
Abstract
Background:
Genetic variations in a common single nucleotide polymorphism in the ninth intron of the KIBRA gene have been linked to memory performance and risk of Alzheimer's disease (AD).
Objective:
We examined the risk of AD related to presence of KIBRA T allele (versus CC homozygote) and to memory performance. The role of established genetic risk factors APOE ε4 and BDNF Met was also considered.
Methods:
Participants were cognitively healthy individuals (n = 19), participants with amnestic mild cognitive impairment (aMCI) due to AD (n = 99) and AD dementia (n = 37) from the Czech Brain Aging Study. Binary and multinomial logistic regressions compared odds of belonging to a certain diagnostic category and multivariate linear regressions assessed associations with memory.
Results:
KIBRA T allele was associated with increased AD dementia risk (odds ratio [OR] = 5.98, p = 0.012) compared to KIBRA CC genotype. In APOE ε4 negative individuals, KIBRA T allele was associated with a greater risk of both aMCI due to AD (OR = 6.68, p = 0.038) and AD dementia (OR = 15.75, p = 0.009). In BDNF Met positive individuals, the KIBRA T allele was associated with a greater risk of AD dementia (OR = 10.98, p = 0.050). In AD dementia, the association between KIBRA T allele and better memory performance approached significance (β = 0.42; p = 0.062). The link between possessing the KIBRA T allele and better memory reached statistical significance only among BDNF Met carriers (β = 1.21, p = 0.027).
Conclusions:
Findings suggest that KIBRA T allele may not fully protect against AD dementia but could potentially delay progression of post-diagnosis cognitive deficits.
Introduction
In the early 2010s, a genome-wide association study identified KIBRA as a potential candidate gene associated with encoding and retrieval of episodic memory. 1 KIBRA (also known as WWC1 for WW-and-C2-domain-containing-protein-1), first described in 2003, is a cytoplasmic protein that is highly expressed in the KIdney and the BRAin. 2
KIBRA is expressed in memory-related brain regions such as hippocampal CA1 and CA2, dentate gyrus, and neocortex, as well as in the cerebellum and hypothalamus.3,4 In addition to being involved in multiple cellular functions such as cell migration, vesicular transport, transcription, synaptogenesis, and neuronal signaling, 5 KIBRA protein interacts with other proteins participating in synaptic plasticity and memory formation, including dendrin and synaptopodin,3,6 as well as protein kinase C, zeta (PKCζ). 7
It has been shown that a common single nucleotide polymorphism (SNP rs17070145, T → C exchange) in the ninth intron of the KIBRA gene influences episodic memory and modulates hippocampal activation during memory acquisition. In 2006, Papassotiropoulos and colleagues 3 reported on a link between the KIBRA SNP and episodic memory in Swiss healthy young adults, which they replicated in two additional cohorts from the United States and from Switzerland. 3 Carriers of the rs17070145 T allele (CT or TT genotypes) showed better delayed recall and less hippocampal activation/effort during memory retrieval using functional magnetic resonance imaging (fMRI) compared to T allele non-carriers. 3 The association between the KIBRA rs17070145 T allele and better memory has been confirmed in several other studies and cohorts, including, for example, those with 64 healthy elderly individuals, 8 312 older adults without dementia, of whom 136 met the study criteria for the diagnosis of amnestic mild cognitive impairment (aMCI), 9 2230 nondemented participants, 10 and 383 healthy subjects. 11 However, the connection between the KIBRA rs17070145 T allele and delayed memory was not confirmed in other studies, 12 and yet other studies report associations between better episodic memory performance in rs17070145 T allele non-carriers. 13 Finally, a meta-analysis reported a significant association between SNP rs17070145 T allele and better performance on both episodic and working memory. 14
In addition, KIBRA may interplay with other genetic correlates of cognitive performance, particularly episodic memory, like the Apolipoprotein E ε4 allele (APOE ε4) 15 and the brain derived neurotropic factor (BDNF) Met allele. 16
In addition, the effect of the KIBRA SNP rs17070145 on Alzheimer's disease (AD) risk has also been investigated, with similarly inconclusive results as studies with memory as the outcome. For example, in a study by Rodríguez-Rodríguez and colleagues, 17 the T allele of KIBRA was found to be significantly associated with an increased risk of late-onset AD, and at least one other study confirmed this association. 18 However, other studies came to the opposite conclusion, such that KIBRA T allele non-carriers possess a greater risk of AD than KIBRA T allele carriers.19,20
Building on prior research, our aim was to examine having KIBRA T allele versus being a KIBRA CC homozygote in relation to the risk of AD, whether in the form of aMCI due to AD or AD dementia, and in relation to performance on memory cognitive tasks. We also tested whether any observed associations may be moderated by the presence of at least one APOE ε4 or the BDNF Met allele.
Methods
Participants
One hundred fifty-five eligible participants were selected from the Czech Brain Aging Study (CBAS), a longitudinal, memory clinic-based study on aging and cognitive impairment. 21 All patients were referred to the clinic by general practitioners, neurologists, or geriatricians based on memory complaints reported by themselves, their relatives, or health professionals. Of these, all eligible participants diagnosed with aMCI due to AD or with AD dementia were included in our analyses. The control group consists of cognitively healthy volunteers who are tertiary university (University of the 3rd Age) students, relatives of patients or of employees, and other volunteers who did not express significant cognitive complaints and scored within the normal range on a standard neuropsychological battery. In addition to standard clinical and laboratory assessments, all participants also underwent KIBRA, APOE, and BDNF genotyping, global neuropsychological assessment, and MRI within three months from the initial visit.
Of the 155 eligible participants, 19 were cognitively healthy individuals, 99 were diagnosed with aMCI due to AD with a high or intermediate probability of AD etiology, 22 and 37 with AD dementia with evidence of the pathophysiologic process of AD. 23 Patients with aMCI due to AD and AD dementia underwent a spinal tap with CSF collection. Commercial ELISA kits (Innogenetics) were used for dementia biomarker analyses (Aβ42, total tau [t-tau], and phospho-tau [p-tau] 181), with cut-off values derived from validation study.21,24 Patients with aMCI due to AD and AD dementia had positive CSF AD biomarkers (reduced Aβ42 and elevated p-tau 181). Cognitively healthy individuals were defined as subjects whose cognitive performance was within the normal range (i.e., not more than 1.5 standard deviations below the age- and education-adjusted norm on any cognitive test). 21 The diagnosis of the aMCI stage was based on Petersen's criteria for aMCI, 25 with memory complaints reported by the patient or caregiver, evidence of memory impairment on neuropsychological tests (i.e., score less than 1.5 standard deviations below the age- and education-adjusted norms in any memory test), as well as generally intact activities of daily living, and no evidence of dementia. 25 In this study, only patients with aMCI—both aMCI single-domain (isolated memory impairment) and aMCI multiple-domain (memory impairment plus impairment of at least one other cognitive domain) phenotypes—were included. Dementia etiology was diagnosed according to established guidelines, using the Diagnostic and Statistical Manual of Mental Disorders (DSM-V) criteria.26,27
All participants included in the study signed an informed consent approved by the Motol University Hospital ethics committee.
Exclusion criteria
The study excluded participants with pre-existing neurological or psychiatric conditions that could impair cognitive function, such as major depressive symptoms, defined as >8 points on the 15-item Geriatric Depression Scale, stroke, tumor, traumatic brain injury, or multiple sclerosis. 21
KIBRA, BDNF, and APOE genotyping
KIBRA, BDNF, and APOE genotypes were determined at the Department of Clinical Biochemistry, Hematology, and Immunology, Homolka Hospital, Prague, Czech Republic. DNA isolation was performed by Zybio Nucleic extraction kit WB-B from whole blood samples according to manufacturer's protocol (Zybio, Chongqing, China).
The genotyping of KIBRA rs17070145 and BDNF rs6265 single nucleotide polymorphism (SNP) were done by high-resolution melting (HRM) analyses. The reactions were performed with LightScanner Master mix (BioFire Diagnostics, SLC., USA) according to manufacturer's PCR reaction conditions with forward and reverse primers: KIBRA conditions: KibfF: 5´- ATA ACC TTT CAT CTA TTC TG-3´; Kibr 5´-GGA ATC TCT TGA CCC AGT AT-3´; Temperature profile was: 95°C for 2 min followed by 40 cycles of 95°C/30 s; 60°C/30 s; 72°C/30 s; Melting 60°C-90°C. BDNF conditions: BDNFf: 5´- AAA GAA GCA ACA TCG AGG ACA A-3´; BDNFr: 5´-ATT CCT CCA GCA GAA AGA GAA GAG-3´; Temperature profile was: 95°C for 2 min followed by 40 cycles of 95°C/30 s; 62°C/30 s; 72°C/30 s; Melting 60°C-90°C. HRM analysis was performed on LightScanner (IdahoTech).
APOE genotyping was performed according to IdahoTech protocol (Luna Probes Genotyping Apolipoprotein [ApoE] Multiplexed Assay) for HRM analysis.
Neuropsychological assessment
A comprehensive neuropsychological examination of all patients included in the CBAS includes the Mini-Mental State Examination (MMSE) as a screening of global cognitive function, 28 and the following tests to assess cognitive domains: 1) attention and working memory by Digit Span Forward (F-DigitSpan-SC) and backward (B-DigitSpan-SC), adaptation from the Uniform Data Set (UDS-cz 2.0) 29 and Trail Making Test (TMT) A, 30 2) memory by the immediate (LOG-I) and 20-min delayed (LOG-D) recall of the Logical Memory, adaptation from the Uniform Data Set (UDS-cz 2.0) 31 and Rey-Osterrieth Complex Figure test, reproduction after 3 min (ROCF-R), 32 3) language by the Boston naming test, 30 odd-items version (BNT-30) 32 and semantic verbal fluency - animals (S-VF-A),33,34 4) executive function by TMT B 30 and phonemic verbal fluency (P-VF; Czech version with letters N-K-P), 33 and 5) visuospatial function by ROCF copy condition (ROCF-C) 31 and the Clock Drawing Test (CDT). 35
Based on the literature, the neuropsychological battery included in this study was composed of the MMSE, 28 and the following tests to assess memory cognitive domain: LOG-I and LOG-D recall of the Logical Memory, adaptation from the Uniform Data Set (UDS-cz 2.0), 29 and ROCF-R. 31
Statistical analysis
All statistical analyzes were adjusted for sex (male/female), years of education and age, measured as age at assessment of controls and as estimated age of onset of aMCI due to AD or AD dementia.
Initially, between-group differences in age, years of education, and memory cognitive domain scores were evaluated using the independent samples t-test for differences in means and the Pearson chi-square test for between-group differences in sex and genotypes.
Prior to the main analyses, composite scores for the memory cognitive domain were calculated by standardizing the raw scores for each neuropsychological test to z-scores using the mean and standard deviation for each subgroup and then averaging to create individual composite scores for memory cognitive domain.
In main analyses, we first estimated logistic regression models to assess whether the risk of cognitive impairment, first overall and then aMCI due to AD or AD dementia specifically, would vary between KIBRA T allele carriers versus non-carriers (i.e., KIBRA CC homozygotes). Unadjusted models were estimated initially, models adjusted for age, sex, and education later. First, we estimated a binary logistic model contrasting cognitive impairment against no impairment. Then, we conducted a multinomial logistic regression in which aMCI due to AD and AD dementia were entered as separate levels of the dependent variable, with participants without cognitive impairment again serving as the reference category. Then, we performed multivariate linear regression models adjusted for age, sex, and education to examine the association between KIBRA T allele and memory performance. Finally, we stratified the sample by APOE ε4 or BDNF Met allele positivity and estimated separate adjusted multinomial logistic regression models to test whether APOE or BDNF variants yield different results than those observed with the overall sample. Achieved statistical power was assessed using the G*Power software. 36
Results
Demographic characteristics
Group demographic and clinical characteristics are reported in Table 1. Compared to cognitively healthy individuals, participants with aMCI due to AD were significantly older (p < 0.001), scored significantly worse on MMSE, LOG-I, LOG-D, ROCF-R, and S-VF-A (p < 0.001), TMT-A (p = 0.016), P-VF (p = 0.002), BNT-30 (p = 0.002), CDT (p = 0.005), and ROCF-C (p = 0.009). There were also significant sex differences between control subjects and patients with aMCI due to AD (p = 0.048). Patients with AD dementia were significantly older (p < 0.001), scored significantly worse on MMSE, LOG-I, LOG-D, ROCF-R, B-DigitSpan-SC, TMT-B, BNT-30, S-VF-A, and CDT (p < 0.001), TMT-A (p = 0.004), P-VF (p = 0.001), and ROCF-C (p = 0.001) compared to cognitively healthy individuals. There were no significant differences in KIBRA CC, CT, and TT allele distributions between controls and patients with aMCI due to AD, but the CC allele appeared overrepresented among patients with AD dementia compared to controls. The genotype distributions for the KIBRA SNP rs17070145 were in the Hardy-Weinberg equilibrium for controls, patients with aMCI due to AD, as well as patients with AD dementia.
Demographic features and cognitive performance of total dataset, control subjects, patients with aMCI due to AD, and AD dementia.
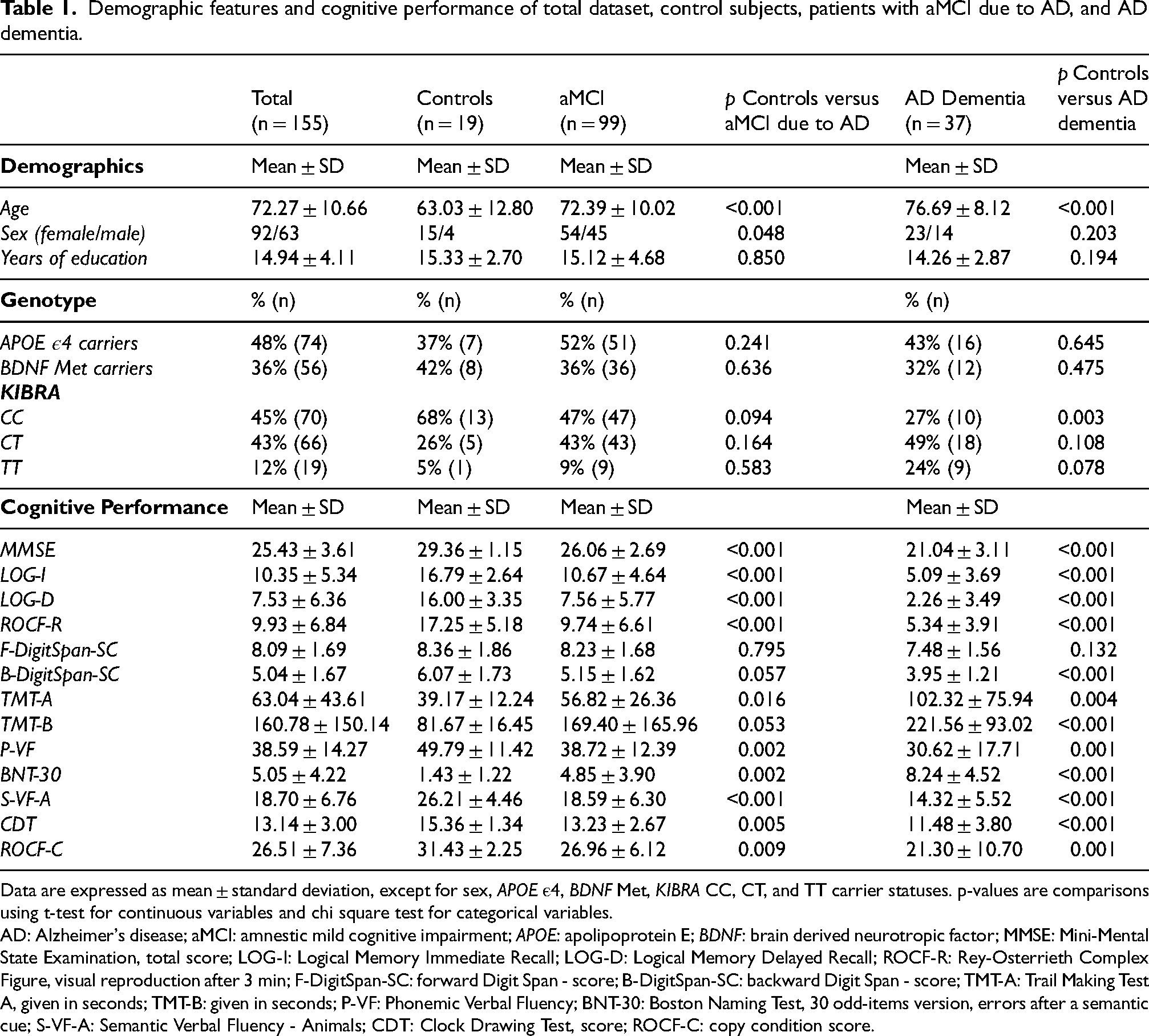
Data are expressed as mean ± standard deviation, except for sex, APOE ε4, BDNF Met, KIBRA CC, CT, and TT carrier statuses. p-values are comparisons using t-test for continuous variables and chi square test for categorical variables.
AD: Alzheimer's disease; aMCI: amnestic mild cognitive impairment; APOE: apolipoprotein E; BDNF: brain derived neurotropic factor; MMSE: Mini-Mental State Examination, total score; LOG-I: Logical Memory Immediate Recall; LOG-D: Logical Memory Delayed Recall; ROCF-R: Rey-Osterrieth Complex Figure, visual reproduction after 3 min; F-DigitSpan-SC: forward Digit Span - score; B-DigitSpan-SC: backward Digit Span - score; TMT-A: Trail Making Test A, given in seconds; TMT-B: given in seconds; P-VF: Phonemic Verbal Fluency; BNT-30: Boston Naming Test, 30 odd-items version, errors after a semantic cue; S-VF-A: Semantic Verbal Fluency - Animals; CDT: Clock Drawing Test, score; ROCF-C: copy condition score.
Examine whether risk of AD, whether in the form of aMCI due to AD or AD dementia, varies between KIBRA T allele carriers versus non-carriers
When comparing participants with AD (aMCI due to AD and AD dementia together) to controls, an unadjusted binary logistic model indicated that having the KIBRA T allele was associated with an increased risk of impairment compared to not having the T allele (odds ratio [OR] = 3.04, 95% CI 1.09–8.48, p = 0.034). This association was reduced to non-significant when adjusted for age, sex, and years of education (OR = 3.16, 95% CI 0.94–10.65, p = 0.063) (see Table 2).
Association between AD in the form of aMCI due to AD or AD dementia and KIBRA T allele.
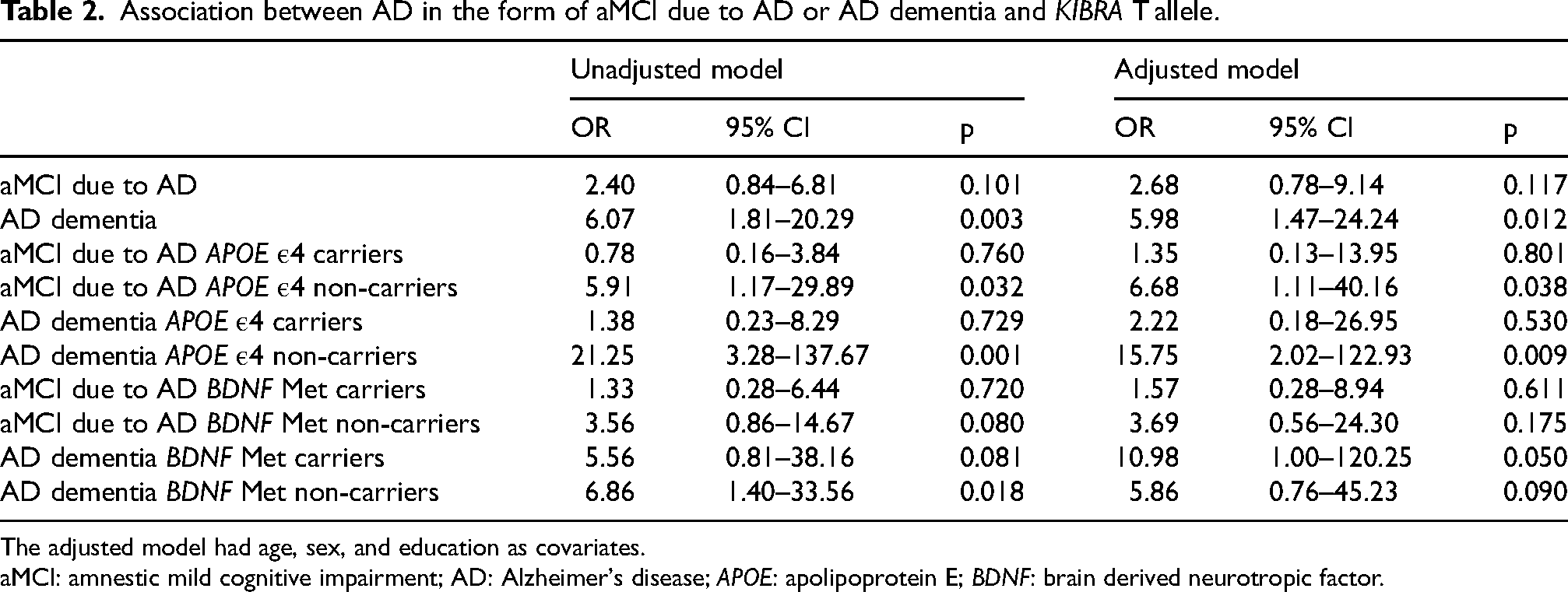
The adjusted model had age, sex, and education as covariates.
aMCI: amnestic mild cognitive impairment; AD: Alzheimer's disease; APOE: apolipoprotein E; BDNF: brain derived neurotropic factor.
When comparing patients with aMCI due to AD and AD dementia individually against controls, an unadjusted multinomial logistic regression model indicated that the KIBRA T allele increased the risk of aMCI due to AD non-significantly (OR = 2.40, 95% CI 0.84–6.81, p = 0.101) and of AD dementia significantly (OR = 6.07, 95% CI 1.81–20.29, p = 0.003). The results were similar after adjustment for age, sex, and years of education (OR = 2.68, 95% CI 0.78–9.14, p = 0.117 for aMCI due to AD and OR = 5.98, 95% CI 1.47–24.24, p = 0.012 for AD dementia).
Examine whether risk of AD (aMCI due to AD and AD dementia) related to having the KIBRA T allele may be different for APOE ε4 or BDNF Met allele carriers
We used the adjusted multinomial logistic regression models to examine the KIBRA T allele in relation to the diagnostic groups in APOE ε4 positive and negative individuals separately. In the APOE ε4 positive individuals, the presence of the KIBRA T allele did not show statistically significant associations with either diagnosis aMCI due to AD (OR = 1.35, 95% CI 0.13–13.95, p = 0.801) or AD dementia (OR = 2.22, 95% CI 0.18–26.95, p = 0.530). In the APOE ε4 negative individuals, having the KIBRA T allele was related to greater risk of both aMCI due to AD (OR = 6.68, 95% CI 1.11–40.16, p = 0.038) and AD dementia (OR = 15.75, 95% CI 2.02–122.93, p = 0.009). The risk of the combined aMCI due to AD and AD dementia group associated with the presence of KIBRA T allele was also significant in this subgroup (OR = 5.77, 95% CI 1.09–30.49, p = 0.032).
In the same adjusted models stratified by BDNF Met positivity, the presence of the KIBRA T allele showed greater odds of AD dementia (OR = 10.98, 95% CI 1.00–120.25, p = 0.050) but not of aMCI due to AD (OR = 1.57, 95% CI 0.28–8.94, p = 0.611) in BDNF Met positive participants. Having the KIBRA T allele was not significantly associated with either diagnosis aMCI due to AD (OR = 3.69, 95% CI 0.56–24.30, p = 0.175) or AD dementia (OR = 5.86, 95% CI 0.76–45.23, p = 0.090) in BDNF Met negative participants.
Examine whether KIBRA T allele carriers perform differently on memory tasks than KIBRA T non-carriers
In multivariate linear regression models adjusted for age, sex, and education, we found no association between the presence of the KIBRA T allele and better memory performance (β = −0.02, p = 0.886), which was similarly non-significant in APOE ε4 carriers (β = 0.16, p = 0.493) and non-carriers (β = −0.24, p = 0.314)
When the sample was stratified based on clinical diagnosis, we found a non-significant negative association between the KIBRA T allele and memory performance among the controls (β = −0.52, p = 0.087) and no association among patients with aMCI due to AD (β = 0.04, p = 0.808).
Among patients with AD dementia (see Figure 1), the association between the presence of KIBRA T allele and better memory performance approached significance (β = 0.42, p = 0.062). Therefore, we proceeded to examine this association further, finding that the association between having the KIBRA T allele and better memory was observably stronger in APOE ε4 carriers (β = 0.63, p = 0.240) than non-carriers (β = 0.24, p = 0.422), albeit non-significant in either subsample. This same association was statistically significant among BDNF Met carriers (β = 1.21, p = 0.027) but was essentially null in BDNF non-carriers (β = −0.11, p = 0.604).
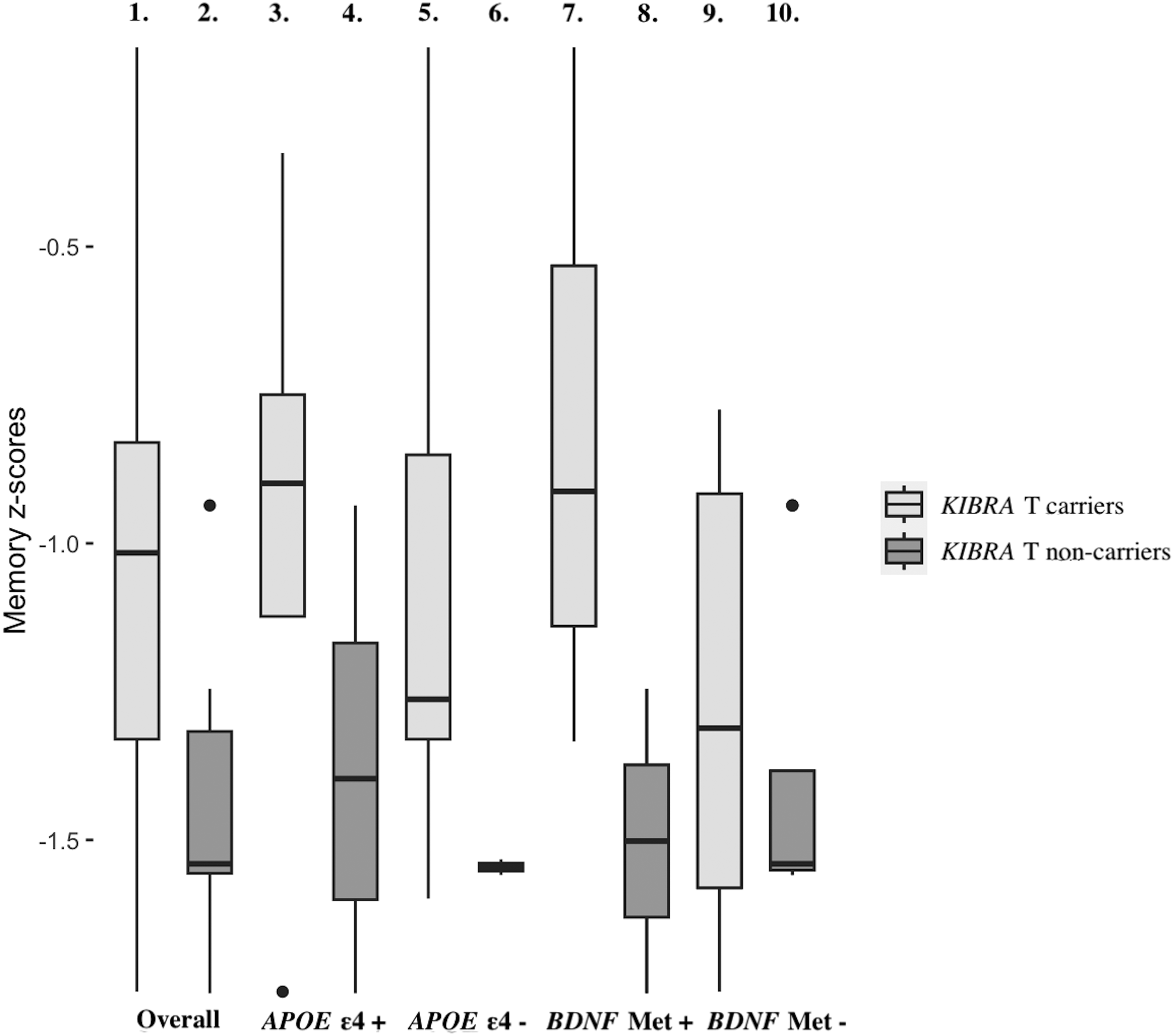
Association between memory performance and diagnosis of AD dementia within KIBRA T allele carriers and non-carriers. The boxplots represent the distribution and median of memory performance, as indicated by memory z-scores, within each group defined by patient characteristics such as KIBRA T allele carriers/non-carriers and APOE ε4 and BDNF Met status. The first box represents overall AD dementia KIBRA T allele carriers, the second overall AD dementia KIBRA T non-carriers, the third AD dementia KIBRA T and APOE ε4 carriers, the fourth AD dementia APOE ε4 carriers without KIBRA T allele, the fifth AD dementia KIBRA T carriers without APOE ε4, the sixth AD dementia KIBRA T and APOE ε4 non-carriers, the seventh AD dementia KIBRA T and BDNF Met carriers, the eighth AD dementia BDNF Met carriers without the KIBRA T allele, the ninth AD dementia KIBRA T carriers without BDNF Met, and the tenth AD dementia KIBRA T and BDNF Met non-carriers.
Discussion
We aimed to investigate the association between KIBRA T allele, AD-related cognitive impairment in the form of aMCI due to AD or AD dementia, and memory performance. First, we assessed whether the KIBRA T allele may vary between participants with versus without cognitive impairment, then proceeding to assess whether KIBRA T allele varies across in patients with aMCI due to AD or AD dementia compared to controls, and whether the presence of at least one APOE ε4 or BDNF Met allele plays a role in any observed associations. Second, we explored whether KIBRA T allele carriers performed differently on memory tasks than non-carriers, and whether any observed links between KIBRA T allele and memory could be affected by the presence of at least one APOE ε4 or BDNF Met allele.
We report two main findings. First, in the unadjusted model, we observed a significant association between carrying the KIBRA T allele (as opposed to KIBRA CC) and an increased risk of aMCI due to AD and AD dementia. Although this association was reduced to non-significant (p = 0.063) in the fully adjusted model in this overall sample, it was significant even after adjustment for covariate among those who were APOE ε4 negative (p = 0.032). Second, we found that having the KIBRA T allele was related to better memory performance among patients with AD dementia, although this result did not reach statistical significance (p = 0.062). The association was observably stronger in KIBRA T allele carriers who were also APOE ε4 positive compared to KIBRA T non-carriers. The association was significantly stronger in KIBRA T allele carriers who were also BDNF Met positive compared to KIBRA T non-carriers.
Regarding the first main finding, we found that in our recessive model (CC versus CT + TT) the presence of the KIBRA rs17070145 T allele may be associated with an increased risk of AD (both aMCI due to AD and AD dementia). The post-hoc estimation of statistical power using G*Power software was 64%, indicating that our study underpowered, exposing our results to bias towards Type II error. Therefore, we cannot exclude the possibility that our non-significant results that appeared to approach the conventional threshold for statistical significance of 0.05 may cross that threshold in a properly powered study (power of >80%).
The association between the presence of the KIBRA T allele and cognitive impairment was greater in magnitude for AD dementia, with achieved power of 65%, increasing the risk of AD about six times. In contrast, the association was non-significant for aMCI due to AD, with a moderate level of power of 58%, indicating that the risk about doubled between patients with aMCI due to AD and AD dementia. So far, the association of KIBRA rs17070145 T allele and greater AD risk has provided inconsistent findings. In a study of 391 patients with AD and 428 control subjects, Rodríguez-Rodríguez and colleagues did not find any evidence of an AD risk association. Using age-specific analysis, the authors found a significant association between the KIBRA rs17070145 T allele and late-onset AD. 17 Contrary to this, Corneveaux and colleagues compared 1629 patients with AD with 936 control subjects and found that non-carriers of the T allele had a higher risk of late-onset AD compared to carriers. 19
In a meta-analysis that included 16 independent samples with 4436 cases and 4334 control subjects, Burgess and colleagues reported a statistical trend toward a decreased risk of dementia among T allele carriers (protective odds ratio for T allele 0.94, p = 0.07). 37 A meta-analysis of 20 case-control studies including 6078 cases and 5581 control subjects has shown an association between the KIBRA rs17070145 C allele and an overall increased AD risk. 20 To our knowledge, the association between MCI and KIBRA genotype has been examined in only three other studies. Almeida and colleagues found that KIBRA allele variants have minimal effect among older adults with aMCI due to AD. 9 A study by Porter and colleagues showed that KIBRA CC and APOE ε4 allele carriers with amyloid-β burden had significantly faster rates of decline in verbal episodic memory compared to KIBRA T carriers. 38 Conversely, in a recent study by Mazzeo and colleagues, 39 carrying the T allele increased the risk of developing MCI twofold compared to CC carriers.
Regarding our second main finding, KIBRA T association with memory performance suggest a nuanced relationship. Overall, our analyses did not reveal a significant association between the presence of the KIBRA T allele and memory performance. However, when we assessed this association within subsamples defined by cognitive status, we found a trend towards an association between the KIBRA T allele and better memory performance in patients with AD dementia. Several studies have demonstrated that the KIBRA T allele is associated with a small but significant improvement in episodic memory and working memory in healthy subjects,3,8,10,11,40–43 while others found little to no association between memory performance and KIBRA genotypes.12,44–46 There have been studies that found memory advantage for T allele carriers in comparison to CC homozygotes in older adults with subjective memory complaints 13 and cognitive impairment. 39 In a meta-analysis, Milnik and colleagues reported a significant association of rs17070145 with both episodic (r = 0.068, p = 0.001) and working memory (r = 0.035, p = 0.018). Their analysis indicated that rs17070145 explains 0.5% of the variance for episodic memory tasks and 0.1% of the variance for working memory tasks. 14 Our findings that KIBRA T allele increased risk of AD dementia and, simultaneously, patients with AD dementia who possessed a KIBRA T allele exhibited better memory may indicate that a) KIBRA T carriers progress towards AD diagnosis with less severity, b) KIBRA T allele does not protect fully against AD dementia diagnosis but slows subsequent decline, or c) KIBRA T conveys a longevity and/or survival effect whereby KIBRA T allele carriers are “hardier” than non-carriers.
Although we could not test these directly, our results provide opportunities to speculate about potential underlying mechanisms. Both KIBRA and APOE SNPs reduce long-term potentiation (LTP), crucial for synaptic plasticity and memory formation, by affecting glutamate receptor function.47,48 Specifically, KIBRA regulates α-amino-3-hydroxyl-5-methyl-4-isoxazole-propionate (AMPA) receptor trafficking through protein interacting with C-kinase 1 (PICK1), whereas APOE modulates N-methyl-D-aspartate (NMDA) receptor expression, affecting synaptic plasticity.47,49 In addition, KIBRA stabilizes PKMzeta (PKMζ) to regulate learning and memory and BDNF is essential for maintaining LTP precisely through PKMζ.49,50 These interactions indicate that genetic variations in KIBRA, APOE, and BDNF affect LTP in memory-related brain regions, which in turn influence cognitive performance and AD risk. The genetic interactions between KIBRA, APOE, and BDNF polymorphisms in brain networks deserve further attention as they can provide new insights into the genetic underpinnings of memory performance with age and of AD risk. The result that KIBRA T was associated with memory more strongly among the (cognitively vulnerable) APOE ε4 carriers supports the latter explanation. Laukka and colleagues found that including APOE ε4 carrier status and other genetic variants as covariates explained away the association between KIBRA CC allele and semantic memory. 44 Porter and colleagues observed an interaction between APOE ε4 and KIBRA on verbal episodic memory decline such that APOE ε4 carriers without the KIBRA T allele experienced fastest cognitive decline. 38 Along the same lines, our results also revealed a significant association between the KIBRA T allele and better memory performance among BDNF Met carriers who generally exhibit inferior episodic memory compared to Val homozygotes, 51 while no significant association was observed among BDNF Met non-carriers. Taken together, the KIBRA T allele may serve as a buffer against poor memory in adults who are otherwise particularly genetically vulnerable to memory and other cognitive deficits (i.e., the APOE ε4 or the BDNF Met carriers).
Today, genetic screening for AD is typically performed under specific circumstances, such as when individuals exhibit symptoms suggestive of the disease or have a strong family history of the condition. Since the presence of the KIBRA T allele may relate to a greater risk of aMCI due to AD and AD dementia in APOE ε4 negative individuals, routine genetic testing for individuals with symptoms, family history, or elevated risk determined by the presence of the better-known familial AD risk factor like APOE ε4, testing for KIBRA polymorphisms could enhance the ability to determine true risk well before disease onset. Finally, as treatment options become available, understanding underlying genetic predisposition across a full spectrum of relevant genes can be a crucial factor in determining treatment approach.
Due to some limitations, our results should be interpreted with caution. First, the limited sample size of control subjects and patients with aMCI due to AD and AD dementia should be taken into consideration before interpreting the data. Our results could be explained by survival bias, which may have contributed to the loss of patients with AD carrying the KIBRA CC genotype. For this reason, we controlled for age in our statistical analysis. Furthermore, we only had access to cross-sectional data. Thus, we can only infer genetic effects on individual differences in memory, not memory decline over time. In addition, of the 19 cognitively unimpaired individuals, 7 were APOE ε4 positive, which exacerbates the low power already present in our study, and which likely increases bias towards Type II error. Finally, our study was underpowered, with achieved power estimates hovering around about 60%, exposing our results to a bias toward Type II error.
Conclusion
In conclusion, we found that the KIBRA rs17070145 T allele was associated with risk of AD in the form of aMCI due to AD and AD dementia in low risk, APOE ε4 negative individuals. In addition, carriers of the KIBRA T allele with AD dementia exhibited better memory performance. Our results also observably revealed an association between the KIBRA T allele and better memory performance among AD dementia APOE ε4 carriers, albeit it did not reach or even approach significance. Taken together, the KIBRA T allele may expose the otherwise genetically lower risk individuals, i.e., those who do not carry the APOE ε4 or the BDNF Met risk alleles, to a greater chance of cognitive impairment, while it may also serve as a buffer against poor memory in adults who are otherwise particularly genetically vulnerable to memory deficits, i.e., those who carry the APOE ε4 or the BDNF Met gene. Overall, the conflicting results across studies that vary in terms of memory tasks used, as well as the age and/or cognitive status of participants should motivate further research into the role of KIBRA in progression across stages of cognitive status and variability in memory functioning. Specifically, methodological improvements such as larger, longitudinal studies, the inclusion of more diverse populations in terms of ethnicity or socioeconomic status, and the use of standardized memory tasks should be considered to enhance the reliability and generalizability of the findings.
Footnotes
Acknowledgments
The authors have no acknowledgments to report.
Author contributions
Vanesa Jurasova (Data curation; Funding acquisition; Investigation; Methodology; Writing – original draft; Writing – review & editing); Ross Andel (Conceptualization; Data curation; Funding acquisition; Investigation; Methodology; Writing – original draft; Writing – review & editing); Alzbeta Katonova (Data curation; Investigation; Writing – review & editing); Raena Nolan (Data curation; Methodology; Writing – review & editing); Zuzana Lacinova (Data curation; Methodology; Writing – review & editing); Tereza Kolarova (Data curation; Methodology; Writing – review & editing); Vaclav Matoska (Data curation; Funding acquisition; Investigation; Methodology; Writing – review & editing); Martin Vyhnalek (Data curation; Funding acquisition; Investigation; Methodology; Writing – review & editing); Jakub Hort (Conceptualization; Data curation; Funding acquisition; Investigation; Methodology; Writing – original draft; Writing – review & editing).
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by the project National Institute for Neurological Research (Programme EXCELES, ID Project No. LX22NPO5107)—Funded by the European Union – Next Generation EU; by the Charles University, project GA UK No. 496122; by the CETOCOEN Excellence TEAMING II project supported by Horizon 2020 (No. 857560); by the CETOCOEN Excellence project (CZ.02.1.01/0.0/0.0/17_043/0009632); and by the Alzheimer Foundation Czech Republic.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability
The data supporting the findings of this study are available on request from the corresponding author. The data are not publicly available due to privacy or ethical restrictions.