
Abstract
Background
Blood pressure variability (BPV) is associated with neurodegeneration and cognitive decline independent of average pressure. The effect of parasympathetic central autonomic network (CAN) impairment on this relationship has not been assessed.
Objective
Determine whether parasympathetic CAN network function affects the relationship between BPV and neurodegenerative markers.
Methods
100 independently living older adults (55–89 years) underwent continuous blood pressure monitoring, neuropsychological testing, venipuncture, and brain MRI. Hippocampal volumes and entorhinal cortex thicknesses were assessed. Functional connectivity within a parasympathetic cardiovascular control network was used as a measure of parasympathetic CAN function. Plasma glial fibrillary acidic protein (GFAP) and neurofilament light (NfL) were used as measures of glial and neuronal injury, respectively.
Results
Elevated BPV was associated with left hippocampal atrophy (p = 0.03) and elevated plasma GFAP (p = 0.005) independent of age, sex, vascular risk factor burden, total intracranial volume (when applicable) and average blood pressure. These relationships were not mediated by parasympathetic central autonomic network impairment. Instead, parasympathetic CAN impairment conferred a vulnerability to elevated BPV. In participants with decreased parasympathetic CAN connectivity elevated BPV was associated with left entorhinal cortex atrophy (p = 0.0001), elevated plasma GFAP (p = 0.0001), elevated plasma NfL (p = 0.001), and memory impairment (p = 0.007).
Conclusions
Findings suggest elevated beat-to-beat BPV is directly related to brain injury, and this effect is not mediated by CAN dysfunction. Instead, CAN impairment may confer a susceptibility to glial and neuronal injury in older adults with elevated beat-to-beat blood pressure variability. Mechanisms underlying increased susceptibility to BPV elevation in those with CAN dysfunction warrants further study.
Keywords
Introduction
Increased blood pressure variability (BPV) occurs with aging and vascular disease and is associated with medial temporal lobe tau pathology, 1 neurodegeneration, 2 cognitive impairment, 3 Alzheimer's disease (AD), 4 decreased cerebral perfusion, 5 decreased cerebrovascular reactivity, 6 and cerebral small vessel disease (CSVD).7,8 The effect of increased BPV on brain injury may be more pronounced in individuals with low heart rate variability. 4 This raises a novel possibility that healthy parasympathetic function blunts the adverse effects of elevated BPV. This protective effect may be conferred via an independent mechanism associated with parasympathetic function, such as dynamic cerebral autoregulation,9,10 or another yet to be identified mechanism.
Parasympathetic function is modulated in part by the brain's central autonomic network (CAN), a network of interconnected cortical and subcortical brain regions. 11 Neurodegenerative disease may be associated with autonomic dysfunction, 12 since neurodegeneration can occur within these CAN regions. 13 This raises the possibility that parasympathetic network dysfunction and elevated BPV interact to create a vicious neurodegenerative cycle. As BPV increases with age, neurodegeneration in parasympathetic CAN brain regions could create a susceptibility to the adverse effects of BPV.
In a previous analysis, 14 we studied the effects of central autonomic impairment in the context of CSVD, a pathology associated with neurodegeneration. 15 In that study a moderating effect was reported, such that individuals with decreased CAN function were most susceptible to the adverse effects of elevated BPV. This could be due to impairment of another physiological processes regulated by the CAN, such as dynamic cerebral autoregulation. 16 The aim of the present study is to test whether parasympathetic CAN functional impairment confers a similar susceptibility to BPV-related neurodegeneration in brain regions susceptible to AD pathology in older adults. Alternatively, parasympathetic CAN functional impairment might mediate the relationship between BPV elevation and neurodegeneration, which could suggest reverse causation whereby neurodegeneration within CAN regions drives autonomic dysfunction and BPV elevation. Thus, both mediation and moderation models were tested in the present study to distinguish between these possibilities. Characterization of the parasympathetic CAN's relevance to neurodegenerative disease processes will help clarify its role as either a susceptibility factor, resilience factor, or therapeutic target.
Methods
Participants
Participants were recruited from Orange County communities through outreach events, mailing lists, word-of-mouth, online portals, a research volunteer registry, and through the University of California Irvine (UCI) Alzheimer's Disease Research Center (ADRC), and all procedures were conducted as part of the Vascular Senescence and Cognition (VaSC) Study at UCI. Older adults aged 55 to 89 years who were living independently were included (Table 1). Study exclusions were a prior diagnosis of dementia, history of clinical stroke, family history of dominantly inherited neurodegenerative disorders, current neurological or major psychiatric disorders that may impact cognitive function, history of moderate-to-severe traumatic brain injury, current use of medications impairing the central nervous system, current organ failure or other uncontrolled systemic illness, and contraindications for brain MRI. Eligibility for the study was verified by a structured clinical health interview and review of current medications with the participant and, when available, a knowledgeable informant study partner. This study was approved by the UCI Institutional Review Board, and all participants gave informed consent. The anonymous data that support the findings of this study are available upon reasonable request from the corresponding author, DAN, through appropriate data sharing protocols.
Participant characteristics and demographics (N = 100).

A: plasma biomarker subset n = 52, APOE4: apolipoprotein ε4; BPV: blood pressure variability indexed by systolic average real blood pressure variability; CAN: central autonomic network; GFAP: glial fibrillary acidic protein; NfL: neurofilament light chain.
Continuous BP data acquisition
Beat-to-beat BP measurements were obtained continuously during supine rest in a 3 T Siemens MRI scanner, using an MRI compatible non-invasive continuous BP finger cuff device (Biopac®). Participants were asked to take medications as normally prescribed and abstain from caffeine the morning of data collection. After calibration, continuous BP was monitored during MRI for 7 min and further data processing was performed as previously described to remove motion artifacts and low quality data segments. 17
Systolic blood pressure average real variability
BPV was quantified as systolic average real variability, defined as the average of absolute changes between consecutive blood pressure readings as described elsewhere.17,18 This measure of BPV has been shown to be more reliable and is less susceptible to outliers,17,19 and is calculated as shown: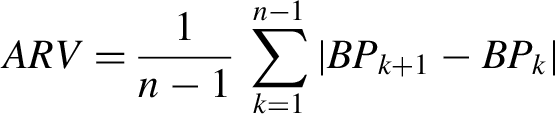
In this formula n represents the total number of systolic blood pressure readings taken during seven minutes of continuous blood pressure recording while represents a systolic blood pressure reading index.
Vascular risk factors
Vascular risk factor (VRF) burden was determined through clinical interviews with the participant and informant (when available), and review of current medications and medical history. The assessed VRFs included history of cardiovascular disease, hypertension, hyperlipidemia, type 2 diabetes, atrial fibrillation, left ventricular hypertrophy and transient ischemic attack. Total VRFs were summed for each participant and elevated VRF burden was defined as ≥2 VRFs (versus 0–1) as described previously.20,21
Plasma biomarkers
Plasma levels of GFAP and NfL were determined using single molecule array, (Simoa®) Neurology 2-Plex B (N2PB) Kit (Quanterix), following the manufacturer's protocol on the HD-X machine as previously described. 22
APOE genotyping
Fasted blood samples were obtained by venipuncture and used to determine participant APOE genotype. Genomic DNA was extracted using the PureLink Genomic DNA Mini Kit (Thermo). APOE genotyping was performed as previously described.14,23,24 All analyses were performed at the same lab at the University of Arizona (KR).
Neuroimaging
All participants underwent brain MRI scans conducted on a 3 T Siemens Prisma scanner with 20-channel head coil. High-resolution 3D T1-weighted anatomical (Scan parameters: TR = 2300 ms; TE = 2.98 ms; TI = 900 ms; flip angle = 9 deg; FOV = 256 mm; resolution = 1.0 × 1.0 × 1.2 mm3; Scan time = 9 min) images were acquired, using 3-dimensional magnetization-prepared rapid gradient-echo (MPRAGE) sequences. Post-processing of T1 scans was accomplished in FreeSurfer 7.4.1 using a semi-automated quantification algorithm 25 according to methods described elsewhere. 24 After segmentation, individual data were manually checked for inaccuracies. Where inaccuracies were identified, corrections were made using the built in FreeSurfer editing tools. Data were then reprocessed and included for analysis.
Parasympathetic central autonomic network connectivity quantification
The CAN chosen for this analysis was derived from a voxel-based meta-analysis of 43 studies and includes 9 brain regions that form a central network associated with high frequency heart rate variability. 26 Functional and anatomical data were preprocessed using CONN (RRID:SCR_009550) release 22.a and SPM (RRID:SCR_007037) release 12.7771, 27 according to methods described elsewhere. 22
ROI-to-ROI connectivity matrices were estimated characterizing the patterns of functional connectivity within selected regions of interest (Figure 1). Functional connectivity strength was represented by Fisher-transformed bivariate correlation coefficients from a weighted general linear model (weighted-GLM 28 ), defined separately for each pair of target areas, then averaged.
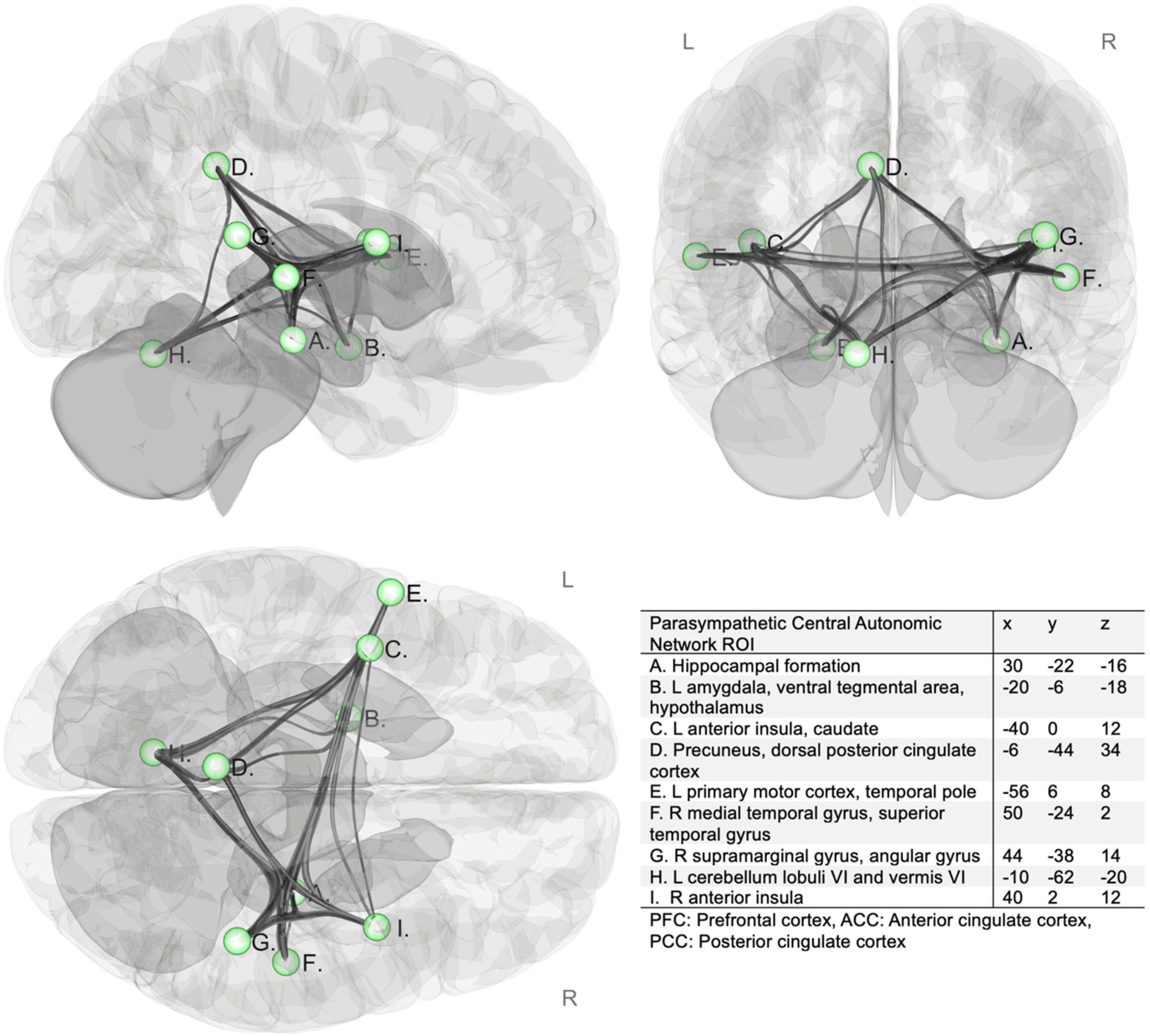
Parasympathetic central autonomic network regions of interest (ROIs). 26 L: left; R: right. Network regions of interest (ROI) are 5 mm spheres illustrated in green.
Neuropsychological testing
All participants underwent a clinical interview and comprehensive neuropsychological assessment by a trained technician or doctoral student under the supervision of a licensed clinical neuropsychologist. The assessment included multiple tests of memory, attention/executive function, and language. All neuropsychological testing and diagnostic assessments were conducted blinded to all clinical, biomarker and imaging findings. A memory composite score was created by averaging the demographically corrected (age, sex, and education) memory test z-scores. Participants underwent one of two memory batteries based on their recruitment source. Participants recruited from the community underwent the Rey Auditory Verbal Learning Test (RAVLT) word list learning test and the Wechsler Memory Scale – Revised (WMS-R) 29 Logical Memory story memory test, and those recruited from the UCI ADRC underwent the California Verbal Learning Test 30 – Second Edition (CVLT-II) word list learning test and the Craft Stories story memory test. The two different word list learning (RAVLT and CVLT-II) and story memory (WMS-R Logical Memory and Craft Stories) tests are very similar in structure and have previously been studied together and determined to be largely interchangeable.31,32 Thus, demographically-corrected z-scores were computed for the word list learning long delay free recall (RAVLT Trial 7 and CVLT-II long delay free recall) and story memory delayed free recall (WMS-R Logical Memory II and Craft Stories delayed free recall) and averaged to generate a composite memory score.33–36
Statistical analyses
101 participants underwent brain fMRI, anatomical T1w MRI, neuropsychological testing, and continuous BP monitoring. 52 participants also had plasma NfL and GFAP biomarker assays performed at the time of analysis. All data was screened for outliers (±3SD) and 1 outlier was identified and removed with left hippocampal volume +4.35 SD and right hippocampal volume +4.31 SD. After outlier screen, the total overall analyzed sample was N = 100 and for plasma biomarker analyses a subset of n = 52.
Multiple linear regression analyses were used to assess the effects of BPV on nine dependent variables: left/right entorhinal cortex thicknesses, left/right hippocampal volumes, memory composite score, executive function composite score, language composite score, plasma GFAP, and plasma NfL. Where significant relationships were identified Hayes Process macro 37 model 4 (simple mediation) was used in R 38 to test the potential mediating effects of parasympathetic CAN connectivity on the effect of BPV where x = BPV, Y = dependent variable, and m = parasympathetic CAN ROI-to-ROI connectivity matrix. Hayes PROCESS Model 1 (simple moderation) was used to assess the potential moderating effect of parasympathetic CAN functional connectivity on the relationship between BPV and the dependent variables where x = BPV, Y = dependent variable, and w = parasympathetic CAN ROI-to-ROI connectivity matrix. All variables used in moderation analyses are continuous variables. Statistical results are therefore based on the continuous relationship between independent and dependent variables conditional upon the value of the continuous moderator. However, for visualization of moderation results, parasympathetic CAN connectivity will be transformed into a categorical variable (+1 SD, mean, and −1 SD). These categories are for display purposes only, and do not represent clinically significant cutoff values. Linear regression assumptions regarding linearity, multicollinearity (VIF < 5), and homoscedasticity (Breusch-Pagan test) were met.
Minimum detectable effect sizes given a power of 0.80, alpha of 0.05, and sample of 100 for mediation analyses was calculated using Monte Carlo simulations 39 as 0.07 for the indirect effect, or 0.15 in the biomarker subset (n = 52) analysis. For moderation analyses 40 the minimum detectable R2 change given a power of 0.80, alpha of 0.05, and N = 100, is 2.5%, or 6% in the biomarker subset (n = 52) analysis.
Results
100 participants were included for analysis, participant characteristics and demographics for this sample are shown in Table 1. No significant differences in participant characteristics were observed between the overall sample and the plasma biomarker subset (Supplemental Table 1).
Increased beat-to-beat BPV was associated with decreased left hippocampal volume (B = −99.89, p = 0.03) and increased plasma GFAP (B = 31.44, p = 0.005) independent of age, sex, VRF, mean systolic blood pressure, and where applicable TIV. These direct effects were not statistically mediated by parasympathetic CAN functional connectivity decline (Figure 2A-D). BPV was not associated with right hippocampal volume (B = −72.30, p = 0.11), left entorhinal cortex thickness (B = −0.08, p = 0.07), right entorhinal cortex thickness (B = 0.06, p = 0.15), or plasma NfL (B = 1.93, p = 0.17) after adjustment for age, sex, VRF burden, and mean systolic blood pressure.
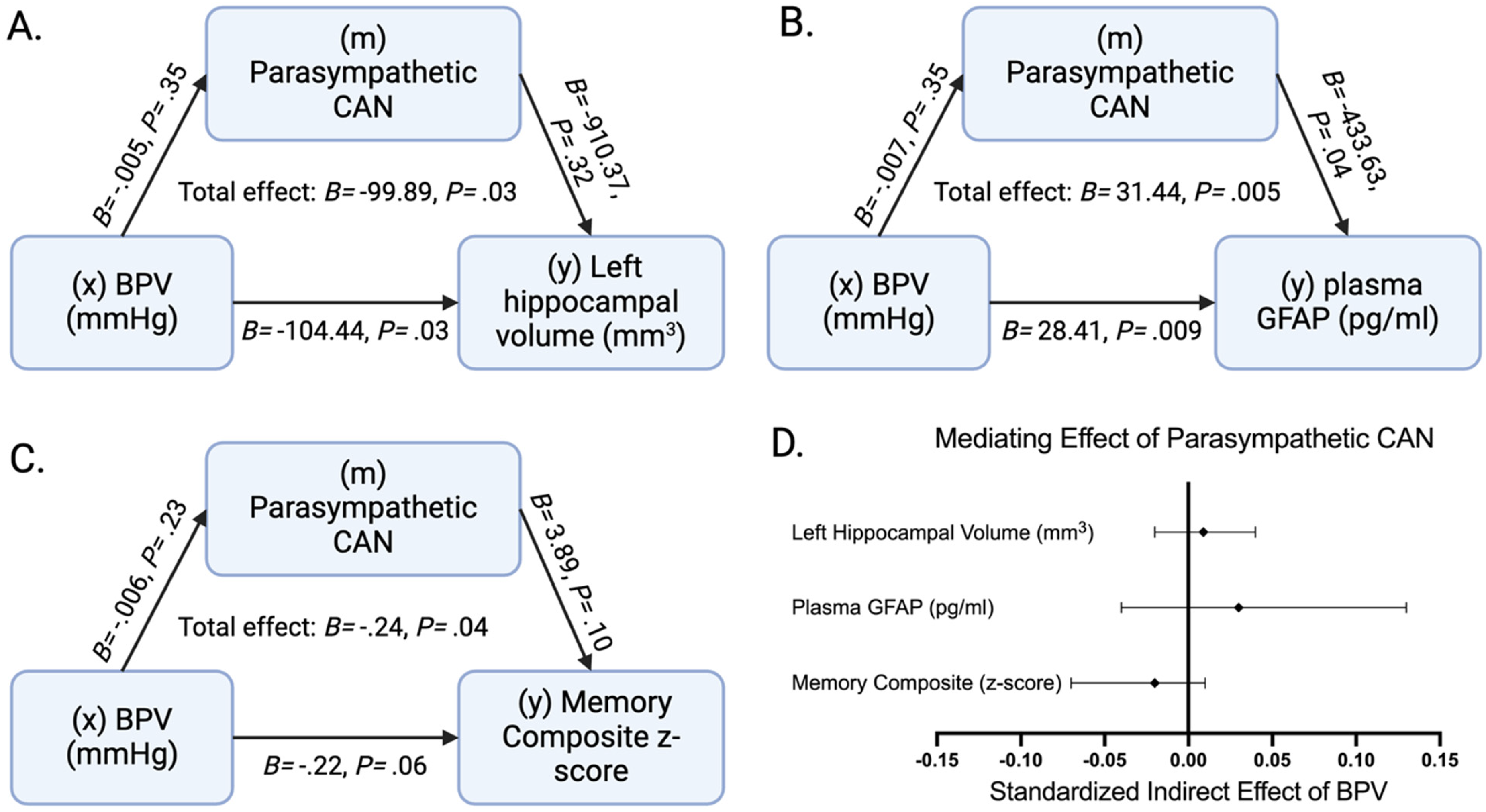
The potential mediating effect of parasympathetic central autonomic network (CAN) functional connectivity. The potential mediating role of parasympathetic CAN functional connectivity on the adverse effect of elevated blood pressure variability (BPV) on A: left hippocampal volume (N = 100) adjusted for age, sex, total intracranial volume, vascular risk factor burden, and average systolic blood pressure, B: plasma GFAP (n = 52) adjusted for age, sex, vascular risk factor burden and average systolic blood pressure, and C: demographically (age, sex, and education) adjusted memory composite z-score (N = 100) adjusted for vascular risk factor burden and average systolic blood pressure is tested. D: The mediating effects of parasympathetic CAN are displayed as the standardized indirect effect of BPV with 95% confidence intervals from the three displayed mediation models. B = unstandardized β, p = p-value.
In moderation analyses, the interaction between BPV and parasympathetic CAN function was associated with left entorhinal cortex (ROI shown in Figure 3A) thickness (B = 3.78, p = 0.001; Figure 3B), plasma GFAP (B = −685.91, p = 0.01; Figure 3C), and plasma NfL (B = −90.72, p = 0.01; Figure 3D) and remained so after adjustment for age, sex, mean systolic blood pressure, VRF burden, and where appropriate, total intracranial volume (Figure 3B-3D). In participants with low parasympathetic CAN functional connectivity, increased BPV was associated with decreased left entorhinal cortex thickness (B = 0.20, p = 0.0001; Figure 3B), increased plasma GFAP (B = 50.27, p = 0.0001; Figure 3C), and increased plasma NfL (B = 5.21, p = 0.001; Figure 3D). The BPV-parasympathetic CAN functional connectivity interaction was not significantly associated with left hippocampal volume (B = 2282.32, p = 0.13), right hippocampal volume (B = 1141.65, p = 0.41), or right entorhinal cortex thickness (B = 2.15, p = 0.08).
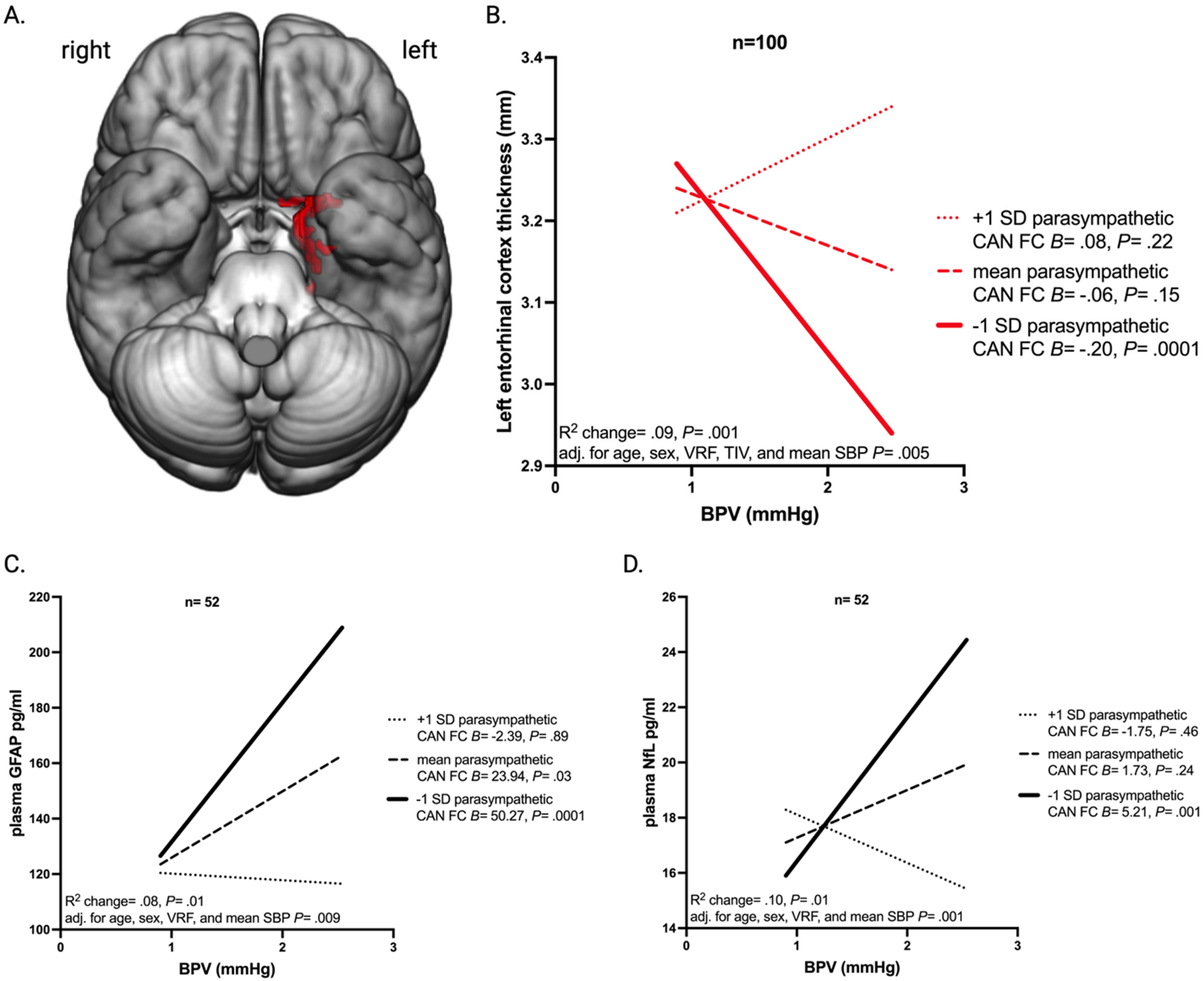
Conditional and interactive effects of beat-to-beat BPV. (A) Rendering of the left entorhinal cortex region of interest. (B) The relationship between blood pressure variability (BPV) indexed as beat-to-beat systolic average real variability and left entorhinal cortex thickness conditional upon parasympathetic central autonomic network (CAN) functional connectivity (FC) (−1 SD, mean, and +1 SD groupings for display purposes). (C) The relationship between BPV and plasma GFAP conditional upon parasympathetic CAN FC (−1 SD, mean, and +1 SD). (D) The relationship between BPV and plasma NfL conditional upon parasympathetic CAN FC (−1 SD, mean, and +1 SD). Panels B, C, and D display the within group unstandardized beta coefficients (B) and p-values (p), the change in R2 associated with the addition of the interaction term, the p associated with the interaction term, and the age, sex, vascular risk factor burden (VRF), and mean systolic blood pressure (SBP) adjusted interaction term p.
The interaction between BPV and parasympathetic CAN function was also associated with demographically (age, sex, and education) adjusted memory composite z-score (B = 6.61, p = 0.046) independent of mean systolic blood pressure, and VRF burden adjustment (Figure 4). Specifically, in participants with low parasympathetic CAN functional connectivity, increased BPV was associated with decreased memory composite z-score (B = 0.09, p = 0.007; Figure 4). No significant relationship between the BPV and Parasympathetic CAN interaction term and executive function or language composite z-scores was found.
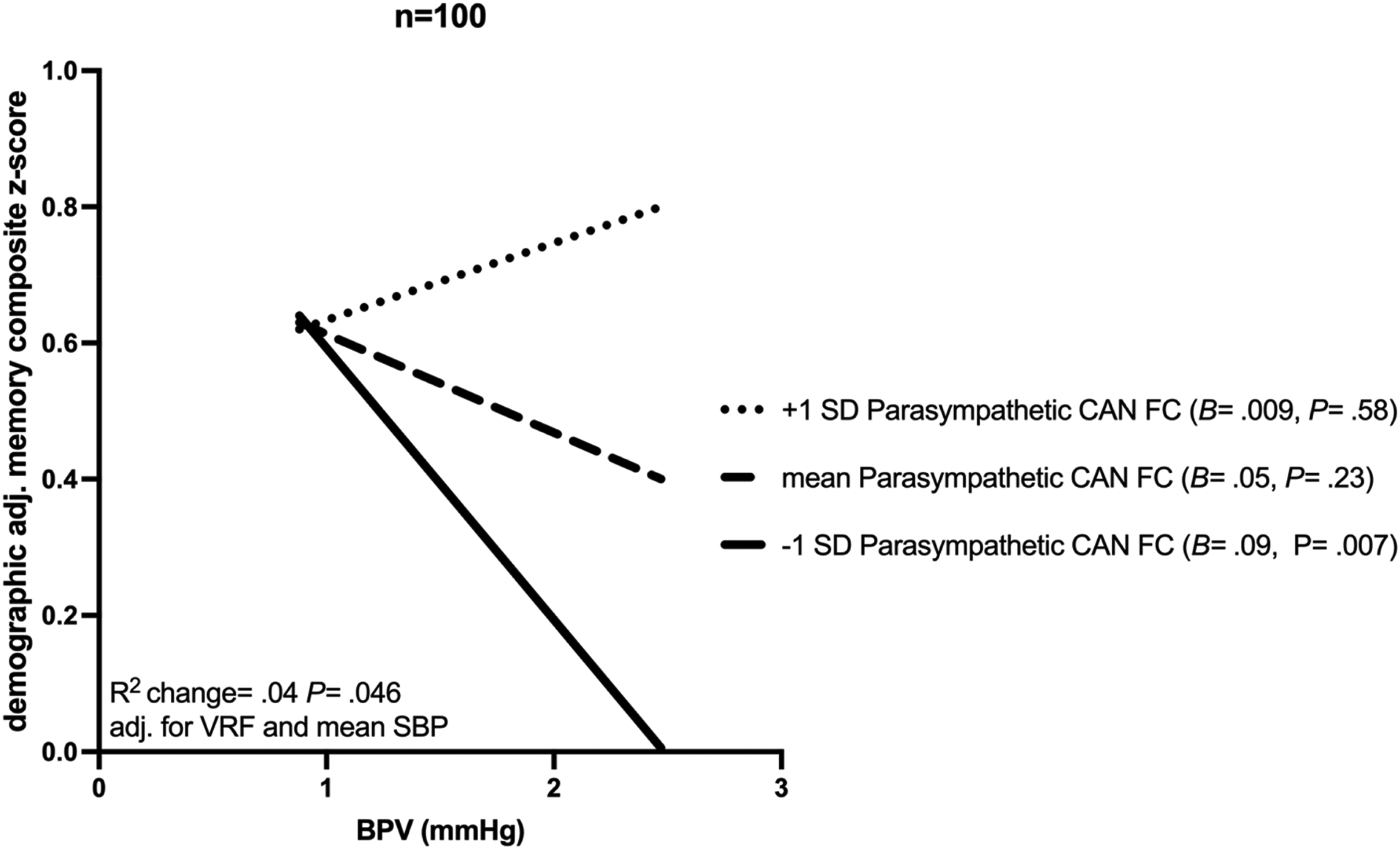
Conditional and interactive effects of beat-to-beat BPV. The relationship between blood pressure variability (BPV) indexed as beat-to-beat systolic average real variability and demographically (age, sex, and education) adjusted memory composite z-score conditional upon parasympathetic central autonomic network (CAN) functional connectivity (FC) (−1 SD, mean, and +1 SD groupings for display purposes). The unstandardized beta coefficients (B) and p-values (P) are displayed for each group in the figure legend, while the change in R2 and the p associated with the interaction term adjusted for vascular risk factor burden (VRF), and mean systolic blood pressure (SBP) are displayed along the x-axis.
Benjamini-Hochberg false discovery rate (FDR) correction 41 was also applied for the three interaction effects plus the nine subgroup analyses. All significant findings survived FDR correction.
Discussion
Elevated beat-to-beat BPV was independently associated with left hippocampal atrophy, memory impairment, and elevated plasma GFAP as reported previously. 24 These relationships were not mediated by parasympathetic CAN functional decline, failing to support the hypothesis 42 that elevated BPV is merely a consequence of structural and functional changes within central autonomic brain regions associated with AD pathophysiology. Instead, parasympathetic CAN functional connectivity moderated the effect of elevated BPV on left entorhinal cortex volume, memory impairment, plasma GFAP, and plasma NfL. This supports our a priori hypothesis that parasympathetic central autonomic functional decline may impart a susceptibility to elevated beat-to-beat BPV-related neurodegeneration including atrophy in a brain region susceptible to early AD pathology, 43 and elevation of two non-specific biomarkers of neurodegeneration, GFAP, a biomarker of glial injury,44–46 and NFL, a biomarker of neuroaxonal injury.47–50
The hypothesis that BPV and parasympathetic impairment may interact to confer neurodegenerative risk has been previously demonstrated in a large clinical sample of 48,204 adults. 4 In that investigation the authors determined that participants in both the highest quartile of BPV and lowest quartile of heart rate variability displayed increased dementia risk. Heart rate variability is a useful clinical measure of parasympathetic function, 51 and previous research suggests that parasympathetic function may be related to dynamic cerebral autoregulation. 9 Specifically, cholinergic blockade leads to an increased transfer function coherence between blood pressure and cerebral blood flow, which could indicate a reduction in dynamic cerebral autoregulation.9,10 Our laboratory recently found a relationship between functional connectivity within brain regions associated with parasympathetic function and plasma biomarkers of neurodegeneration in healthy older adults, 22 raising the possibility that central autonomic dysfunction may be implicated early in AD pathogenesis.
This relationship between the cholinergic system and cerebral autoregulation provides a potential mechanism for the observed interaction in the present study. These findings could indicate that rather than neurodegeneration in central autonomic regions causing BPV, it instead creates a susceptibility to elevated BPV which leads to accelerated neurocognitive pathology. However, additional research is needed to fully elucidate the relationship between CANs and dynamic cerebral autoregulation before further conclusions can be drawn.
The present study adds confirmation that increased BPV is not fully explained by autonomic impairment related to neurodegeneration as has been proposed. 42 Instead it suggests an interactive effect between parasympathetic impairment and elevated BPV similar to what we previously reported in the context of CSVD. 14 Our findings suggest that parasympathetic CAN impairment represents an independent susceptibility factor, perhaps due to its relationship with cerebral autoregulation, rather than a source of elevated BPV. Future studies should examine the longitudinal relationships between central autonomic function, peripheral hemodynamic variables, and neurodegeneration to confirm the present study findings. Although we cannot infer causality from these cross-sectional findings, future confirmation in a longitudinal analysis may provide evidence that elevated beat-to-beat BPV is a potentially druggable causal factor in neurodegeneration rather than merely a symptom of neurodegenerative disease which would have significant clinical implications.
Strengths of the present study include convergent findings using neuropsychological testing, neuroimaging, and plasma biomarkers of neurodegeneration. An additional strength includes the use of a CAN derived from a voxel-based meta-analysis to account for the CAN heterogeneity present in literature. 26 Study limitations include the cross-sectional design which limits causal interpretation, a relatively small sample size which may limit generalizability, and the availability of only a subset of participants for plasma biomarker analysis.
Although additional confirmation is needed in larger, longitudinal samples, the present study implications include the potential ability to identify individuals at elevated neurodegenerative risk due to BPV. This might inform early interventions, risk stratification, or personalized therapeutic approaches designed to target BPV reduction in those individuals through aggressive BP management, an increased focus on antihypertensive medication adherence, or other interventions designed to mitigate BPV.
Supplemental Material
sj-docx-1-alz-10.1177_13872877251376715 - Supplemental material for Impaired parasympathetic network function confers susceptibility to neurodegeneration and memory impairment in older adults with elevated beat-to-beat blood pressure variability
Supplemental material, sj-docx-1-alz-10.1177_13872877251376715 for Impaired parasympathetic network function confers susceptibility to neurodegeneration and memory impairment in older adults with elevated beat-to-beat blood pressure variability by Trevor Lohman, Isabel Sible, Arunima Kapoor, Allison C Engstrom, Fatemah Shenasa, John Paul M Alitin, Aimee Gaubert, Kathleen E Rodgers, David Bradford, Mara Mather, S Duke Han, Elizabeth Head, Lorena Sordo, Julian F Thayer and Daniel A Nation in Journal of Alzheimer's Disease
Footnotes
Acknowledgements
We thank all participants for contributing their time to this study and for their continued commitment to this research. This research was supported by National Institutes of Health grants (DAN: R01AG064228, R01AG060049, R01AG082073, P01AG052350, P30AG066530), (SDH: K24AG081325), (EH: P30AG066519), the American Heart Association (AK: 23PRE1014192), and the Southern California Clinical and Translational Science Institute (TL: KL2TR001854).
Ethical considerations
This study was approved by the University of California, Irvine Institutional Review Board (HS-2019-5324).
Consent to participate
All participants gave written informed consent.
Consent for publication
Not applicable
Author contributions
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by National Institutes of Health grants (DAN: R01AG064228, R01AG060049, R01AG082073, P01AG052350, P30AG066530), (SDH: K24AG081325), (EH: P30AG066519), the American Heart Association (AK: 23PRE1014192), and the Southern California Clinical and Translational Science Institute (TL: KL2TR001854).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
The anonymous data that support the findings of this study are available upon reasonable request from the corresponding author, DN, through appropriate data sharing protocols.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.