
Abstract
Dementia with Lewy bodies (DLB), a synucleinopathy, is the second most common form of dementia after Alzheimer's disease (AD). Yet, DLB is often more aggressive and clinically challenging to diagnose due to overlapping symptoms with AD and Parkinson's disease dementia. Reliable biomarkers are necessary, since a definitive diagnosis of DLB currently requires postmortem tissue analysis. Blood-based biomarkers represent a minimally invasive and cost-effective avenue for earlier and more accurate diagnosis. This review integrates current evidence on blood biomarkers for DLB, at times extrapolated from other synucleinopathies, with a focus on α-synuclein and its derivatives, including extracellular vesicle-associated α-synuclein, seeding amplification assays, and autoantibodies. Additional biomarkers such as phosphorylated tau and amyloid-β highlight the frequent co-pathology with AD, while markers of neurodegeneration, neuroinflammation, and oxidative stress, heart-type fatty acid-binding protein, proteomic profiling, lipid, and other metabolites offer complementary diagnostic and prognostic potential. Although no single blood biomarker currently provides definitive diagnostic, differential, and prognostic accuracy for DLB, the studies reviewed here provide converging evidence that combined blood-based biomarker panels may offer meaningful clinical promise in facilitating earlier detection, guiding prognosis, and improving the design of targeted therapeutic trials for DLB.
Keywords
Introduction
Dementia with Lewy bodies (DLB) is the second most common form of dementia after Alzheimer's disease (AD), but it is often more aggressive and clinically disruptive. It typically presents with early neuropsychiatric and motor symptoms, including fluctuating cognition, visual hallucinations, parkinsonism, and sensitivity to antipsychotics.1,2 Age of onset is generally in the late 60 s to 70 s, and the clinical course is marked by rapid decline compared to AD, with greater risks of early institutionalization and mortality. 3 These factors contribute to a poor prognosis, particularly as neuropsychiatric symptoms often reduce quality of life and complicate patient care.
The burden of DLB is substantial, yet it remains under-recognized relative to its prevalence. Epidemiological estimates suggest that DLB accounts for 10–15% of dementia cases, though autopsy studies reveal that it may be underdiagnosed in life due to overlapping clinical features with AD and Parkinson's disease dementia (PDD). 4 The prevalence increases with age, but sex-based differences remain unclear, with some studies suggesting a slight male predominance. 5 Beyond prevalence, DLB is associated with higher healthcare utilization and costs compared to AD, largely due to the complexity of care and the neuropsychiatric symptoms that drive caregiver burden and hospitalizations. 1 Despite this impact, the disorder remains relatively underrepresented in clinical trials and research efforts compared to AD.
The defining pathological hallmark of DLB is the abnormal aggregation of misfolded alpha(α)-synuclein, which forms Lewy bodies and Lewy neurites within neurons.6,7 This pathology is not limited to the cortex but is distributed throughout multiple brain regions, disrupting synaptic transmission and neuronal survival. The aggregation process involves truncation and post-translational modification of α-synuclein, which accelerates fibrillization and oligomer formation, contributing to neuronal dysfunction and death.8–10 While α-synuclein pathology is the core feature, co-pathologies are common, with a high proportion of patients with DLB exhibiting tau and amyloid-beta (Aβ) pathology of AD, further complicating diagnosis and prognosis. 3
Currently, the diagnosis of DLB relies on consensus clinical criteria revised in 2017, including fluctuating cognition, REM sleep behavior disorder, visual hallucinations, and parkinsonism. 3 Non-invasive indicative biomarkers include reduced dopamine transporter (DAT) levels in the basal ganglia assessed by single-photon emission computed tomography (SPECT), reduced 123I-metaiodobenzylguanidine (123I-MIBG) uptake on myocardial scintigraphy, and polysomnography confirming rapid eye movement (REM) sleep behavior disorder. 3 However, definitive diagnosis requires postmortem neuropathological confirmation. Clinical diagnosis alone is often uncertain due to overlap with AD and PDD, and reliance on temporal rules such as the “1-year rule” has been shown to be flawed.4,5 Blood-based biomarkers represent a promising avenue to complement clinical assessment, as they are less invasive than cerebrospinal fluid (CSF) analysis and more cost-effective than imaging modalities.11,12 Blood biomarkers may allow for earlier detection, improved differentiation from other dementias, and better prognostic stratification of DLB by providing a biological pathway based framework for diagnosis (Figure 1).
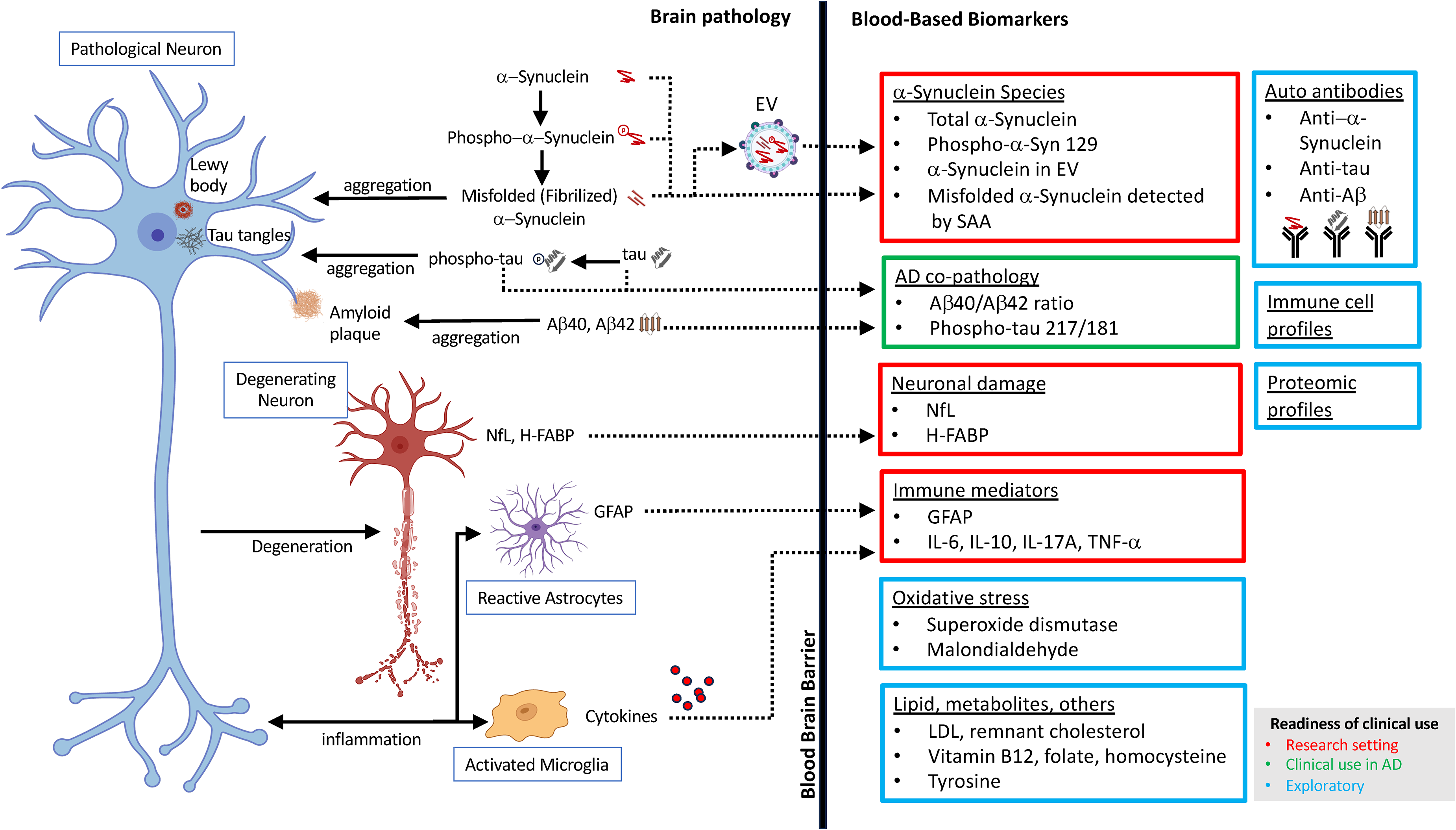
A schematic summarizing the blood-based biomarker pathways in DLB. Each class of biomarkers are color-coded for the level of readiness for clinical use estimated by the authors. Red: research setting. Green: clinical use in AD. Blue: exploratory. Created with BioRender.com.
α-Synuclein overview and diagnostic relation to DLB
The pathological aggregation of α-synuclein, manifested as intracellular spherical Lewy bodies and corkscrew-like Lewy neurites, is recognized as a hallmark of synucleinopathies.6,7 Although the precise mechanisms underlying the abnormal accumulation of α-synuclein in neurons and glial cells remain incompletely understood, it has been proposed that truncation of α-synuclein at its C-terminal acidic domain plays a role in this aggregation. 8 Recent breakthroughs in this area of research have identified the structure of α-synuclein fibrils in DLB. 10 Additionally, abnormalities in the non-amyloid domain of α-synuclein may also contribute to this process. 9
α-Synuclein is a protein found abundantly in the synapses and nuclei of neurons, from where its name was derived. 13 α-Synuclein is intrinsically disordered, with no permanent tertiary structure. Its properties make it amyloidogenic, partially explaining its specific aggregation in synucleinopathies. 14 α-Synuclein is also found as a monomer in the cytoplasm of neurons and glial cells; however, the abnormal aggregation potential of α-synuclein leads to its binding with lipid membranes and subsequent oligomerization.7,15–17 This binding and oligomerization can ultimately result in membrane collapse, disrupting cell communication and molecular transport. 18 Normal α-synuclein is believed to play several crucial roles within the central nervous system (CNS), including the regulation of synaptic vesicles and neurotransmitter release, and other synaptic processes. 19 Consequently, any disruption in its function can further contribute to cellular communication failures. 19
Regardless of the precise pathophysiological roles of α-synuclein, the presence of Lewy body pathology remains diagnostic for DLB and other synucleinopathies. Given the close association between DLB and α-synuclein, several diagnostic assays that utilize α-synuclein have been explored, including tests for this protein in CSF, skin, and blood with a goal to enhance diagnostic accuracy.
Extrapolation from other synucleinopathies
While DLB, PDD, Parkinson's disease (PD), and multiple system atrophy (MSA) share overlapping α-synuclein-driven pathology, they represent distinct clinical entities with established differences in disease onset, progression, and accompanying co-pathologies. As a result, biomarker findings derived from PD or PDD cohorts may not fully translate to DLB populations. Throughout this review, findings originating from studies of other synucleinopathies are explicitly identified as extrapolations where DLB-specific data are lacking. This distinction is crucial, as differences in these disorders may substantially influence the relevance of similar biomarker testing in DLB. However, these extrapolations are necessary to gain a comprehensive understanding of the current available blood biomarker resources and to guide future research endeavors, given the relative scarcity of large, well-characterized DLB-specific populations (Table 1).
List of potential blood-based DLB biomarkers.
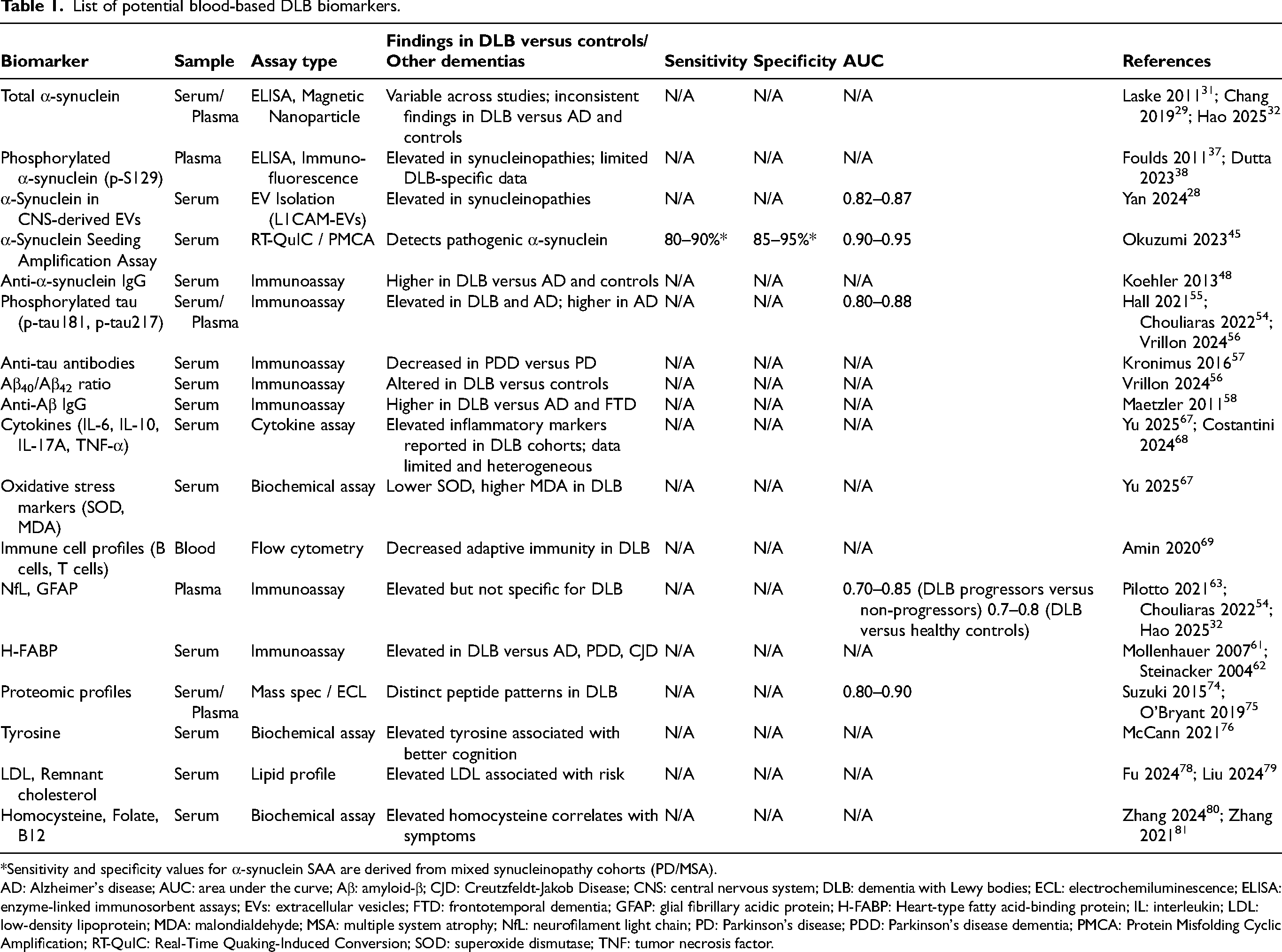
*Sensitivity and specificity values for α-synuclein SAA are derived from mixed synucleinopathy cohorts (PD/MSA).
AD: Alzheimer's disease; AUC: area under the curve; Aβ: amyloid-β; CJD: Creutzfeldt-Jakob Disease; CNS: central nervous system; DLB: dementia with Lewy bodies; ECL: electrochemiluminescence; ELISA: enzyme-linked immunosorbent assays; EVs: extracellular vesicles; FTD: frontotemporal dementia; GFAP: glial fibrillary acidic protein; H-FABP: Heart-type fatty acid-binding protein; IL: interleukin; LDL: low-density lipoprotein; MDA: malondialdehyde; MSA: multiple system atrophy; NfL: neurofilament light chain; PD: Parkinson's disease; PDD: Parkinson's disease dementia; PMCA: Protein Misfolding Cyclic Amplification; RT-QuIC: Real-Time Quaking-Induced Conversion; SOD: superoxide dismutase; TNF: tumor necrosis factor.
α-Synuclein in cerebrospinal fluid and skin
α-Synuclein levels in CSF have been studied extensively. However, due to conflicting results and significant overlap in values with healthy controls and other disease states, their utility as biomarkers for DLB remains limited until assays are further refined. 20 Several studies have found that total α-synuclein levels in CSF measured using enzyme-linked immunosorbent assays (ELISA) are lower in PD patients compared to healthy controls or patients with other non-degenerative neurological diseases. 21 In addition to total α-synuclein, other forms of α-synuclein in CSF have been investigated, including phosphorylated and oligomeric forms. 22
In 2019, skin biopsy kits were introduced to assess the presence of phosphorylated α-synuclein in cutaneous nerve fibers using immunofluorescence techniques. 23 Notably, the detection of phosphorylated α-synuclein in skin biopsies is nearly 100% specific for synucleinopathies, confirming the systemic deposition of these abnormal proteins in individuals with these conditions. 24 While no current studies exist, it is plausible to suggest a future in which a combination of multiple biomarker types, including skin samples, is utilized to aid in the diagnosis of DLB.
α-Synuclein in extracellular vesicles
The mechanisms by which α-synuclein migrates from the CNS to the blood are complex and not fully understood. One mechanism involves lipophilic transport proteins that act as “Trojan Horses,” facilitating α-synuclein to leave the CNS. Specifically, neuronal and astrocytic-originating extracellular vesicles (EVs) have been implicated in the transport of α-synuclein from the CNS to the systemic circulation. 25 EVs have multiple functions, including the transport of messenger RNA, non-coding RNAs, lipids, and proteins throughout the CNS, and have primary roles in cellular communication and maintenance of myelin/axons, antigen presentation, and synapse integrity.26,27
While playing a role in the expulsion of pathological α-synuclein from neurons in neurodegenerative conditions such as synucleinopathies, EVs also contribute to the transfer of α-synuclein in the CNS and outside of the CNS, as they are often absorbed by recipient cells in the transport pathway. 26 This unintentional mechanism likely allows α-synuclein to cross the blood-brain barrier and reach systemic circulation, where it can be quantified in the blood. The concentration of CNS-originating molecules in the systemic circulation is generally low; therefore, the direct capture of α-synuclein in the bloodstream is not always reliable. However, by analyzing CNS-originating EVs in the blood, the biochemical transport of α-synuclein may be better assessed. 25 The isolation of CNS-derived EVs has been promising in blood biomarker research for diagnosing synucleinopathies. In a cross-sectional analysis involving 576 serum samples, CNS-derived EV levels, specifically L1CAM-EVs, were used to differentiate patients with prodromal synucleinopathies from healthy controls with an area under the curve (AUC) of 0.82–0.87. 28 Importantly, this study also assessed other biomarker ratios, including Syntenin-1, to support these differentiations, highlighting a complex interplay between EV-α-synuclein and other CNS-derived products. 28
α-Synuclein in blood
α-synuclein in the blood can be measured in healthy controls, but typically at a much lower concentration than in individuals with a synucleinopathy. For example, one study using monoclonal antibodies with magnetic nanoparticles found that plasma α-synuclein levels in 40 healthy controls were averaged at 0.157 ± 0.285 pg/mL, while levels measured in 48 PD patients were averaged at 3.598 ± 2.531 pg/mL. 29 In the same study, the serum level of α-synuclein in healthy controls was averaged at 0.0038 ± 0.0020 pg/mL and 0.031 ± 0.042 pg/mL in PD patients. 29 Interestingly, the blood levels of α-synuclein and the disease characteristics associated with α-synuclein also vary based on sex. One study that used ELISA for quantifying α-synuclein in plasma found that lower α-synuclein levels were isolated in men with more severe PD, but this connection was not found in women. 30
The literature on blood α-synuclein analysis in specific DLB populations is limited. A study using ELISA analysis found that 40 patients with DLB had significantly lower levels of α-synuclein compared to 40 controls and 80 patients with AD, contrary to expectation. 31 However, a more recent study, using an ultrasensitive single molecule array (SIMOA) immune assay method, found that plasma α-synuclein level was elevated in individuals with DLB compared to healthy controls. 32
Several methodological factors likely contribute to this variability in total α-synuclein levels in blood. A major challenge is the high concentration of α-synuclein present in red blood cells, which can lead to significant measurement error when even minimal hemolysis occurs during sample collection or processing. 33 As a result, small differences in pre-analytical handling may substantially alter measured plasma or serum α-synuclein concentrations. In addition to pre-analytical variability, differences in assay design may influence reported values. Studies have used a variety of antibody epitopes and detection platforms, including conventional ELISA and more recently developed ultrasensitive detection systems such as SIMOA assay. 34 These technologies differ in their ability to detect low-abundance proteins and may target distinct forms of α-synuclein, including monomeric, oligomeric, or phosphorylated species. 35 Consequently, results across studies may not be directly comparable.
Given these limitations, total α-synuclein alone is unlikely to function as a reliable stand-alone biomarker for DLB. But combination analysis with other possible biomarkers may render these studies more valuable. For example, in a study enrolling 102 patients with PD, serum levels of α-synuclein, homocysteine, and leucine-rich α2-glycoprotein were measured, and elevations in all three of these molecules predicted cognitive impairment using the Montreal Cognitive Assessment (MoCA). 36
Besides total α-synuclein, phosphorylated α-synuclein at serine-129 (p-S129) has been assessed as a blood biomarker.37,38 Phosphorylated α-synuclein is the most common form of post-translational modification isolated from Lewy bodies and glial cytoplasmic inclusions. 38 Therefore, p-S129 may be a more promising marker than non-phosphorylated α-synuclein requiring further research.
α-Synuclein seeding amplification assay
α-Synuclein seeding amplification assays (SAA) originated as adaptations of prion disease assays, exploiting the ability of α-synuclein seeds to convert normal monomeric α-synuclein into oligomeric and fibrillar forms in a prion-like manner.39,40 Numerous studies have confirmed that various SAAs, including Protein Misfolding Cyclic Amplification (PMCA) and Real-Time Quaking-Induced Conversion (RT-QuIC), can effectively diagnose synucleinopathies using CSF with specificities as high as 95–100% versus non-synucleinopathies.41,42 Furthermore, SAA in different genetic populations of synucleinopathies yielded varied results. For example, in PD individuals with LRRK2 mutations, SAA found less α-synuclein relative to idiopathic PD and GBA-PD.43,44 While most SAA studies have been completed with brain or CSF samples, SAA has also been applied to analyze blood samples in individuals with synucleinopathies. A combination of the RT-QuIC assay and immunoprecipitation has proven effective in detecting pathogenic α-synuclein in these patients. Notably, the amplified seeds retain disease-specific properties, enabling the differentiation of samples from individuals with PD versus those with MSA based on α-synuclein levels and structure with a specificity of 85–95% and sensitivity of 80–90%. 45
Despite their strong diagnostic promise, α-synuclein SAAs, even with CSF samples, currently face several barriers to widespread clinical use. These include high operational costs, limited availability of specialized laboratory infrastructure, and the need for highly trained personnel. In addition, inter-laboratory variability and the absence of universally accepted standard operating procedures limit reproducibility across centers. 46 Given these limitations, at present, blood-based α-synuclein SAAs remain primarily research tools rather than clinical-grade diagnostics. Further validation of blood-based SAAs is required to establish robustness, standardized cut-off values, and real-world diagnostic performance in DLB populations. 47
Furthermore, many currently reported diagnostic performance metrics are derived from mixed synucleinopathy populations, including PD and MSA, rather than from studies focused exclusively on DLB. While these findings support the underlying biological relevance of α-synuclein seeding activity, the degree to which these results generalize to isolated DLB populations remains uncertain. Future studies specifically enrolling individuals with clinically probable or pathologically confirmed DLB will be essential to determine the true diagnostic performance of blood-based SAAs in this disorder.
α-Synuclein autoantibodies in the blood
Autoantibodies against α-synuclein have been assessed in multiple studies related to DLB, PD, and MSA. In one study, patients with DLB and AD were assessed for IgG to α-synuclein, revealing that individuals with DLB had significantly higher IgG production than AD and healthy controls. 48 Antibody studies completed for other synucleinopathies have confirmed the significance of anti-α-synuclein antibodies as biomarkers for these conditions, as these studies also found elevated antibodies to α-synuclein in patients with PD and MSA.49–51 Contrary to these studies, another study only reported significantly elevated anti-α-synuclein in patients with the genetic form of PD, suggesting a possible inherited role of these antibodies. 52
Tau biomarkers in DLB and AD co-pathology
While tau and Aβ are well-established pathological hallmarks for AD, a significant number of patients with DLB also exhibit AD pathology. Research indicates that in the brains of individuals diagnosed with DLB, the prevalence of moderate to severe AD co-pathology can exceed 70%.53,54 The presence of this AD co-pathology is well documented to be associated with an older age of onset, a quicker progression to dementia, and reduced overall survival rates.53,54
A study evaluating the serum sample from 300 participants, including 173 patients with DLB, for phosphorylated tau at threonine-181 (p-tau181) found that this protein was elevated in DLB and AD but was significantly more elevated in AD. 54 While not aiding in specific DLB diagnosis, it demonstrates potential for using blood tau biomarkers to differentiate between various dementias. Additionally, phosphorylated tau at threonine 217 (p-tau217) has been evaluated in the blood of individuals with dementia, including DLB and AD. Results of the analysis of blood p-tau217 in combination with tau-PET findings in individuals with DLB suggest that blood tau levels could help assess for DLB-AD co-pathology with an AUC of about 0.84–0.88. 55 Moreover, p-tau181 level was higher in individuals diagnosed with DLB than in healthy controls, suggesting a potential role of p-tau181 in DLB isolated from AD. 56 p-tau181 and p-tau217 have emerged as the most effective blood markers for differentiating amyloid-positive DLB from amyloid-negative DLB, consistent with previous reports of co-pathology with an AUC of about 0.8–0.85 with p-tau217 being slightly more reliable. 56
Tau has also been evaluated in blood through antibody analysis. One study that assessed the presence of antibodies against tau in serum in individuals with PDD and non-demented PD found that there was significantly decreased antibody production against tau in PDD, further identifying a role for tau in cognitive impairment and synucleinopathies. 57
Amyloid biomarkers in DLB and AD co-pathology
Blood-based Aβ biomarkers, specifically Aβ42/40 ratio, have been investigated in patients with DLB; however, current evidence suggests that changes in these measures primarily assess co-existing AD pathology rather than DLB-specific neurodegeneration. 56 Interestingly, when assessing IgG to Aβ in combination with IgG to α-synuclein was significantly higher in patients with DLB than in patients with AD and frontotemporal dementia. 58 While blood AD markers, such as p-tau 217/181 and Aβ40/42 ratio, do not diagnose DLB, they are increasingly recommended to be utilized in suspected DLB cases, since they provide important information identifying candidates for Aβ-directed therapies in AD co-pathology cases. 59
Heart-type fatty acid-binding protein in DLB
Besides the previously discussed measurements of α-synuclein, tau, and Aβ, researchers have explored alternative proteins and peptides in patients with DLB. One of the most promising proteins studied in patients with DLB is heart-type fatty acid-binding protein (H-FABP). H-FABP was first isolated in patients with myocardial infarction as a marker after myocyte damage but has since been studied in the context of nervous tissue injury. 60 One study that enrolled 33 individuals with a diagnosis of DLB found that they had significantly elevated H-FABP relative to healthy controls, individuals with AD, and individuals with PDD. 61 Similarly, another study revealed significantly elevated serum H-FABP in individuals with DLB compared to individuals with AD, Creutzfeldt-Jakob Disease, and healthy controls. 62
Neurofilament light chain and glial fibrillary acidic protein in DLB
Neurofilament light chain (NfL), a nonspecific indicator of neurodegeneration, and glial fibrillary acidic protein (GFAP), a nonspecific indicator of neuroinflammation, have also been evaluated in DLB. Several studies have demonstrated that circulating levels of these proteins are elevated in individuals with DLB relative to healthy controls, and are also higher in patients with DLB who exhibit cognitive progression compared to those who remain clinically stable.32,54,63 However, these increases are not unique to DLB and are observed across multiple neurodegenerative disorders. 64 Consequently, these biomarkers lack sufficient specificity to serve as standalone diagnostic markers for DLB, but may show promise in monitoring disease progression or severity.
Despite this limitation, NfL and GFAP may still provide clinically useful information. Because blood NfL levels correlate with axonal injury and overall neurodegenerative burden, these markers may be valuable for monitoring disease progression, estimating disease severity, or identifying individuals with rapidly progressive neurodegeneration.64,65 In this context, such biomarkers may function more effectively as indicators of disease activity or progression rather than as disease-specific diagnostic markers, particularly when interpreted alongside pathology-specific biomarkers such as α-synuclein or p-tau.64,65
Inflammatory biomarkers in DLB
Neuroinflammation in DLB is characterized by altered activity of microglia and astrocytes, particularly concerning α-synuclein clearance, as well as changes in immune cell populations. 66 In a study comparing patients with DLB, AD, and healthy controls, pro-inflammatory cytokines such as interleukin IL-6, IL-10, and IL-17A were found in the highest concentrations among those with DLB. 67 Additionally, oxidative stress was assessed in this study through measurements of superoxide dismutase and malondialdehyde levels, which were respectively lower and higher in DLB patients compared to controls. 67 Support for a peripheral inflammatory state in DLB was further corroborated by another study, which measured serum IL-6 and tumor necrosis factor-alpha (TNF-αmpa#alpha;) levels, revealing significantly elevated levels of these pro-inflammatory cytokines in patients with DLB compared to healthy controls. 68 Moreover, another study that examined a sample of 32 DLB patients found a reduction in the activity of the adaptive immune system, specifically a decrease in B cells and helper T cells relative to AD, alongside a hyperactive acute inflammatory response indicated by interleukin levels. 69 This observation suggests a potential pattern of low adaptive immune response coupled with heightened acute immune response in DLB patients, which, if reproducible, could serve as an indirect immunophenotype for DLB. 69
These immune alterations likely reflect α-synuclein–mediated activation of innate immune pathways, as misfolded α-synuclein promotes microglial activation and cytokine release while impairing protein clearance mechanisms. 70 However, these inflammatory biomarkers are not yet ready for clinical use because of variability of results, likely due to inconsistent methodologies, small sample sizes, and heterogeneities in patient age, disease progression, and disease severity. Additionally, the frequent presence of co-existing AD pathology in DLB may further contribute to variability in inflammatory and oxidative stress markers across studies. 71 Lastly, chronic neuroinflammatory signaling may lead to compensatory changes in adaptive immunity, including reduced circulating B and T cell populations, reflecting immune dysregulation at both central and peripheral levels. 72
Proteomic, lipid, and metabolites biomarkers in DLB
Recently, proteomic biomarker profiling has emerged as a promising avenue to discover new biomarkers for DLB. 73 One study used mass spectrometry (MS) to isolate peptide fragments including complement 4b, Wnt-2b, and lipopolysaccharide-binding protein. 74 This study found that different fragment profiles were distinguishable between DLB, AD, and controls. 74 Another study using electrochemiluminescence, panels of proteins were identified, including several cytokines, that could differentiate DLB and PD from controls and DLB from PD. 75 Although employing MS to analyze protein profiles in DLB patients may be too complex for routine clinical use, the identification of specific proteins highlighted in these studies could pave the way for future research utilizing more practical assays.
Research on amino acids in relation to DLB is limited; however, one study found a positive correlation between tyrosine levels and improved scores on the Mini-Mental State Examination for individuals with DLB. 76 This finding suggests that tyrosine may serve as a potential cognitive marker in this patient population.
Although apolipoprotein E4 (APOE4) is a strong risk factor for AD, APOE4 has also been increased in DLB cases independent of amyloid. 77 This potential connection between DLB and APOE4 has prompted research into lipid profiles within this patient population. Investigations utilizing genome-wide association studies and two-sample Mendelian randomization have demonstrated that elevated levels of low-density lipoprotein (LDL) are linked to an increased risk of DLB and may even exert a causal influence on the condition. 78 Additionally, a genome-wide association study assessed lipid profiles in individuals with APOE4-positive DLB and found a causal relationship (OR = 1.343; 95% CI, 1.094–1.649) between LDL/remnant cholesterol and risk of DLB. 79
Cyanocobalamin (B12) metabolism has also been explored as a blood biomarker in DLB. A study involving 82 patients with DLB found that elevated homocysteine levels correlated with higher scores on the Neuropsychiatric Inventory compared to healthy controls. 80 The study also demonstrated that homocysteine levels were higher, while folate and B12 levels were lower in individuals with DLB than in the control group. 80 A previous case-control analysis revealed similar findings, indicating that elevated total homocysteine levels are associated with DLB, and this elevation has a stronger correlation with DLB than with AD. 81 These studies suggest the potential for using serum homocysteine as a marker of neuropsychiatric severity in DLB, as well as to help differentiate DLB from AD.
Clinical and translational implications
Blood-based biomarkers hold significant potential to complement existing diagnostic tools for DLB. In current clinical workflows, the diagnosis of DLB relies heavily on clinical features supported by indicative biomarkers such as DAT imaging, cardiac MIBG scintigraphy, and polysomnography-confirmed REM sleep behavior disorder. 82 Blood biomarkers could serve as an accessible first-line biological screening method, particularly in settings where advanced technologies are unavailable or contraindicated, but it is unlikely to replace them in the near future.
An additional consideration in implementing blood-based biomarkers for DLB is the influence of systemic comorbidities common in older adult populations. Conditions such as chronic kidney disease, cardiovascular disease, and systemic inflammatory states may alter the clearance, metabolism, or peripheral concentrations of circulating proteins.83–85 These factors may introduce variability in biomarker measurements independent of neurodegenerative pathology. As blood-based diagnostic strategies are developed for DLB, future studies will need to account for these potential confounders to establish reliable reference ranges and clinically meaningful diagnostic thresholds.
In a realistic diagnostic framework, blood biomarker panels incorporating, for example, α-synuclein-related measures, p-tau181/217, and Aβ ratios, could be used to stratify patients by underlying pathology prior to confirmatory tests or CSF analysis. Such an approach may improve diagnostic confidence, guide the selection of additional diagnostic tools, and aid in prognostic counseling by identifying individuals with mixed DLB-AD pathology.
Limitations of current studies on blood-based biomarkers for DLB
A key limitation in the current literature on blood-based biomarkers for DLB is the frequent reliance on cohorts of patients with other synucleinopathy diagnoses, such as PD and PPD, reflecting the relative scarcity of large, well-characterized DLB-specific populations (Table 1). Although these extrapolations provide valuable mechanistic insight, they may overestimate diagnostic performance when applied to DLB, particularly in the presence of mixed pathology. Future studies that enroll individuals with clinically probable or pathologically confirmed DLB, independent of other synucleinopathy cohorts, will be critical to refining biomarker specificity and establishing clinically meaningful thresholds for this population.
Moreover, quantitative diagnostic performance metrics are available for only a subset of blood biomarkers evaluated in DLB (Table 1). Plasma phosphorylated tau biomarkers demonstrate moderate-to-high discriminative accuracy (AUC of approximately 0.80–0.88) for identifying AD co-pathology within Lewy body disease, but do not reliably distinguish DLB from controls. α-Synuclein SAAs exhibit high sensitivity and specificity for synucleinopathies in CSF; however, blood-based SAAs currently lack DLB-specific validation, with reported performance metrics largely derived from mixed synucleinopathy cohorts. In contrast, conventional plasma α-synuclein measures, EV–derived biomarkers, and peripheral inflammatory markers are primarily supported by descriptive or group-level analyses, with few studies reporting formal sensitivity, specificity, or AUC values.
Conclusions
Identifying reliable blood-based biomarkers for DLB should be a priority in DLB research to facilitate an earlier and accurate biological diagnosis of this disorder. Current studies suggest that DLB shares overlapping pathological features with other forms of dementia, notably AD. A confident diagnosis is needed for providing support for patients and their families in life planning, which is essential given the unique neuropsychiatric needs of this patient group.
Furthermore, research on blood-based biomarkers for DLB should prioritize large, longitudinal, and multi-center studies to validate the most promising candidates, particularly SAAs, EVs, and p-tau181/217, using more sensitive quantification technologies. These markers, when combined with Aβ ratios, may allow for the development of a reproducible biomarker profile that can differentiate DLB from AD and PDD while also stratifying for co-pathology. Additional work is needed to integrate supportive biomarkers, such as H-FABP, inflammatory markers, proteomic profiling, and metabolite markers into combined blood-based biomarker panels that capture both disease-specific and systemic aspects of pathology. Finally, future research should emphasize the translation of these biomarker profiles into clinical practice, where minimally invasive blood-based diagnostics could facilitate earlier detection, guide prognosis, and improve the design of targeted therapeutic trials for DLB.
Footnotes
Acknowledgements
Artificial intelligence–assisted tools (ChatGPT and OpenEvidence) were used for language editing and structural refinement of the manuscript. All scientific content was reviewed and verified by the authors to ensure accuracy, originality, and consistency with the intended scholarly interpretation.
Author contribution(s)
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This article publication was supported by the Office of Research and Development, Department of Veterans Affairs (grant number I01RX003347) to YBC.
Declaration of conflicting interests
The authors declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: JA declares no conflict of interest. NB declares no conflict of interest. VD declares no conflict of interest. YBC declares no conflict of interest. The contents do not represent the views of the U.S. Department of Veterans Affairs or the United States Government.