
Abstract
Background
Epigenetic age acceleration (EAA) refers to the extent to which an individual's biological age, estimated from DNA methylation patterns, exceeds their chronological age, indicating accelerated cellular and tissue aging.
Objective
We investigated the association between EAA and Alzheimer's disease (AD), with a focus on sex-based differences.
Methods
EAA was estimated from blood samples in 127 participants with Alzheimer's disease-related cognitive impairment (ADCI) and 143 cognitively unimpaired (CU) participants, recruited from a nationwide multicenter study under the Precision Medicine Platform for Mild Cognitive Impairment (PREMIER) consortium in Korea.
Results
EAA measures indicated higher acceleration in the ADCI group compared to the CU group, particularly for extrinsic epigenetic age acceleration (EEAA), AgeAccelResidualHannum, and AgeAccelPheno. Sex-specific analyses revealed that EEAA significantly differed between the ADCI and CU groups in both men and women, with a greater EEAA in men. Logistic regression analysis demonstrated that increased EEAA, the presence of APOE ɛ4 allele, and poorer nutritional status were significantly associated with a higher likelihood of ADCI. EEAA increased ADCI risk more strongly in men than in women, whereas chronological age showed a protective effect only in women.
Conclusions
As a marker of immune system aging, EEAA may be associated with ADCI. These findings suggest that EEAA serves as a complementary indicator of systemic biological aging within the AD spectrum. The greater EAA observed in men was consistently present in ADCI, highlighting the importance of considering sex differences in EAA-related AD research.
Introduction
Alzheimer's disease (AD) is a leading cause of dementia, accounting for 60%–70% of cases and remains a major public health concern. Its pathological features include amyloid plaques and neurofibrillary tangles, which result from the accumulation of abnormal amyloid-β (Aβ) and tau proteins in the brain. 1 Despite extensive research, the mechanisms underlying AD pathogenesis remain unclear, necessitating further investigation into its biological foundations.
The epigenetic clock, derived from DNA methylation patterns, 2 provides a novel framework for understanding AD pathogenesis. By offering insights into biological aging, the epigenetic clock may help clarify the intricate mechanisms driving AD development, highlighting the interplay between aging, epigenetics, and the pathological features of the disease. Although chronological age, the time elapsed since birth, is the primary risk factor for AD, 3 it does not account for biological heterogeneity among individuals. Epigenetic age acceleration (EAA) refers to a phenomenon in which biological age advances more rapidly than chronological age, as indicated by DNA methylation patterns, reflecting the accelerated aging of cells and tissues. 2 To quantify this phenomenon, epigenetic clocks estimate biological age by modeling methylation levels at specific age-associated CpG sites, and EAA is typically calculated as the residual difference between methylation-predicted biological age and chronological age. 4 Given that accelerated biological aging has been suggested as a potential factor influencing various age-related conditions, including AD, dementia, and cognitive decline, 2 understanding this discrepancy could be essential for predicting AD risk.
Previous studies have reported inconsistent findings regarding the relationship between epigenetic clocks and AD. EAA in brain tissue has been correlated with AD pathology, whereas its association with cerebrovascular neuropathology appears less pronounced.5,6 Dunedin Pace of Aging Calculated from the Epigenome (DunedinPACE), a method developed as part of the Dunedin Study to estimate the pace of aging, has been associated with mild cognitive impairment (MCI), AD, and lower cognitive test scores. 7 Additionally, epigenetic clocks in mouse brain regions have indicated accelerated aging in an AD model, particularly in the cortex. 8 Furthermore, Hannum's clock is a DNA methylation-based estimator of biological age derived from blood markers and has been linked to hippocampal volume in AD. 9 However, some studies have reported that blood-based EAA does not predict dementia risk 10 and is not associated with cognitive status or amyloid positivity.9,11
Sex is another established risk factor for AD, with a higher prevalence in women. 12 In cognitively healthy individuals, women exhibit worse AD biomarkers than men, including greater amyloid deposition on amyloid positron emission tomography (PET), lower glucose metabolism, and reduced gray and white matter volumes on magnetic resonance imaging (MRI). 13 Sex disparities in AD risk factors, including classic cardiovascular risk factors, major depression, and the Apolipoprotein E (APOE) ɛ4 genotype, have been documented.14–17 This discrepancy is attributed to multiple factors, including biological differences (e.g., postmenopausal hormonal changes), genetic predispositions, and lifestyle influences. 18
Recent studies suggest sex-based differences in EAA, with men generally exhibiting a greater degree of biological aging than women.19,20 Factors such as body mass index (BMI) and smoking partially mediate this effect, particularly in older individuals. 21 Studies from Mediterranean Blue Zones indicate that men exhibit higher EAA values across various epigenetic clocks (e.g., Horvath, Hannum, and GrimAge) than women. 22 These findings suggest that sex directly influences biological aging, with differences becoming more pronounced with age. 21 This body of evidence supports the hypothesis that sex-based differences in EAA contribute to AD risk. Understanding these differences may be crucial for elucidating the mechanisms underlying sex-related disparities in AD onset and progression.
Despite the high prevalence of AD in women and the potential influence of sex on AD onset and progression, the role of sex-specific EAA in AD remains unclear. Investigating sex-based differences in EAA could provide new perspectives for developing personalized diagnostic tools and treatments. The aims of this study were to examine the association between blood-based EAA measures and AD, identify factors influencing EAA, and to investigate sex-based differences in the relationship between EAA measures and AD.
Methods
Participants
Participants were recruited from a nationwide multicenter study conducted in collaboration with the Precision Medicine Platform for Mild Cognitive Impairment, based on the Multi-omics, Imaging, Evidence-based R&BD (PREMIER) consortium in Korea. The PREMIER consortium included patients with AD dementia, MCI, subjective cognitive decline (SCD), and cognitively normal controls (NC). While participating in the PREMIER study, all participants underwent clinical interviews, neurological and neuropsychological examinations, questionnaires, blood sample collection, MRI, and amyloid PET. 23 Exclusion criteria for PREMIER included a current or past history of neurological or psychiatric illnesses, such as schizophrenia, bipolar disorder, epilepsy, brain tumors, encephalitis, extensive white matter changes, hearing or vision loss, severe cardiac or respiratory disorders, malignancy, hepatic or renal disorders, or severe head trauma. A total of 127 patients with AD-related cognitive impairment (ADCI) and 143 cognitively unimpaired (CU) participants were analyzed. Participants were recruited from 16 centers between March 2020 and January 2021. The CU group included amyloid-negative NC and amyloid-negative SCD. The ADCI group comprised 37 patients with amnestic MCI (aMCI) and 90 patients with AD dementia, all of whom were amyloid-positive. The original study (PREMIER consortium) was approved by the Institutional Review Board (IRB) of Samsung Medical Center, Seoul, Republic of Korea (IRB No. 2020-01-024). This study was exempted from review by the IRB of Pusan National University (PNU IRB/2022_93_HR).
Clinical diagnosis
Patients with aMCI met both the National Institute on Aging and Alzheimer's Association (NIA-AA) core clinical criteria for MCI 24 and the modified Petersen's criteria, 25 which include: 1) subjective memory complaints reported by either the patient or a caregiver; 2) normal activities of daily living (ADL), based on clinical findings and the Korean Instrumental Activities of Daily Living Scale (K-IADL) 26 ; 3) objective memory dysfunction, defined as a Z score of −1.0 standard deviations (SD) or lower relative to age- and education-matched normal individuals; and 4) the absence of dementia. Patients with AD dementia met the NIA-AA core clinical criteria for probable AD dementia. 27 SCD was defined as subjective memory complaints reported by either the patient or a caregiver, with no objective cognitive dysfunction based on cognitive domain evaluations. The criteria for NC were: 1) absence of cognitive complaints, and 2) no objective cognitive dysfunction based on cognitive domain evaluations.
Neuropsychological tests
General cognitive function was assessed in all participants using the Korean version of the Mini-Mental State Examination (K-MMSE). 28 All participants underwent comprehensive neuropsychological assessments using the Seoul Neuropsychological Screening Battery (SNSB) for diagnostic evaluation.29,30 The SNSB comprises tests of language (Korean version of the Boston Naming Test and the Controlled Oral Word Association Test), visuospatial function (Rey Complex Figure Test [RCFT]), verbal memory (Seoul Verbal Learning Test: immediate recall, delayed recall, and recognition), visual memory (RCFT: immediate recall, delayed recall, and recognition), attention and executive function (Digit Span Forward and Backward tests and the Trail Making Test parts A and B). 29
Questionnaire
Alcohol consumption and smoking status were assessed regardless of time. The Mini Nutritional Assessment was conducted, and BMI was calculated using measured height and weight to classify nutritional status as normal, at risk of malnutrition, or malnourished. 31 The Korean Version of the Short Form Geriatric Depression Scale (SGDS-K) was used, with scores of 8 or higher indicating a depressive state. 32 The Korean version of the International Physical Activity Questionnaire (IPAQ) was used to calculate MET scores. These scores were determined by multiplying the MET value assigned to each activity (8.0 for vigorous, 4.0 for moderate, and 3.3 for walking) by the duration of the activity per day and the number of days per week. 33 The total MET score was obtained by summing the MET-min/week values for all activity types.
Amyloid PET scan
18F-florbetaben (n = 71) or 18F-flutemetamol (n = 199) was injected intravenously, and PET images were acquired 90–110 min post-injection. PET images were visually assessed by a nuclear physician blinded to clinical data using a previously described method.34,35 Images were assigned either a positive or negative score based on intensity comparisons between the cortical gray matter and adjacent white matter. Aβ negativity was defined as lower tracer uptake in the gray matter than in the adjacent white matter, whereas Aβ positivity was defined as equal or greater tracer uptake in the gray matter than in the white matter.
Apolipoprotein E genotype
APOE genotyping was performed using real-time polymerase chain reaction with the Real-Q Apo E genotyping kit (BioSewoom, Seoul, Korea) following the manufacturer's instructions.
DNA methylation analysis
Genomic DNA extracted from blood samples was treated with bisulfite using an EZ DNA Methylation Kit (Zymo Research, USA). DNA was amplified via isothermal multiple displacement amplification (IMDA), fragmented, purified, and hybridized. DNA methylation analysis was performed using the Infinium MethylationEPIC BeadChip (Illumina, CA, USA) following the manufacturer's instructions. A researcher blinded to clinical data conducted the analysis. Fluorescence images were obtained from BeadChips using the iScan™ System (Illumina, CA, USA). Methylation levels at interrogated loci were analyzed by calculating the ratio of fluorescent signals from methylated and unmethylated sites.
DNA methylation data were analyzed using the R package ChAMP. 36 We excluded samples with high detection p-values (>0.05 in more than 1% of probes) and normalized the data using the beta-mixture quantile (BMIQ) method to correct for probe-type bias. Batch effects were adjusted using ComBat, a Bayesian framework for estimating and correcting batch effects. 37
In addition to ChAMP's default filtering, we removed probes with low beadcount (<3 beads in >5% of samples), probes that map to multiple genomic locations (cross-hybridizing), and probes containing SNPs at the CpG site or single base extension. We also excluded probes located on sex chromosomes to avoid sex-specific methylation bias. Sample-level quality was further assessed using signal intensity distributions and principal component analysis to detect outliers. Finally, we inferred biological sex from methylation data and confirmed concordance with reported sex information to exclude potential sample mismatches.
Epigenetic age acceleration estimation
EAA was calculated using the HorvathAge (353 CpGs), HannumAge (71 CpGs), PhenoAge (513 CpGs), and GrimAge (1030 CpGs) algorithms.4,13,38,39 To estimate these values, the sample annotation file and β-values, which reflect methylation levels ranging from 0 to 1 at each CpG site, were uploaded to the online DNA Methylation Age Calculator (https://dnamage.clockfoundation.org). The output included multiple EAA measures: AgeAccelResidual, intrinsic epigenetic age acceleration (IEAA), EEAA, AgeAccelResidualHannum, AgeAccelPheno, and AgeAccelGrim. AgeAccelResidual, IEAA, and EEAA were calculated as residuals by regressing DNAm age, estimated using the Horvath algorithm, on chronological age. The same method was applied to predict AgeAccelResidualHannum, AgeAccelPheno, and AgeAccelGrim.
The DunedinPACE (173 CpGs) is a next-generation method for tracking an individual's rate of physical aging by analyzing methylation patterns. 40 This algorithm assessed 19 biomarkers associated with age-related changes in individuals of the same chronological age over a 20-year period and identified methylation patterns predictive of the rate of aging. DunedinPACE results can be interpreted in reference to an average biological aging rate of one year per year of chronological aging. DunedinPACE was calculated using an R package available at https://github.com/danbelsky/DunedinPACE.
Statistical analysis
All statistical analyses were performed using SPSS (version 25.0; IBM SPSS Statistics, Armonk, NY, USA) and R statistical software (version 4.2.2; The R Foundation for Statistical Computing, Vienna, Austria). A p-value < 0.05 was considered statistically significant. Continuous variables are presented as mean ± SD, while categorical variables are expressed as numbers and percentages. Demographic and clinical characteristics were analyzed using the chi-square test or independent t-test. Differences in EAA measures between groups were evaluated using analysis of covariance (ANCOVA), adjusting for potential confounders identified during baseline characterization. False discovery rate (FDR) correction using the Benjamini–Hochberg procedure was applied to control for multiple comparisons among EAA measures. A linear regression analysis was performed to identify factors influencing EAA, which consistently differed between the CU and ADCI groups across the overall sample and within sex-specific analyses. To control for confounding, linear regression model was adjusted for established demographic factors (e.g., chronological age, sex), lifestyle habits (drinking and smoking), and clinical variables that showed significant inter-group differences. Logistic regression analysis was also conducted to predict ADCI, with the models adjusted for clinical and demographic covariates that exhibited significant baseline differences between the CU and ADCI groups.
Results
Demographic and clinical characteristics
Table 1 presents the demographic and clinical characteristics of participants, categorized into the CU and ADCI groups and further stratified by sex. Participants in the ADCI group had a lower BMI (22.76 ± 2.79, p = 0.001) and MMSE scores (20.90 ± 5.01, p < 0.001), a lower proportion with normal nutritional status (24.8%, p < 0.001), and a higher prevalence of APOE ɛ4 carrier status (58.7%, p < 0.001) compared with those in the CU group. These trends were consistent in both women and men.
Demographic characteristics of participants by groups, stratified by sex.

CU: cognitively unimpaired; ADCI: Alzheimer's disease-related cognitive impairment; M: mean; SD: standard deviation; BMI: body mass index; APOE: Apolipoprotein E; MMSE: Mini-Mental State Examination; MET: metabolic equivalent task; CRP: C-reactive protein.
The differences between the CU and ADCI groups by sex were as follows: Women in the ADCI group had fewer years of education (6.85 ± 2.21, p = 0.012), a lower incidence of drinking experience (22.2%, p = 0.031), and a higher prevalence of depression (27.2%, p = 0.021) than those in the CU group. Men in the ADCI group had lower physical activity levels (2261.93 ± 2655.32, p = 0.047) than those in the CU group. No significant differences were observed in chronological age, smoking experience, or hs-CRP levels between the CU and ADCI groups in either women or men.
Sex differences in epigenetic age acceleration
Table 2 presents the results of an ANCOVA comparing EAA measures between the CU and ADCI groups, stratified by sex. The analysis adjusted for relevant covariates—including BMI, education, APOE status, nutritional status, and depression—that showed significant differences between the CU and ADCI groups in Table 1. Participants in the ADCI group exhibited greater acceleration in EEAA (1.30 ± 4.51, p < 0.001; FDR-adjusted p = 0.014), AgeAccelResidualHannum (0.69 ± 3.49, p = 0.002; FDR-adjusted p = 0.014), and AgeAccelPheno (0.85 ± 4.54, p = 0.011; FDR-adjusted p = 0.042). Women in the ADCI group exhibited greater acceleration in EEAA (p = 0.012; FDR-adjusted p = 0.042) than those in the CU group. In men, EEAA (p = 0.002; FDR-adjusted p = 0.014) and AgeAccelResidualHannum (p = 0.012; FDR-adjusted p = 0.042) were significantly greater in the ADCI group than in the CU group. EEAA was significantly accelerated in both women and men in the ADCI group, with men showing a significantly greater acceleration (3.40 ± 3.99) than women (0.06 ± 4.36) (Table 2, Supplemental Table 1, and Supplemental Figure 1). Detailed sex-specific subgroup analyses across diagnostic categories (CU, MCI, and AD dementia) are presented in Supplemental Table 2. Both EEAA and AgeAccelResidualHannum were significantly higher in the AD dementia group compared to the CU group. In men, EEAA was significantly higher in both the MCI and AD dementia groups compared to the CU group (Supplemental Table 2). Sex-specific associations of EAA measures are provided in Supplemental Table 3. Additionally, Supplemental Figure 2 illustrates the pairwise correlations between chronological age and epigenetic age measures, including regression lines and Pearson correlation coefficients, presented separately for all participants, men, and women.
Analysis of covariance results comparing epigenetic age acceleration measures between groups, stratified by sex.

Differences in epigenetic age acceleration measures between groups were analyzed using analysis of covariance, adjusting for BMI, education, APOE status, nutritional status, and depression. False discovery rate correction was applied to account for multiple comparisons. The analysis included 227 participants because of missing data on BMI and nutritional state responses.
CU: cognitively unimpaired; ADCI: Alzheimer's disease-related cognitive impairment; M: mean; SD: standard deviation; F: F-statistic; FDR: false discovery rate; IEAA: intrinsic epigenetic age acceleration; EEAA: extrinsic epigenetic age acceleration; DunedinPACE: pace of aging calculated from the epigenome.
Factors influencing extrinsic epigenetic age acceleration
Since EEAA consistently differed between the CU and ADCI groups across the overall sample and within sex-specific analyses (Table 2), we considered EEAA to be a suitable and generalizable measure for identifying factors influencing EAA in this study. Female sex and ADCI diagnosis were significantly associated with EEAA (Table 3). Women exhibited significantly lower EEAA than men (β = −0.27, p = 0.003), whereas participants with an ADCI diagnosis had significantly higher EEAA (β = 0.31, p < 0.001). Other factors, including chronological age, BMI, drinking, smoking, APOE ɛ4 carrier status, nutritional status, depression, and physical activity, did not significantly affect EEAA. The model explained 22.2% of the variance in EEAA (adjusted R2 = 0.18, F = 5.56, p < 0.001), highlighting the substantial impact of sex and ADCI diagnosis on epigenetic aging.
Factors associated with extrinsic epigenetic age acceleration (n = 206).
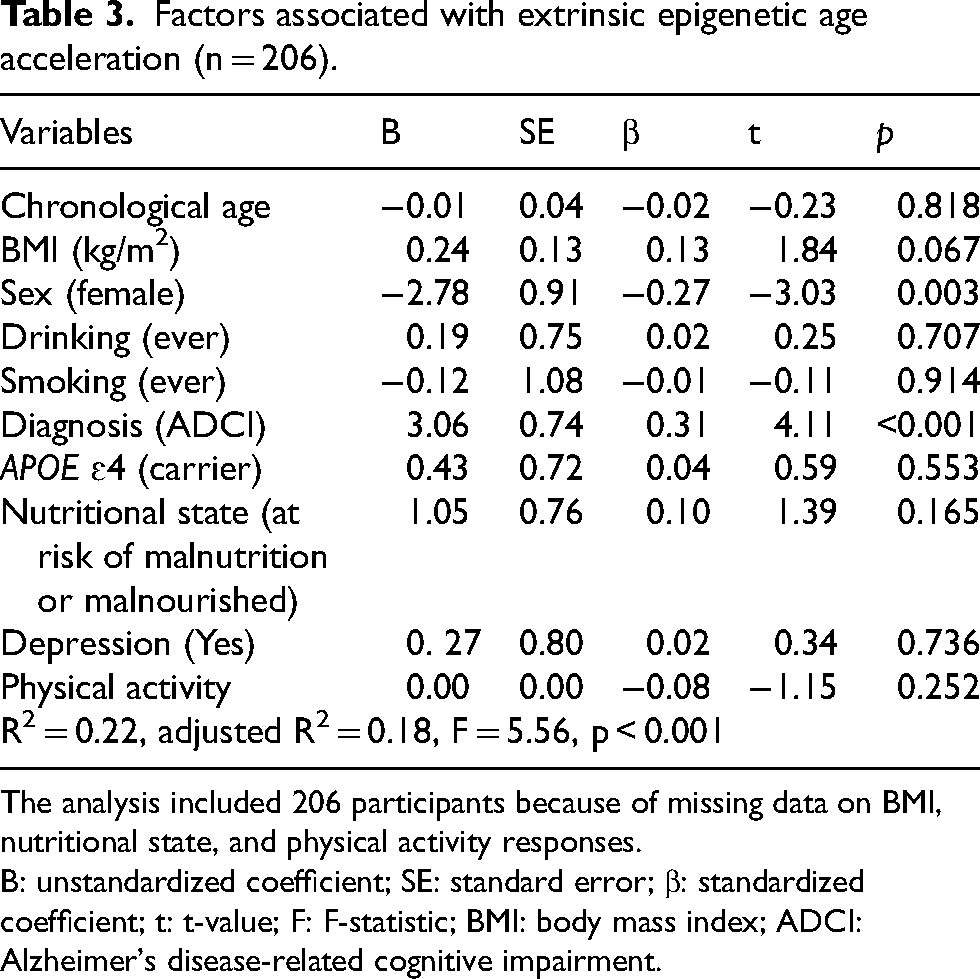
The analysis included 206 participants because of missing data on BMI, nutritional state, and physical activity responses.
B: unstandardized coefficient; SE: standard error; β: standardized coefficient; t: t-value; F: F-statistic; BMI: body mass index; ADCI: Alzheimer's disease-related cognitive impairment.
Associations between Alzheimer's disease–related cognitive impairment and extrinsic epigenetic age acceleration
Table 4 presents the results of logistic regression analysis, which evaluates the factors associated with ADCI, including EEAA. Among the various EAAs, EEAA was selected because it consistently differentiates between CU and ADCI groups in both men and women. In the overall participant model, EEAA is significantly associated with ADCI (OR = 1.18, 95% CI: 1.09, 1.28), along with APOE ɛ4 carrier status (OR = 7.30, 95% CI: 3.44, 15.49) and nutritional status (OR = 2.62, 95% CI: 1.17, 5.87). To evaluate the potential association of EEAA independent of age and APOE ɛ4, we constructed receiver operating characteristic (ROC) curves for two logistic regression models (Model 1: age + APOE ɛ4; Model 2: age + APOE ɛ4 + EEAA). Model estimates were used to compute the area under the curve (AUC). The addition of EEAA was associated with a higher AUC from 0.69 in Model 1 to 0.76 in Model 2 (Supplemental Figure 3).
Logistic regression models for predicting Alzheimer's disease-related cognitive impairment.
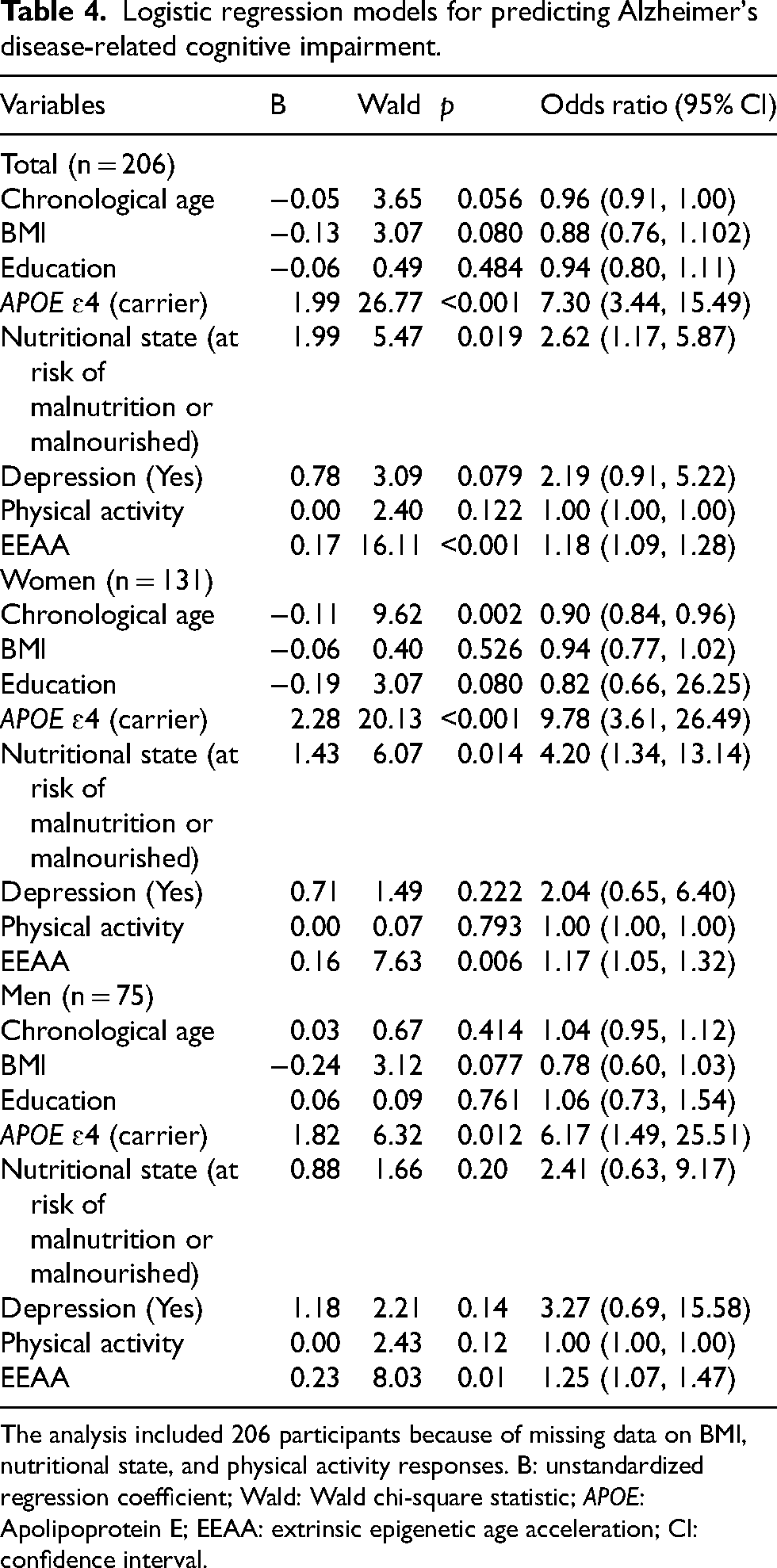
The analysis included 206 participants because of missing data on BMI, nutritional state, and physical activity responses. B: unstandardized regression coefficient; Wald: Wald chi-square statistic; APOE: Apolipoprotein E; EEAA: extrinsic epigenetic age acceleration; CI: confidence interval.
Among women, each unit increase in EEAA is associated with a 17.4% increase in the odds of developing ADCI (OR = 1.17, 95% CI: 1.05, 1.32). Among men, EEAA significantly increases the risk by 25.4% (OR = 1.25, 95% CI: 1.07, 1.47). Additionally, chronological age has a protective effect in women (OR = 0.90, 95% CI: 0.84, 0.96). APOE ɛ4 carrier status markedly increases the risk in both sexes, with women carriers exhibiting nearly tenfold higher odds (OR = 9.78, 95% CI: 3.61, 26.49) and men carriers showing over a sixfold increase (OR = 6.17, 95% CI: 1.49, 25.51).
Discussion
In this study, we demonstrated that EAA is associated with amyloid-positive ADCI and identified ADCI diagnosis and sex as factors influencing EAA. Additionally, we found that EAA, along with the APOE ɛ4 genotype and nutritional status, is significantly associated with ADCI. EAA was more pronounced in men than in women, both in healthy individuals and in those with AD.
Our primary finding was that EAA was correlated with AD. EAA measures, including EEAA, AgeAccelResidualHannum, and AgeAccelPheno, were significantly higher in the ADCI group than in the CU group, despite no differences in drinking, smoking, physical activity, or CRP levels between the groups. Even when ADCI was subdivided into MCI and AD dementia, the difference between CU and AD dementia in EEAA remained significant among the total participants and in men. Additionally, ADCI was associated with EEAA in both linear regression and logistic analyses. These results indicate that the ADCI group exhibits greater biological aging than the CU group. However, previous studies on EAA and its relationship with AD and neurodegenerative diseases have yielded inconsistent results. The epigenetic clock measured in brain tissues has been associated with the pathological diagnosis of AD and amyloid-β load. 6 In a longitudinal study spanning 15–20 years, both baseline blood and follow-up epigenetic age were associated with cognitive decline. 41 In a study of patients with frontotemporal dementia (FTD) and amyotrophic lateral sclerosis (ALS), higher blood EAA was linked to shorter disease duration and earlier onset. 42 Conversely, some findings suggest that blood EAA may not predict dementia risk. 10 Certain studies have failed to show a significant relationship between blood EAA and cognitive status or amyloid positivity.9,11 In one study, EAA was not associated with cognition or amyloid burden but was correlated with hippocampal volume, another biomarker of AD. 9
These inconsistencies may be attributable to a range of contributing factors. First, racial or ethnic variation may contribute to these inconsistencies. To date, most studies examining EAA in the context of AD have been conducted in North America and Australia.9–11 A prior study has demonstrated extensive DNA methylation differences between Europeans and South Asians, which are thought to be largely driven by differences in blood cell composition. 43 Second, inconsistency in results may be attributable to the age at blood sample collection. 44 Several studies have observed that in individuals aged 70 and above, epigenetic age appears younger, or exhibits reversed acceleration relative to chronological age.10,41 The average participant age was in the mid-60s, which is relatively young compared with previous cohorts. Finally, another factor may be the measurement methods employed. Previous research utilized different EAA measures, leading to variability in findings. In some cases, EAA was calculated simply as the difference between DNA methylation age and chronological age (AgeAccelerationDiff). In contrast, other investigations used regression models to adjust for potential confounding variables and estimated EAA as residuals from the regression of DNA methylation age on chronological age (AgeAccelerationResidual), a more recently developed approach. 44 In addition, EEAA, which yielded significant results in this analysis, has been measured in only a limited number of studies. EEAA is closely associated with the composition of immune cells in blood, particularly cytotoxic T cells (CD8 cells), helper T cells (CD4 cells), and B cells, all of which have been linked to systemic inflammatory responses and immune system aging. 2 Since neurodegenerative diseases such as AD are linked to chronic inflammatory states, EEAA may be especially sensitive to these pathological processes.
A second finding was that EAA was greater in men than in women, a pattern consistently observed in both the CU and ADCI groups (Supplemental Table 1). Furthermore, linear regression analysis controlling for factors such as APOE genotype and nutritional status identified sex as a significant factor influencing EEAA. These results align with previous research indicating that biological sex significantly influences the aging process and the development of age-related diseases.19,20 One investigation using HorvathAge and HannumAge demonstrated that men experience higher EAA than women across all age clocks. 45 Additionally, a twin study reported that women exhibited significantly reduced EAA compared with men. 21 Similar to the general population, men exhibited greater biological aging than women in the AD group in this study. However, further research is required to elucidate the implications of these differences in the mechanisms underlying AD. Prevalence of AD is higher in women, likely because of a combination of biological factors, such as hormonal differences, and environmental factors, such as education and occupation, with recent research also suggesting that higher male mortality from cardiovascular disease before dementia onset may contribute to this disparity. 12 Although EAA is associated with AD, explaining the difference in AD prevalence between men and women solely through the epigenetic clock remains challenging. This difference in EAA may reflect inherent sex-specific characteristics. Therefore, in AD research using EAA, it is appropriate to conduct separate analyses for each sex.
A further explanation for the protective effect of chronological age in women may be related to survival bias (Table 4). Due to lower midlife mortality among women, a larger proportion of women, including those at elevated risk of dementia, survive into older age. In contrast, higher midlife mortality among men leads to a more selectively healthy male population in later life. 12 As a result, chronological age may appear protective in women not because aging itself reduces AD risk, but because a greater fraction of at-risk women survive to older adulthood. These differential mortality patterns highlight the importance of considering survival effects when interpreting sex-specific associations between chronological age, EAA, and AD risk.
Our study demonstrated that EEAA measured in peripheral blood is associated with AD, regardless of sex. Although this study could not prove the precise mechanisms underlying the relationship between AD, brain disease, and peripheral EEAA, a potential link can be inferred based on the principles of EEAA measurement. As previously mentioned, EEAA may serve as a marker of immune system aging.2,46 Previous studies have shown that with aging, the balance between naive and senescent CD8 cells is disrupted. 47 These aged CD8T cells release large amounts of inflammatory molecules which can induce senescence in surrounding cells, including those in the brain.48–50 While some intrinsic aging markers, such as IEAA, appear resilient to these immune changes, extrinsic aging markers, particularly EEAA, are sensitive to shifts in cell-type composition. 2 EEAA, which reflects external influences on aging, may provide deeper insights into the interplay between aging and AD.
Brain inflammation plays a crucial role in the pathology and progression of various neurodegenerative diseases. 51 Evidence suggests that inflammatory substances generated peripherally alter the properties of the blood-brain barrier and the meningeal lymphatic system, thereby influencing the central nervous system. 52 In a study of patients with ALS and FTD carrying C9orf72 mutations EAA was similar between blood and brain tissue, except in the cerebellum. 42 There is also evidence linking inflammatory proteins to adverse brain outcomes53,54; however, whether this relationship is causal or consequential remains unclear.55–57 Our findings suggest a connection between peripheral immune system aging and AD, but further investigation is needed to elucidate the underlying mechanisms.
In contrast to the findings of Sugden et al., 7 we observed no significant difference in DunedinPACE between CU and ADCI in our study. Several factors may account for this discrepancy. Differences in age (75 years in ADNI versus 66–67 years in our study) and ethnicity could contribute to the observed differences. In addition, there were methodological differences: the FHS cohort used for validation in the study was not clinic-based but a population-based cohort, with the study design focused on assessing the risk of incident dementia. 7 Furthermore, this discrepancy may be explained by the conceptual properties of DunedinPACE, which reflects the current rate of biological aging rather than cumulative biological age. 40 Because AD pathology develops over a decades-long preclinical period, it is still unknown at which stage of the disease the accelerated aging processes begin to change noticeably. Thus, once the clinical syndrome of AD is established, the pace aging may not differ markedly from that of CU individuals.
Our study has several limitations. First, as a cross-sectional study, causal inferences cannot be drawn, and the temporal relationship between EEAA and cognitive impairment remains unclear. Specifically, the observed acceleration in epigenetic aging might partly reflect downstream biological consequences of AD pathology. Second, caution is required when interpreting the generalizability of our findings across different ages and populations. Our cohort differs from those in prior studies, as it comprises Korean participants who are younger and tend to have relatively lower educational attainment.10,11 For our findings to be broadly accepted and to help reconcile previous inconsistencies in the literature, external validation in cohorts encompassing diverse age ranges, ethnicities, and countries is necessary. Third, smoking and drinking were measured only as ever/never variables without details on current use or exposure intensity. This limited measurement likely attenuated their associations with EEAA, so the nonsignificant results should be interpreted cautiously. Finally, although data on medication use and certain metabolic or inflammatory comorbidities were unavailable and therefore could not be incorporated into the models, we adjusted for all other key demographic and clinical covariates known to influence epigenetic aging.
Nonetheless, we leveraged a well-characterized dataset, carefully controlled for factors that may influence epigenetic age, and focused on participants confirmed to be amyloid-positive to closely approximate AD pathology. Through these efforts, our study provides important insights into the potential relationship between EAA and AD. We suggest that EEAA should not be viewed as a substitute for established diagnostic biomarkers like p-tau 217, but rather as a complementary indicator of systemic biological aging and modifiable risk factors. This underscores the potential utility of EEAA in monitoring aging-related processes within the AD spectrum.
Supplemental Material
sj-pdf-1-alz-10.1177_13872877261431795 - Supplemental material for Sex-specific accelerated epigenetic aging in the Alzheimer's disease spectrum
Supplemental material, sj-pdf-1-alz-10.1177_13872877261431795 for Sex-specific accelerated epigenetic aging in the Alzheimer's disease spectrum by Jongmin Park, Hyuk-Jung Kwon, Joungsu Joo, Hyemin Jang, Sang Won Seo, Duk L. Na, Na-Yeon Jung and in Journal of Alzheimer's Disease
Supplemental Material
sj-docx-2-alz-10.1177_13872877261431795 - Supplemental material for Sex-specific accelerated epigenetic aging in the Alzheimer's disease spectrum
Supplemental material, sj-docx-2-alz-10.1177_13872877261431795 for Sex-specific accelerated epigenetic aging in the Alzheimer's disease spectrum by Jongmin Park, Hyuk-Jung Kwon, Joungsu Joo, Hyemin Jang, Sang Won Seo, Duk L. Na, Na-Yeon Jung and in Journal of Alzheimer's Disease
Footnotes
Acknowledgements
This publication is part of the PREMIER consortium. The authors would like to express their sincere gratitude to all the participants of the PREMIER project.
Ethical considerations
The original study (PREMIER consortium) was approved by the Institutional Review Board (IRB) of Samsung Medical Center, Seoul, Republic of Korea (IRB No. 2020-01-024). This study was exempted from review by the IRB of Pusan National University (PNU IRB/2022_93_HR).
Consent to participate
All participants provided written informed consent prior to their participation.
Consent for publication
Not applicable.
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by a grant from the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (HI19C1132); the Basic Science Research Program through the National Research Foundation of Korea (NRF), funded by the Ministry of Education (2017R1D1A1B04028328); the Bio & Medical Technology Development Program of the National Research Foundation (NRF), funded by the Korean government (MSIT) (No. 2022M3A9H1015723); the National Research Foundation of Korea (NRF) grant funded by the Korea Government (MEST) (NRF-2021R1A4A1021950); the National Research Foundation of Korea (NRF) grant funded by the Korean government (Ministry of Science and ICT) (No. 2022R1C1C1004826).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
The data supporting the findings of this study are available on request from the corresponding author. The data are not publicly available due to privacy or ethical restrictions.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.