
Abstract
Background
Cognitive decline represents a major challenge in aging populations. Probiotics have been proposed to influence cognitive function through gut–brain interactions, but clinical findings remain inconsistent.
Objective
This study evaluated the effects of probiotic supplementation on cognitive function as the primary outcome, and on BDNF levels, inflammatory markers, and oxidative stress biomarkers as secondary outcomes in adults aged 50 years and older.
Methods
A systematic search of PubMed, EBSCO, ProQuest, and Google Scholar was conducted through 1 May 2024 using predefined search terms related to probiotics, cognitive function, BDNF, inflammation, and antioxidant activity. Study quality was assessed using the RoB 2 tool. Meta-analyses were performed using random-effects models, and publication bias was explored using Egger's test where study counts permitted.
Results
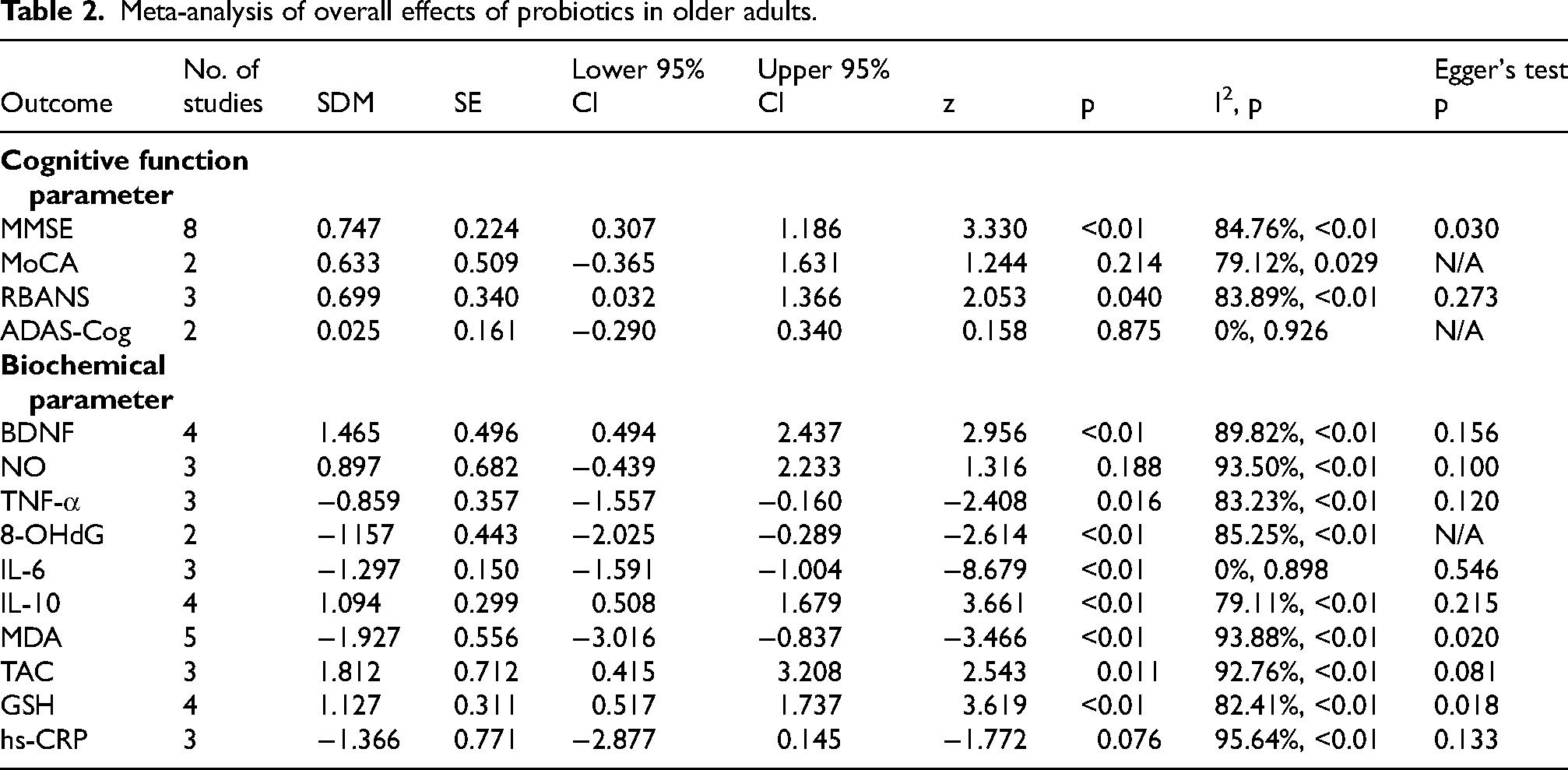
Sixteen studies demonstrated significant improvement in cognitive function among participants receiving probiotics compared to placebo. Cognitive function, measured using the Mini-Mental State Examination (MMSE), yielded a standardized mean difference (SMD) of 0.747 (95% CI 0.307–1.186) which corresponds to moderate-to-large effects. In comparison, the Repeatable Battery for the Assessment of Neuropsychological Status (RBANS) showed significant results with an SMD of 0.340 (95% CI 0.032–1.366) which corresponds to small-to-moderate effects. Probiotics also led to significant changes in several biochemical parameters, including BDNF, TNF-α, 8-OHdG, IL-6, IL-10, MDA, TAC, and GSH. Multi-strain probiotics showed better results compared to single-strain.
Conclusions
Probiotic supplementation may offer modest cognitive benefits in aging populations, particularly in studies enrolling cognitively impaired individuals, but substantial heterogeneity and limited biomarker evidence restrict the certainty of these findings. Larger, longer-duration, and standardized trials are needed to clarify the clinical relevance and potential biological pathways underlying probiotic effects on cognition.
Introduction
Aging is a complex process characterized by a gradual loss of cognitive function, elevated inflammatory levels, and decreased levels of antioxidant activity.1–3 The aging of the world's population is causing a rise in the prevalence of neurological disorders including Alzheimer's disease (AD) and cognitive decline, which presents serious public health issues.4,5 Cognitive decline is one of the most devastating features of aging, affecting memory, learning, and general quality of life. Several investigations on the relationship between gut bacteria and brain function have identified the gut-brain axis as a bidirectional communication mechanism involving neural, immune and metabolic pathways, a biologically plausible mechanism through which gut microbiota may influence the central nervous system. 6 While studies support the possibility that modulating gut microbiota can affect neuroinflammation, neurotrophic signaling and oxidative stress, human randomized controlled trials (RCTs) have produced mixed and heterogenous findings. 7 Thus, probiotics supplementation should be viewed as a potential modulator of neurobiological pathways rather than definitively altering central neurobiology.
Existing evidence suggests that probiotics improve cognitive function in older adults, though the exact mechanisms are still under investigation. Several studies have demonstrated changes in brain-derived neurotrophic factor (BDNF), inflammatory markers, and antioxidant activity in individuals taking probiotics. BDNF is a crucial neurotrophin that plays a key role in synaptic plasticity, neuronal survival, and growth. 8 Several neuropsychiatric diseases and cognitive decline are associated with lower levels of BDNF. 9 Patients with AD exhibit significantly lower serum BDNF levels compared to healthy individuals. 10 According to the research, probiotics such as Lactobacillus and Bifidobacterium may raise BDNF levels, which would benefit person's cognitive health. 11 Aside from cognitive decline, aging is linked to increased systemic inflammation, known as “inflammaging,” and a decrease in the body's antioxidant defenses.12,13 Cognitive impairment and neurodegenerative diseases have been linked to elevated levels of pro-inflammatory cytokines, including tumor necrosis factor-alpha (TNF-α) and interleukins (IL-6), as well as oxidative stress markers, such as 8-hydroxy-2′-deoxyguanosine (8-OHdG) and malondialdehyde (MDA).3,14–17 It is thought that probiotics have anti-inflammatory and antioxidant properties, which may help to mitigate these harmful processes. Probiotics improve gut health by balancing the gut microbiota, increasing mucus secretion, and protecting tight junction proteins by lowering lipopolysaccharide (LPS) levels, which prevents LPS from activating inflammatory responses via toll-like receptors.18,19 Probiotics help prevent overactivation of the immune system and lower inflammatory indicators by reducing intestinal permeability and gut dysbiosis. They also promote the production of Th2 cytokines including IL-4 and IL-10 and help in T-cell differentiation. 18 Superoxide dismutase (SOD) and other antioxidant enzymes are produced by probiotics, which can counteract oxidants. 20 Their capacity to bind metal ions, which can cause oxidative stress by degrading hydrogen peroxide or starting lipid peroxidation, is another important component of their antioxidant function. For example, it has been demonstrated that Lactobacillus rhamnosus GG and L. paracasei Fn032 considerably lessen hydrogen peroxide generation brought on by ferrous ions. 21 There is growing evidence of the utilization of probiotics in older adults; however, the association of probiotics with these markers has mixed results. Therefore, there is a need for conducting a systematic review to accumulate existing evidence regarding this topic, particularly in older adults.
Probiotics have been proposed as a potential strategy for supporting cognitive health in older adults through their influence on the microbiota–gut–brain axis. Preclinical studies demonstrate that gut microbes can affect neuroinflammatory, metabolic, and neurotrophic pathways, suggesting a biologically plausible mechanism by which probiotics may modulate brain function. However, clinical evidence in humans remains heterogeneous. While some randomized controlled trials have reported improvements in cognitive performance or related biomarkers, others show minimal or no effect, and differences in probiotic strains, dosages, intervention duration, and participant populations complicate interpretation.8–13 Therefore, the overall therapeutic potential of probiotics for cognitive aging remains uncertain. In this context, cognitive function was assessed using validated neuropsychological scales such as the Mini-Mental State Examination (MMSE) and the Repeatable Battery for the Assessment of Neuropsychological Status (RBANS) was defined as the primary outcome of interest. We also examined several secondary outcomes representing mechanistic pathways implicated in cognitive aging: (1) neurotrophic factors (e.g., BDNF), due to their role in synaptic plasticity and neuroprotection; (2) inflammatory markers (e.g., IL-6, TNF-α), which reflect neuroimmune modulation relevant to aging and neurodegeneration; and (3) oxidative stress and antioxidant biomarkers (e.g., 8-OHdG, MDA, total antioxidant capacity (TAC), glutathione (GSH)), given the recognized contributions of oxidative imbalance to cognitive decline. These biomarkers were selected to explore whether probiotic-induced physiological changes align with pathways theorized to influence brain aging. Our objective is to systematically evaluate evidence from randomized controlled trials to determine whether probiotic supplementation improves cognitive function in older adults and to assess whether changes in neurotrophic, inflammatory, and oxidative stress markers support plausible biological mechanisms relevant to clinical practice.
Methods
This systematic review and meta-analysis were conducted following the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) statement guidelines. This study was registered in PROSPERO (CDR42024582428). The objective was to evaluate the effects of probiotics on cognitive function, BDNF levels, inflammation markers, and antioxidant activity in aging populations. In accordance with our protocol, “aging” was operationally defined as adults aged ≥50 years. This threshold was selected to capture early age-related cognitive vulnerability while accommodating the inclusion criteria commonly used in probiotic RCTs.
Eligibility criteria
Studies were included if they met the following criteria: 1) Study design: RCT; 2) Population: human participants aged 50 years and older; 3) Intervention: probiotic or probiotic fortified products; 4) Comparator: placebo; 5) Outcomes: Cognitive function is the primary outcome, while BDNF levels, inflammation markers, and antioxidant activity are secondary outcomes; 6) Studies reporting sufficient data to calculate effect sizes.
The exclusion criteria were: 1) Non-human studies; 2) Observational studies, case reports, reviews, and editorials.
Search strategy
A comprehensive literature search was performed across several electronic databases, including PubMed, EBSCO, ProQuest, and Google Scholar supplemented by extensive manual searching of reference lists, and forward/backward citation tracking, from inception to 1 May 2024. The search strategy included a combination of keywords and Medical Subject Headings (MeSH) terms related to probiotics, aging or older adults, cognitive function, BDNF, inflammation, and antioxidant activity. The detailed search strategy is in Supplemental Table 1.
Data selection, collection, and extraction
Titles and abstracts were separately checked for eligibility by two reviewers. Full-text papers were acquired and evaluated for relevance to consider for inclusion. A third reviewer was consulted or discussed to address any discrepancies. The study's author, year, country, design, participant characteristics (sample size, population cognitive health status), intervention details (probiotic strain [single- or multi-strain], active bacteria, duration of studies), and outcomes were among the data that were extracted. The primary outcome was cognitive function measured by any validated cognitive measurement tools, while the secondary outcomes included BDNF, NO, TNF-α, 8-OHdG, Il-6, IL-10, MDA, TAC, GSH, and high-sensitivity C-reactive protein (hs-CRP).
Quality assessment
The quality of the included studies was assessed using RoB 2 tools.
Data analysis and synthesis
Meta-analyses were conducted using Comprehensive Meta-Analysis (CMA) software. CMA implements the random-effects model using the DerSimonian and Laird estimator, and we followed the software's default procedures for variance estimation. For studies with multiple probiotic arms or other outcome measures, we used CMA's built-in procedures to avoid double-counting by combining arms into a single comparison or selecting the most relevant cognitive outcome as prespecified, ensuring that each study contributed only one effect size per meta-analysis. Continuous outcomes were pooled using standardized mean difference (SMD) with 95% confidence intervals (CIs). Because the included trials varied substantially in their methodological characteristics including differences in cognitive instruments, probiotic strains and formulations, dosages, intervention durations, and biomarker assay protocols, we used SMDs as the primary effect metric. The use of SMD allowed us to synthesize outcomes across heterogeneous study designs and measurement approaches in a statistically comparable manner, ensuring that the variability in protocols did not bias or limit the pooled estimates. Mean differences (MDs) were applied only in instances where studies used the same cognitive scale with identical scoring systems, such as MMSE raw scores; however, SMD was preferred for the main analyses to accommodate the diversity in tools, timing, and probiotic interventions across the RCTs. Heterogeneity was assessed using the I2 statistic, with I2 values of 25%, 50%, and 75% indicating low, moderate, and high heterogeneity, respectively. A random-effects model was used when significant heterogeneity (I2 > 50%) was present; otherwise, a fixed-effects model was applied. Publication bias was assessed using Egger's test. Subgroup analysis was conducted for the cognitively impaired population and based on probiotic type (single- or multi-strain).
Results
A complete search of PubMed, ProQuest, EBSCO, and Google Scholar revealed 3085 results. There were 2578 records left after 507 duplicates were eliminated. Upon screening these records, 2543 were excluded based on abstracts and titles. A full-text review was performed on 35 papers, yielding 19 research that met the inclusion criteria for qualitative synthesis and 16 studies for quantitative synthesis.22–40 PRISMA flow diagram (Figure 1) provides a full description of the research selection procedure.
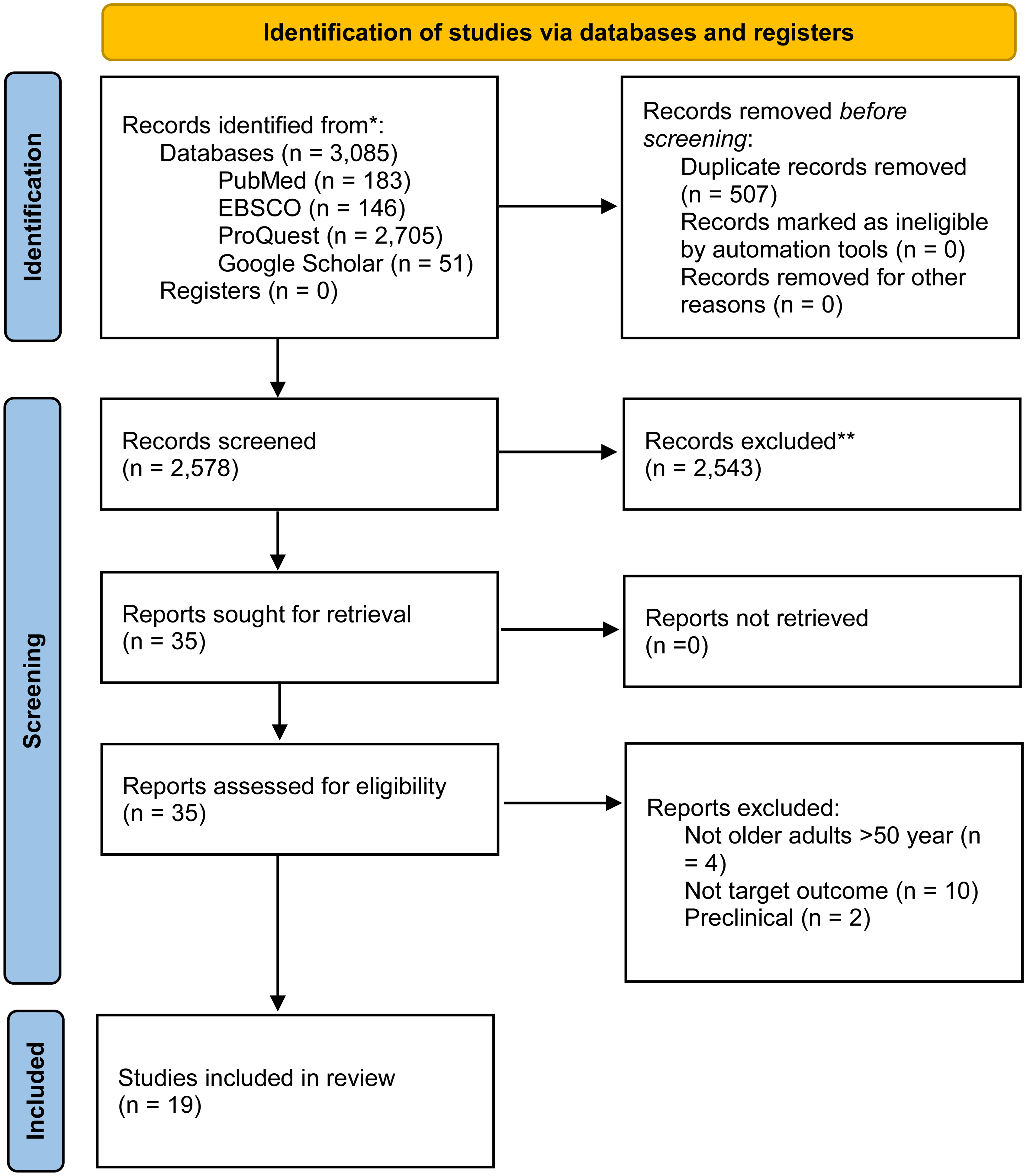
PRISMA flow diagram 2020.
Study characteristics
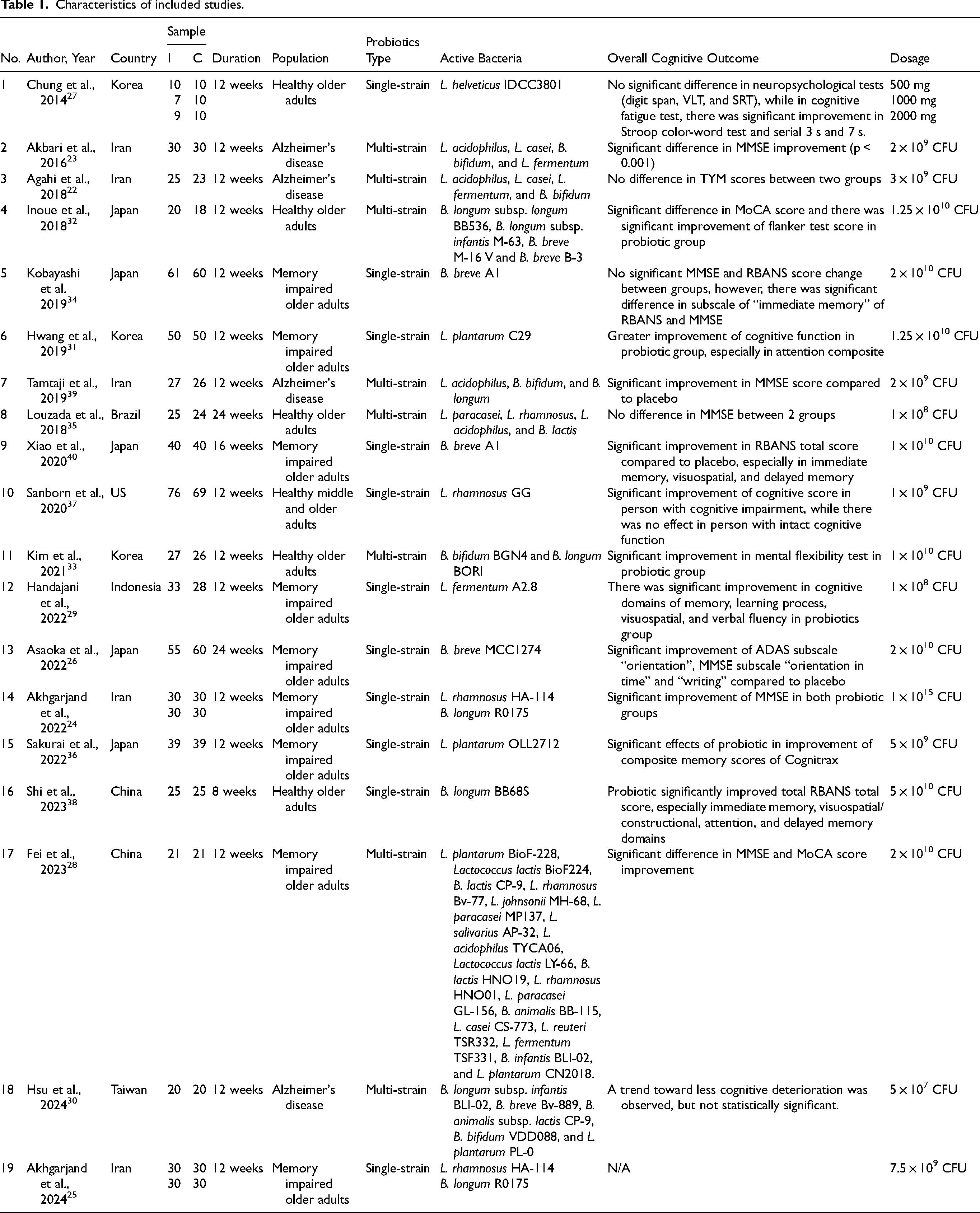
The included studies, which are listed in Table 1, were carried out from 2014 to 2024 in Korea, Iran, Japan, the US, Brazil, Indonesia, China, and Taiwan. Every study that was included had a randomized controlled trial design. The sample sizes ranged from 36 to 145, with participants aged 50 and older. All interventions lasted 8 to 24 weeks, mostly 12 weeks involving cognitively healthy older adults in 6 studies, while the other 13 studies involved cognitively impaired older adults. Probiotic supplementation was the main focus of the therapies, with a placebo serving as the comparator. Eight studies employed multi-strain probiotics while eleven studies used single-strain probiotics. Cognitive function was assessed mostly using MMSE, Alzheimer's Disease Assessment Scale-Cognitive Subscale (ADAS-Cog), and RBANS. Other relevant outcomes included BDNF, NO, TNF-alpha, 8-OHdG, IL-6, IL-10, MDA, TAC, GSH, and hs-CRP.
Characteristics of included studies.
Risk of bias assessment
The risk of bias was assessed using the Cochrane Risk of Bias Tool (RoB 2). Only 1 study has a high potential risk of bias, while others have a low risk or some were unclear (Supplemental Figure 1).
Overall results
The results of individual studies mostly yield positive results (Table 1). Sixteen studies showed significant improvement in cognitive function among participants receiving probiotics compared to placebo, while the other 3 studies found no significant improvement. However, some heterogeneities were noted, such as differences in population characteristics, type of bacteria, species of the bacteria, duration of study, and different outcome measurements. The meta-analysis included 16 studies (Table 2, Supplemental Material). The pooled effect size for the primary outcome, cognitive function measured using the MMSE, yielded an SMD of 0.747 (95% CI 0.307–1.186, I2 = 84.76%), indicating a significantly high improvement in the probiotics group
Meta-analysis of overall effects of probiotics in older adults.
Subgroup analysis
Further subgroup analysis focusing on cognitively impaired older adults revealed that only the MMSE yielded significant results, with an SMD of 0.761 (95% CI 0.269–1.253, I2 = 86.66%) (Table 3). In biochemical parameters, TNF-α, 8-OHdG, TAC, and GSH have the same results due to only including cognitively impaired older adults. IL-6 and IL-10 remained significant with SMD of −1.281 (95% CI −1.631–0.949, I2 = 0%) and 0.997 (95% CI 0.305–1.689, I2 = 81.77%), respectively. BDNF, NO, and hs-CRP serum levels were insignificant.
Meta-analysis of overall effects of probiotics in cognitively impaired older adults.

Another subgroup analysis, categorized based on probiotics type resulted significant result of improvement of MMSE score in the multi-strain subgroup (SMD of 0.878 [95% CI 0.107–1.648]), while there result of single-strain probiotics was insignificant (SDM of 0.653 [95% CI −0.023–1.329]) (Figure 2).

Forest plot of MMSE change between probiotic and control group based on probiotics type.
Meta-regression
Moderator analyses were performed to determine whether probiotic characteristics, dosage, probiotic formulation type, and intervention duration explained heterogeneity in cognitive outcomes in both the overall older adult population and the cognitively impaired subgroup (Supplemental Figure 2).
In the overall older adult population, none of the moderators significantly influenced MMSE effect sizes. High-dose probiotics did not produce greater cognitive improvement than standard doses (coefficient = −0.31, 95% CI −1.17–0.56, p = 0.489), and multistrain products were not superior to single-strain formulations (coefficient = 0.18, 95% CI −0.72–1.08, p = 0.697). Intervention duration also had no significant effect; the single 24-week study did not differ meaningfully from the 12-week trials (coefficient = −0.55, 95% CI −1.61–0.51, p = 0.307). Across all models, heterogeneity remained high (I2 ≈ 83–85%) and R2 analog values (0–5%) indicated minimal explanatory power.
Similar findings were observed in the cognitively impaired subgroup. Dosage again did not significantly influence cognitive outcomes (coefficient = −0.36, 95% CI −1.32–0.60, p = 0.463), and probiotic formulation type was not a meaningful moderator (coefficient = 0.22, 95% CI −0.79–1.23, p = 0.667), explaining none of the between-study variance (R2 = 0%). Duration could not be reliably analyzed as a moderator because only one study included a 24-week intervention, while all others used 12-week protocols.
Microbiome findings and mechanistic alignment
Six included RCTs reported microbiome-adjacent endpoints, including fecal 16S rRNA profiles and/or gut-derived metabolites. Across these studies, probiotics generally produced directional shifts toward anti-inflammatory or gut-beneficial taxa, with several trials demonstrating parallel improvements in cognitive outcomes.
Probiotic supplementation resulted in no major changes in alpha diversity in studies by Kim et al., Sakurai et al. (no detectable increase in overall richness), and Asaoka et al. (global diversity largely preserved). Beta-diversity (Bray–Curtis or UniFrac) showed modest but statistically significant community shifts in Kim et al. and Shi et al., indicating that probiotics altered global microbiome structure even without increasing richness. These compositional shifts corresponded with changes in key functional taxa.
Across the trials that reported taxonomic outcomes, probiotics consistently shifted the gut microbiota toward genera associated with anti-inflammatory or SCFA-related functions. In Kim et al., supplementation reduced several inflammation-associated taxa while increasing Eubacterium and members of Clostridiales changes that correlated negatively with serum BDNF only in the probiotic group, suggesting a link between microbial modulation and neurotrophic signaling. Sakurai et al. similarly observed reductions in Lachnoclostridium, Monoglobus, and Oscillibacter, genera implicated in gut inflammation, accompanied by improvements in memory scores. In Shi et al., probiotics enriched multiple butyrate-producing genera, including Lachnospira, Dorea, and Bifidobacterium, while reducing taxa previously associated with cognitive impairment such as Collinsella, Parabacteroides, Tyzzerella, Bilophila, and Granulicatella, paralleling substantial RBANS improvements. In contrast, Asaoka et al. reported that overall microbial composition remained relatively stable despite cognitive improvements and attenuated brain-atrophy progression, suggesting that some probiotic effects may act through metabolic or immune pathways rather than large-scale taxonomic restructuring. Direct SCFA measurements were available only in a subset of trials (Shi et al., some partial in Kim et al.). Where reported, probiotics led to increases in acetate/propionate/butyrate-associated genera, consistent with improved fermentative capacity even when SCFA concentrations were not directly quantified. None of the trials reported significant changes in LPS, zonulin, or lactulose–mannitol permeability; most did not measure these endpoints.
Publication bias
Publication bias was assessed using Egger's test. The Egger's test indicated the presence of publication bias in MMSE, MDA, and GSH outcome measures.
GRADE summary
The certainty of evidence for the primary cognitive outcome, MMSE, was rated as moderate. Although the pooled effect suggested a modest improvement with probiotics, the certainty was downgraded for inconsistency due to substantial heterogeneity across trials (Supplemental Table 2). However, the body of evidence included multiple randomized trials with generally low-to-moderate risk of bias, so no further downgrading was applied. The certainty for RBANS was assessed as low to moderate, downgraded for inconsistency and imprecision because confidence intervals were wide, and the number of eligible trials was smaller than for MMSE, limiting the precision of the estimate.
Discussion
The current study demonstrates that probiotics were associated with improvements in cognitive function, BDNF levels, and various biochemical markers, including 8-OHdG, IL-6, IL-10, MDA, TAC, and GSH, in older adults. However, these findings should be interpreted deliberately, as the observed biomarker changes are consistent with proposed mechanistic pathways, but do not establish that improvements in cognition were directly mediated through BDNF, inflammation or oxidative stress. Notably, BDNF levels significantly increased in the probiotic group compared to the placebo group. Pro-inflammatory markers like IL-6 and TNF-α decreased, while the anti-inflammatory cytokine IL-10 showed a significant increase. Additionally, oxidative stress markers (MDA, TAC, and GSH) indicated enhanced antioxidant activity. In further subgroup analysis, improvements in cognitive function, TNF-α, IL-6, IL-10, 8-OHdG, TAC, and GSH remained significant, while changes in BDNF were not.
The gut microbiota, which is mostly composed of more than 1014 bacteria, is a dynamic and diverse collection of microorganisms that inhabit the human gastrointestinal tract. 41 This microbiota affects both healthy physiological processes and disease states, which is why it is so important to the host's overall health. Because of its distinct cellular, molecular, and architectural features, the intestine offers the perfect environment for good bacteria to proliferate and carry out their health-improving functions.42,43 The gut microbiota's composition varies greatly depending on the host and is impacted by several factors like nutrition, probiotic exposure, intestinal environment, and other associated factors. 43 Probiotics, which can be sourced from fermented foods like sauerkraut, pickles, yogurt, and kefir, have a beneficial impact on the human gut microbiome. 42 They work through several mechanisms, including enhancing the mucosal barrier, directly antagonizing pathogens, inhibiting bacterial adherence and invasion in the intestinal epithelium, boosting the immune system, and regulating the central nervous system. 44 The main genera of probiotic microorganisms used in human nutrition include Lactobacillus, Bifidobacterium, Lactococcus, Streptococcus, and Enterococcus, with some strains of Bacillus and Saccharomyces also being utilized. 42
BDNF is a protein that is essential for neuronal growth, development, and maintenance. 11 It belongs to the family of growth factors known as neurotrophins, which are crucial for sustaining adult brain neuronal plasticity and function as well as for the survival and differentiation of neurons throughout development. Increased levels of BDNF have been associated with better metabolic and mental health. 45 The effects of using polyphenols, fish oil, vitamins, probiotics, and other compounds to increase BDNF levels have been studied extensively, but the outcomes have been inconsistent.11,46 In the current study, BDNF levels rose dramatically in older persons as an overall group, but not significantly in those who were only cognitively impaired. A meta-analysis by Deghani et al. that includes various mental health conditions also showed no significant BDNF change overall following probiotics supplementation; however, BDNF increase was found to be significant in individuals with neurological disorder and depression. 47 BDNF was found to be increased after supplementation of probiotics compared to baseline; however, no significant difference was found between the placebo and intervention group in RCT involving major depressive disorder patients conducted by Schneider et al. 48 The inconsistent BDNF change might be caused by concomitant treatment, heterogeneity in the BNDF baseline, and measurement methods. The measurement instruments and peripheral BDNF concentration (in plasma or serum) also had an impact on the relevance of BDNF levels. Whether peripheral BDNF concentration correctly represents central BDNF concentration is still unknown. Dietary intake also might affect BDNF levels. 11 There were only 4 studies that were analyzed with only 2 that included cognitively impaired older adults. Hsu et al. used multistrain probiotics showed a significant increase while Hwang et al. who used single-strain probiotics showed no difference in BDNF change.31,49 In healthy older adults, research by Chung et al. who used single-strain probiotics and Kim et al. who used multistrain probiotics, both showed significant changes in BDNF.27,33 Future research should be done to investigate if probiotic type affects the effect of BDNF increase.
Gut dysbiosis in AD patients altered the relative abundance of key butyrate-producing bacteria such as Bacteroidaceae, Veillonellaceae, and Lachnospiraceae, reduction of Gram-negative bacteria, and increased intestinal permeability.52–54 Butyrate is a type of short-chain fatty acid (SFCA), which has anti-inflammatory and neuroactive effects and also has a role in maintaining barrier integrity and neuroimmune mechanisms.55–57 Intestinal microbial infection might also play a role in AD pathogenesis. Cognitively impaired elderly patients with amyloidosis have shown a higher relative abundance of proinflammatory microbial species compared to controls. 58 Infection and inflammation might also accelerate Aβ plaque deposition. 59 Aging population and older adults with neurodegenerative disease were also found to have higher IL-2, IL-4, IL-6, IL-8, IL-10, INF-γ, and TNF-α levels in their cerebrospinal fluid.14,15,60 Pro- and anti-inflammatory cytokines play a crucial role in the development of Aβ plaques in the brains of AD patients. In the aging brain, a combination of cytokines including IL-1β, IL-6, IL-8, IL-10, IL-12, and TNF-α trigger inflammatory processes linked to cortical atrophy. 61 Although probiotics significantly decrease inflammatory cytokines in the current analysis, this does not confirm that cytokine changes directly mediate improvements in cognition, rather, they may represent parallel but related physiological changes.
This study demonstrates that probiotics significantly decrease inflammatory cytokines in older adults, which is consistent with prior research. 50 A meta-analysis done by Milajerdi et al. involving 42 RCTs in various health conditions showed that probiotics significantly decreased pro-inflammatory cytokines (hs-CRP, TNF-α, IL-6, IL-12, and IL-4) with increased anti-inflammatory cytokine (IL-10). 50 Probiotics can modulate JAK/STAT signaling and NF-κB pathways to affect inflammation and cytokine production. 51 A study by Petrella et al. showed that prolonged multi-strain probiotics supplementation prevents the increase of LPS-dependent inflammation in the mouse's hippocampus and cortex and triggers pro-neurogenic response enhancing hippocampal neurogenesis, in addition to improving gastrointestinal epithelial barrier. 62 B. breve supplementation significantly decreased Aβ and reduced mRNA expression of proinflammatory cytokine in the brain through activation of the ERK/HIF-1α signaling pathway. 63 IL-10, an anti-inflammatory cytokine, has multiple immunoregulatory effects, especially important during the resolution phase of inflammation. 60 In this meta-analysis, IL-10 was found to increase significantly in the probiotics group, highlighting improved anti-inflammatory reaction. IL-10 inhibits the production of several inflammatory cytokines such as TNF-α, IL-1β, IL-6, and IFN-γ from Toll-Like Receptor (TLR)-activated myeloid lineage cells. 60 However, increased IL-10 does not necessarily indicate that cognitive improvements were directly driven by these changes, as causal pathways cannot be inferred from peripheral biomarkers alone.
Oxidative stress, defined as an imbalance between oxidants and antioxidants leading to elevated oxidant levels, is a recognized clinical marker of AD. Reactive oxygen species (ROS) and Aβ are key mediators of oxidative stress. Fracassi et al. found dramatic redox imbalance, associated with impaired antioxidant defenses in the AD brain. By contrast, non-demented individuals display low oxidative damage, which is associated with high levels of scavenging systems, possibly resulting from a lack of PGC1a miRNA-485-related inhibition. 16 This study found improved anti-oxidant capacity in the probiotics group, which may align with hypothesized mechanisms of cognitive protection, though direct mediation cannot be confirmed. Increased serum antioxidant vitamins (vitamins A, C, and E) and carotenoids significantly reduced the risk of all-cause dementia. Besides probiotics, polyphenols such as resveratrol, rosmarinic acid, curcuminoids, and anthocyanins also have anti-oxidant activity and are proposed to prevent dementia. 64 Immune cells create nitric oxide (NO) by expressing the iNOS gene, which is triggered by microbial substrates and different cytokines. NO possesses several important properties, including immunosuppressive, tissue-damaging, antibacterial, and anticancer actions. 65 But because NO is a reactive radical, it can combine with anion radicals to create peroxynitrite, a strong oxidant that can cause inflammation. Reducing NO generation is therefore thought to be an efficient therapy approach to lessen these negative consequences, however, in this study, NO levels did not change after probiotic supplementation. In the Kim et al. study, lactic acid bacteria treatment can relieve oxidative stress by scavenging DPPH and ABTS radicals and suppressing NO production. 65 The mechanisms underlying the antioxidant activity of probiotics are unclear. It is proposed that lactic acid bacteria contribute to antioxidant effects through several pathways: scavenging ROS, metal chelation, enhancing the levels of antioxidant enzymes, and altering the gut microbiota. 66
Across trials reporting fecal microbiome outcomes, probiotics consistently produced anti-inflammatory or SCFA-related microbial shifts and, in several cases, these aligned with cognitive benefits. Although alpha diversity changes were minimal, probiotics altered community composition and enriched taxa linked to butyrate production (Lachnospira, Dorea, Eubacterium), while reducing genera associated with inflammation or cognitive impairment (Collinsella, Tyzzerella, Bilophila). Notably, in Kim et al., taxa altered by supplementation showed significant correlations with BDNF levels, and in Shi et al., cognitive improvement paralleled enrichment of beneficial taxa. These findings collectively support a mechanistic model in which probiotics improve cognition through modulation of gut microbial composition, SCFA-related metabolic pathways, and downstream neuroimmune signaling, even in the absence of large shifts in alpha diversity.
Although probiotics were associated with cognitive improvement in both the overall older adult population and in cognitively impaired participants, these effects should not be generalized to all older adults. The subgroup analysis showed only significant changes in multi-strain studies. The effect of multi- and single-strain probiotics has never been investigated before for the cognitive function of older adults. However, there was some research investigating these differences in another medical condition.67–69 It is hypothesized that multi-strain probiotics had optimal effects compared to single-strain and more consistent results in gastrointestinal diseases and pathogen inhibition. A study by Fijan et al. showed that multi-strain probiotics showed statistically significant higher antagonistic effects of wound pathogens compared to single-strain microbes. 67 Multi-strain microbes are said to be better because they offer several advantages over single-strain microbes. These advantages include enhanced efficacy, diverse functionality, increased resilience, complementary actions, and adaptability. In another review by McFarland et al., involving a total of 65 RCTs, multi-strain probiotics were not significantly more effective than single-strain probiotics. 69 Although several trials reported changes in neurotrophic, inflammatory, and oxidative stress biomarkers, these findings should not be interpreted as evidence that probiotics directly improve cognition through these pathways. The biomarker results are best viewed as signals that align with proposed microbiota–gut–brain mechanisms, rather than confirmation of causal mediation.
Limitations
Several important limitations should be acknowledged. Many included trials were of short duration, typically 8–12 weeks, which restricts conclusions about the sustainability of cognitive effects. Although several studies reported changes in biomarkers such as BDNF, cytokines, or oxidative stress markers, these were peripheral measurements that may not accurately reflect central neurobiological processes, and their interpretation is further limited by heterogeneity in assay methods, biospecimen types (serum versus plasma), and unit reporting. Variability in probiotic strains, combinations, dosages, and delivery formats also complicates comparisons across trials and limits the ability to identify which formulations, if any, are most effective. The evidence base for many biomarkers remained extremely small, with only two or three studies contributing to certain outcomes such as 8-OHdG, TAC, and GSH, resulting in statistically fragile estimates that should be viewed as exploratory. Additionally, the literature search was constrained by institutional access, which prevented the use of Embase and CENTRAL; although this was mitigated through extensive manual searching and registry screening, the possibility of missed studies cannot be entirely excluded. Taken together, these limitations indicate that the findings, particularly for mechanistic biomarkers—should be interpreted cautiously and underscore the need for larger, longer-duration, and methodologically standardized trials.
Conclusion
Probiotic supplementation was associated with modest improvements in cognitive outcomes among older adults, with more consistent signals observed in individuals with mild cognitive impairment or subjective cognitive decline. However, these findings should be interpreted cautiously due to substantial between-study heterogeneity, the generally short duration of interventions, and the limited number of studies assessing mechanistic biomarkers, many of which employed heterogeneous methodologies. The current evidence is insufficient to draw firm conclusions regarding specific probiotic formulations, optimal dosages, or underlying biological pathways. Rather than supporting routine clinical use, these results suggest that probiotics, particularly multistrain formulations, represent a promising area for further investigation in cognitively at-risk populations. Well-powered, longer-term, and methodologically standardized randomized controlled trials, including those explicitly designed to evaluate mechanistic mediation, are required to clarify the robustness, clinical relevance, and biological basis of probiotic effects on cognitive aging.
Supplemental Material
sj-docx-1-alz-10.1177_13872877261415596 - Supplemental material for Probiotics and aging: Impacts on cognitive function, brain-derived neurotrophic factor, inflammation, and antioxidant activity. A systematic review and meta-analysis
Supplemental material, sj-docx-1-alz-10.1177_13872877261415596 for Probiotics and aging: Impacts on cognitive function, brain-derived neurotrophic factor, inflammation, and antioxidant activity. A systematic review and meta-analysis by Yvonne Suzy Handajani, Yuda Turana, Antoninus Hengky and Kevin Kristian in Journal of Alzheimer's Disease
Footnotes
Acknowledgements
The authors have no acknowledgments to report.
Ethical considerations
Not applicable
Consent to participate
Not applicable
Consent for publication
Not applicable
Author contribution(s)
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
The data supporting the findings of this study are available on request from the corresponding author.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.