the hypoxia group compared to the control group (P < 0.01). Compared to the hypoxia group, the LC3II/LC3I protein in HCMEC/D3 in both the hypoxia + AOAA group and the hypoxia + cyclopamide group showed a significant reduction (P < 0.01,P < 0.01). Caspase- 3 protein level in HCMEC/D3 did not significantly differ between the hypoxia and control groups. However, the caspase- 3 protein level of HCMEC/D3 in the hypoxia + AOAA group was significantly up-regulated compared to the hypoxia group (P < 0.01) (Figure 6B). These results indicate that H2S secreted by HMC3 cells can promote autophagy in hypoxia-damaged HCMEC/D3 cells via SHH signaling pathway, facilitating the removal of damaged organelles and mitigating cellular injury. Simultaneously, they can alleviate the apoptosis in HCMEC/D3 cells caused by hypoxia, thereby preventing extensive cell death and reducing hypoxia-induced damage.
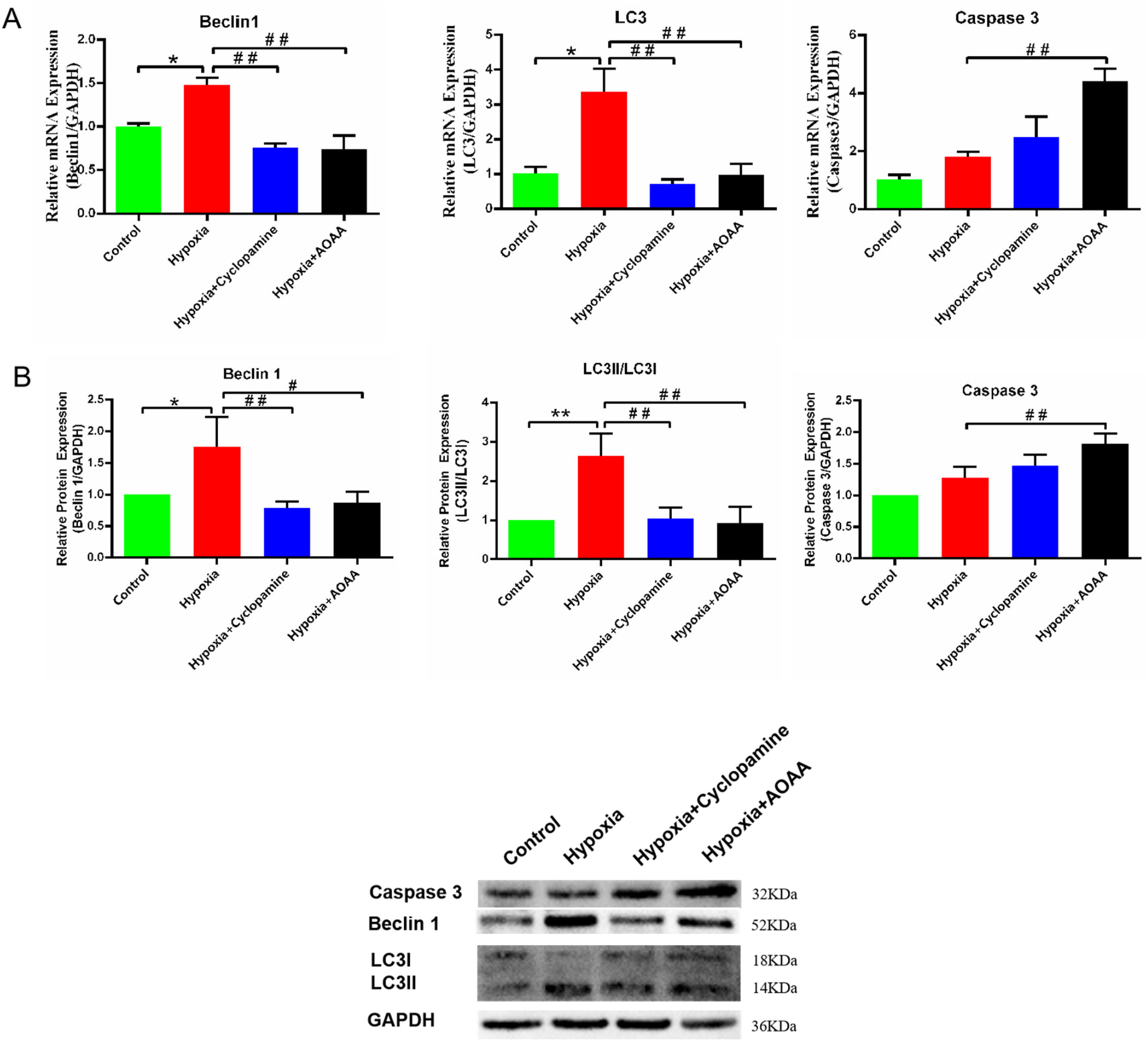
Effects of H2S and SHH passage on autophagy and apoptosis of HCMEC/D3. (A) Changes in Caspase- 3, Beclin-1, and LC3 mRNA levels in HCMEC/D3. (B) Changes in Caspase −3, Beclin-1, and LC3 protein levels in HCMEC/D3. Data are represented as mean ± SD. *P < 0.05. **P < 0.01, compared with the control group. # P < 0.05. ## P < 0.01, compared with the hypoxia group.
Discussion
Ischemic stroke has long posed a significant threat to the quality of human life and overall survival. The formation of collateral circulation is a fundamental prerequisite for the restoration of blood flow following an ischemic stroke. This process is intimately linked to the proliferation and regeneration of HCMEC/D3 cells. Recent research has identified several factors influencing angiogenesis, including angiogenic factors like VEGF and basic fibroblast growth factor (bFGF), gas transmitters such as H2S and carbon monoxide(CO), and signaling pathways like the SHH pathway, mechanistic target of rapamycin (mTOR) signaling pathway, and mitogen-activated protein kinases (MAPK) signaling pathway.29–31
The synthesis of H2S is mainly controlled by three enzymes: CSE, CBS and 3MST, among which CBS is mainly produced in brain tissue. CBS is mainly expressed by astrocytes, microglia and neurons, and astrocytes and neurons mainly perform the function of signal transmission in brain tissue,while microglia are a kind of immune cells associated with anti-inflammatory hypoxic repair. 32 Studies have shown that under hypoxic stimulation, microglia will react quickly to be activated, migrate and prolifically. More microglia will produce more CBS, which is conducive to H2S synthesis and accelerate the recovery of brain tissue function after hypoxia.33,34 Therefore, microglia were selected as the donor cells of endogenous H2S in this study.
Research has demonstrated that H2S can increase the mRNA and protein levels of HIF-1α and VEGF in brain capillary endothelial cells, thereby enhancing the binding activity of HIF-1α to promote angiogenesis under hypoxic conditions. 13 In cerebral ischemic diseases, H2S mitigates ischemia by promoting the phosphorylation of AKT and ERK and increasing the expression of VEGF and angiopoietin-1 (Ang-1). Similarly, the SHH signaling pathway plays a crucial role in neovascularization, angiogenesis, and carcinogenesis in adults. Studies have shown that SHH acts as an indirect angiogenic agent by upregulating angiogenic growth factors, including VEGF and the angiopoietins Ang-1 and Ang-2. 35 In addition, the SHH pathway can promote the phosphorylation of ERK1/2, thereby facilitating cell proliferation and regeneration. 36 Upregulation of VEGF levels and the phosphorylation of ERK1/2 are critical for promoting vascular endothelial cell proliferation and new blood vessel formation. In this context, the H2S and SHH pathways mostly play significant regulatory roles. In this study, when H2S secretion increases, the length of the vascular network and branches formed by human brain microvascular endothelial cells increases. Conversely, when H2S is inhibited, the length of the vascular network and branches formed by human brain microvascular endothelial cells also decreases.Our study demonstrates that increased H2S levels correlate with elevated VEGF and ERK1/2 levels, enhancing vascular endothelial cell proliferation and angiogenesis. Conversely, inhibition of H2S synthesis or the SHH pathway results in decreased VEGF and ERK1/2 levels, thus impeding the proliferation of vascular endothelial cells.
The H2S and SHH pathways are crucial not only for angiogenesis but also for regulating autophagy and apoptosis in disease pathogenesis. In cardiovascular disease research, H2S has been shown to decrease atherosclerotic formation and plaque vulnerability by promoting autophagy of vascular smooth muscle cells (VSMC), promoting collagen secretion, and inhibiting apoptosis. 18 Additionally, some scholars have proved that H2S can mitigate cell damage by suppressing apoptosis and promoting autophagy. 37 Moreover, low concentrations of H2S protect brain tissue by inhibiting apoptosis and regulating autophagy after cerebral ischemia reperfusion. 38 Similarly, the SHH signaling pathway promotes the production of autophagosomes in hypoxic-damaged cells and maintains cell homeostasis under hypoxic stimulation.39–41 Researchers have investigated the role of inhibiting SHH in promoting apoptosis during the development of cancer. 42 also have proven that the SHH signaling pathway can regulate cell apoptosis both in vivo and in vitro, 43 these findings align with the present study, where increased levels of H2S and SHH promote autophagy and suppress apoptosis in hypoxic-damaged vascular endothelial cells. Conversely, inhibiting H2S synthesis or the SHH pathway results in opposing effects. In conclusion, the H2S and SHH pathways facilitate autophagy, suppress apoptosis, and uphold cell homeostasis during hypoxic injury.
Some researchers have demonstrated that H2S can regulate the SHH signaling pathway to inhibit cancer development in cancer therapy. 44 However, whether H2S can play a promoting role in the process of angiogenesis by regulating the SHH signaling pathway remains unproven. In this study, we investigated the effects of H2S on the proliferation, autophagy, and apoptosis of hypoxic-injured HCMEC/D3 cells by regulating the SHH pathway. Initially, the concentration of H2S in the HMC3 medium was measured under varying durations of hypoxia. Subsequently, the impact of H2S and SHH secreted by HMC3 on the proliferation, autophagy, and apoptosis of HCMEC/D3 cells was explored through co-culture experiments. The results indicate that H2S influences the secretion of SHH protein and regulates VEGF levels via the SHH pathway, thereby affecting the regeneration and proliferation of HCMEC/D3 cells. Additionally, H2S and SHH contribute to cell proliferation by regulating ERK phosphorylation. H2S and SHH alleviate hypoxic damage by promoting autophagy and inhibiting apoptosis in HCMEC/D3 cells. Overall, H2S regulates hypoxia-induced cell regeneration and cell homeostasis in HCMEC/D3 cells by controlling the SHH signaling pathway.
In this study, we investigated the relationship between CBS inhibition and SHH pathway inhibition to determine any potential interactions. The results demonstrated that H2S production decreased and SHH protein content decreased when CBS was inhibited. Conversely, inhibition of the SHH pathway had no significant effect on CBS protein expression. These findings suggest that H2S synthesized by CBS could affect SHH protein expression and the function of the SHH signaling pathway. H2S was found to promote HCMEC/D3 proliferation and regeneration by regulating the SHH pathway and alleviating hypoxic injury. However, there are some limitations to this study and more in-depth research is needed to elucidate the precise mechanism by which H2S affects SHH protein expression.
Footnotes
Abbreviations
Authors’ contributions
Conception and design of the research: Hai-yan Chen, Xue-bin Li, Jie Wang.
Acquisition of data: Pin Zheng, Gui-xin Yang, Bing-bing Qin, Meng-Xue Zang.
Analysis and interpretation of the data: Jian-min Huang, Pin Zheng, Gui-xin Yang.
Statistical analysis:Jian-min Huang, Bing-bing Qin, Meng-Xue Zang.
Obtaining financing: Xue-bin Li, Hai-yan Chen.
Writing of the manuscript: Hai-yan Chen, Xue-bin Li, Jie Wang.
Critical revision of the manuscript for intellectual content: Hai-yan Chen, Xue-bin Li, Jie Wang.
All authors read and approved the final draft.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by a grant from National Natural Science Foundation of China (No. 81860226); Guangxi Medical and Health Appropriate Technology Development and Application Project (No. S2018073); Young and Middle-aged Backbone Talent Scientifc Research Projects of the Affiliated Hospital of Youjiang Medical University for Nationalities in 2021(No. Y20212610); Guangxi Health Commission Self -Financed Research Project (No. Z-L20230898).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Availability of data and materials
The datasets generated and/or analysed during the current study are not publicly available but are available from the corresponding author (Xue-bin Li) on reasonable request.