
Abstract
Introduction
The local hematopoietic bone marrow (BM) renin–angiotensin system (RAS) is a complex network of tissue-specific signaling pathways operating through autocrine, paracrine, and intracrine mechanisms that govern stem cell fate, lineage commitment, and marrow microenvironmental integrity. Neoplastic dysregulation of the local BM RAS drives leukemogenesis through a shift toward the proliferative angiotensin-converting enzyme (ACE)/angiotensin II (Ang II)/angiotensin type 1 receptor (AT1R) axis. The discovery of alamandine and its receptor, Mas-related G protein-coupled receptor D (MrgD), has expanded the non-classical RAS paradigm by introducing a distinct protective signaling branch.
Method
The alamandine–MrgD axis within the local BM RAS and its role in leukemic transformation was examined and synthesized by searching PubMed/MEDLINE using keywords including alamandine, MrgD receptor, bone marrow renin–angiotensin system, and leukemogenesis.
Conclusions
The alamandine–MrgD axis exerts anti-proliferative, anti-inflammatory, and antioxidant effects that counterbalance oncogenic ACE/Ang II/AT1R signaling in the hematopoietic marrow microenvironment. This axis functions as an endogenous counter-regulatory component of the BM RAS, modulating cellular proliferation, inflammatory signaling, oxidative stress, and microenvironmental homeostasis during leukemic transformation, positioning it as a critical regulator of physiological hematopoiesis. The biological expression and activity of the alamandine–MrgD axis may serve as both a prognostic biomarker and a therapeutic target in leukemia management.
Keywords
Introduction to the bone marrow renin-angiotensin system
We had first proposed that there is a local renin-angiotensin system (RAS) in the bone marrow (BM), three decades ago. 1 The hematopoietic BM RAS is an autocrine/paracrine network regulating physiological and neoplastic hematopoiesis (Figure 1). During the pathobiological complicated course of leukemogenesis, the local BM RAS network is dysregulated shifting from its biological role of hematopoiesis into promoting malignant leukemic progenitor cell survival and proliferation. 2
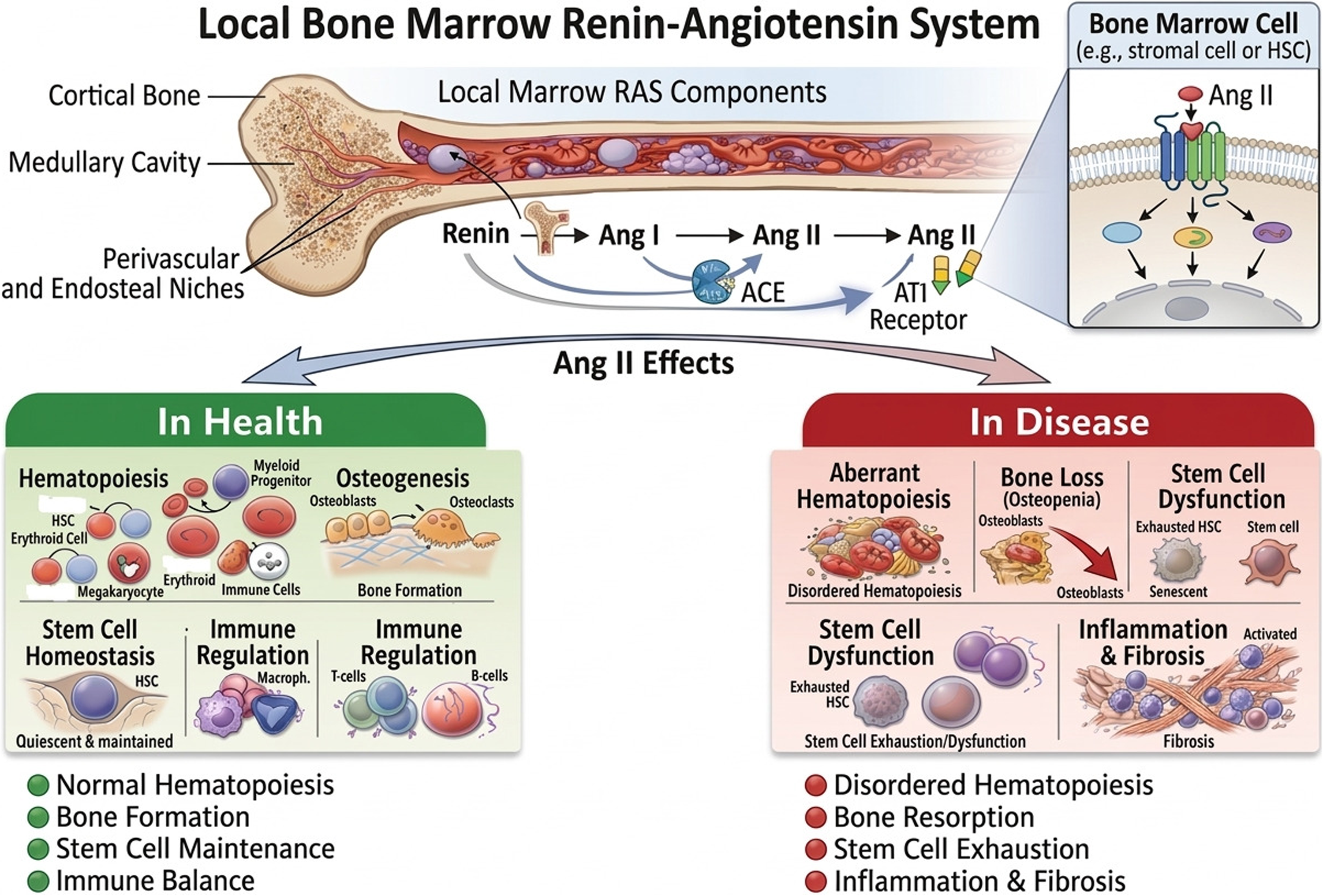
Local bone marrow renin-angiotensin system in health and in disease. The bone marrow has a locally active RAS. Under physiological conditions, local RAS signaling supports normal hematopoiesis, osteogenesis, stem cell homeostasis, and immune regulation. In abnormal conditions, Ang II activity that is not properly controlled can lead to abnormal hemostasis, bone loss (osteopenia), stem cell exhaustion and dysfunction, and inflammatory responses resulting in fibrosis.
Bone marrow renin-angiotensin system and leukemogenesis
The hematopoietic BM RAS functions as a dual counter-regulatory axis system during the complicated genesis of leukemia.3–5 Bone marrow is an active site of RAS peptide synthesis and receptor expression, which together form a local regulatory network that coordinates the life cycle of hematopoietic stem cells. Specifically, the angiotensin-converting enzyme (ACE)/ angiotensin II (Ang II)/ angiotensin type 1 receptor (AT1R) pro-neoplastic autocrine pathway promotes leukemogenesis while the ACE2/Ang1–7/MasR axis generally acts as a paracrine protective counterbalance.
6
The pathobiological interactions among local BM RAS and cellular neoplastic proliferation of leukemia manifest through several key mechanisms;
The alamandine/MrgD axis in bone marrow RAS and leukemogenesis: a novel paracrine hypothesis
The aim of this paper is to propose the hypothesis that the alamandine–MrgD axis11–13 within the context of local BM RAS, could be a critical regulator for maintaining physiological hematopoiesis in the hematopoietic marrow microenvironment. The hypothesis is that alamandine–MrgD axis represents a critical regulatory component of the bone marrow renin–angiotensin system with significant implications for leukemogenesis. By counterbalancing the pro-oncogenic effects of the classical ACE/Ang II/AT1R axis, this pathway modulates proliferation, inflammation, oxidative stress, and microenvironmental stability.14,15 The biological expressions of the alamandine–MrgD axis could be both a prognostic biomarker and a therapeutic target in leukemias.
Alamandine is a heptapeptide structurally related to Ang-(1–7), differing by substitution of alanine for aspartate at the N-terminus.16,17 This modification alters receptor specificity, shifting binding preference from Mas to MrgD. The discovery of alamandine, its Mas-related G protein-coupled receptor D (MrgD), and downstream metabolites such as alamandine-(1–5) has expanded the non-classical local RAS paradigm, introducing a distinct protective signaling branch for neoplastic disorders (Figure 2).
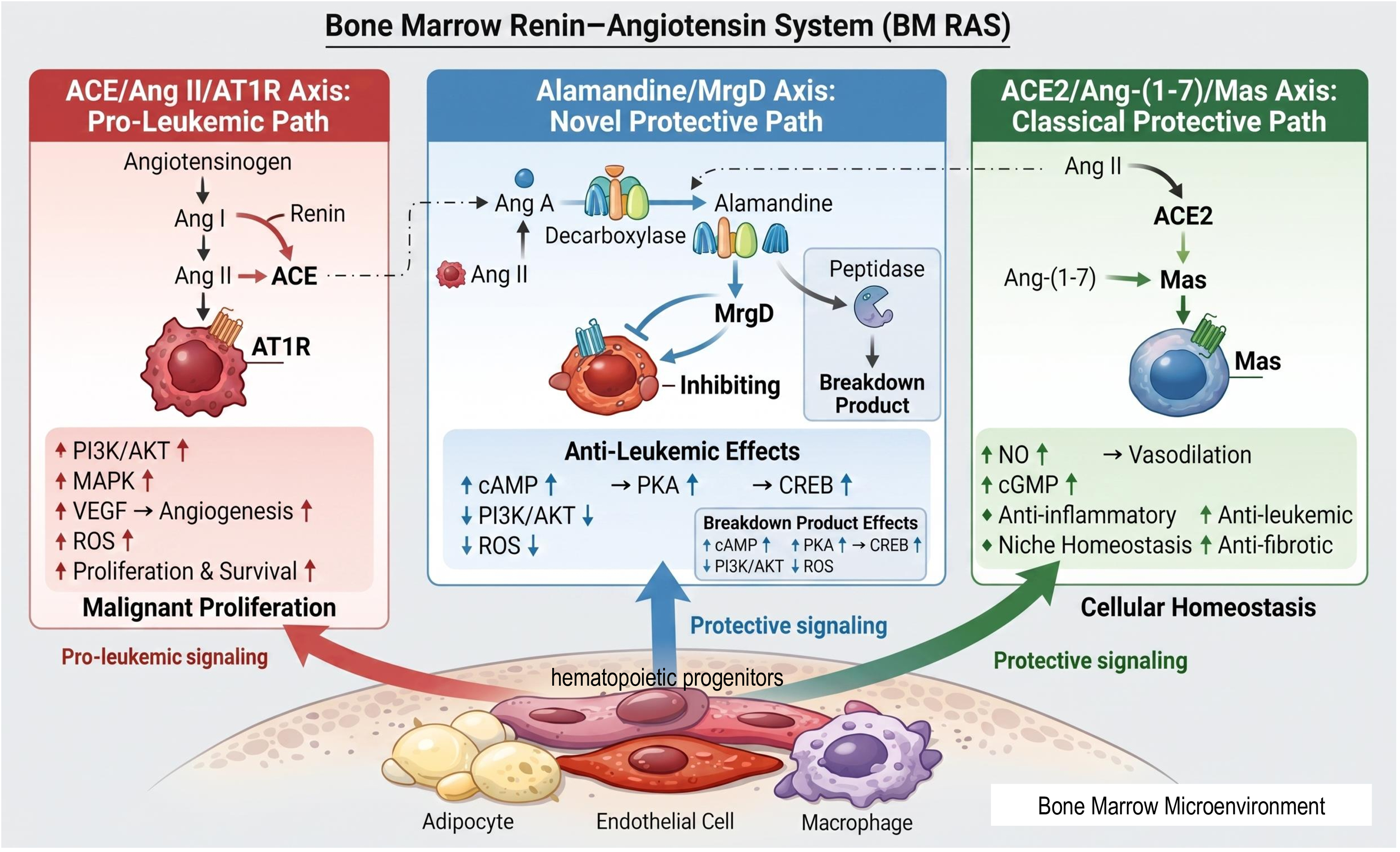
The Alamandine/MrgD axis as a novel paracrine pathway within the bone marrow hematopoietic RAS microenvironment. The schematic of the local bone marrow (BM) renin-angiotensin system (RAS) and its dual counter-regulatory axes. The diagram illustrates the integrated signaling pathways within the BM microenvironment, distinguishing between pro-neoplastic and protective branches. •Pro-Leukemic Path (Left Column): This section depicts the classical ACE/Ang II/AT1R signaling axis. Angiotensinogen is converted via Renin and ACE into Angiotensin II (Ang II), which binds to the AT1R receptor on morphologically irregular, spiked leukemic cells. Activation of this pathway leads to the upregulation (uparrow) of PI3K/AKT, MAPK, VEGF-mediated angiogenesis, and ROS, driving malignant proliferation and survival. •Novel Protective Path (Middle Column): This central section highlights the Alamandine/MrgD axis. Precursors such as Angiotensin A or Ang-(1–7) are converted into Alamandine by the Decarboxylase enzyme complex. Alamandine acts through the MrgD receptor to exert anti-leukemic effects, characterized by the stimulation (uparrow) of the cAMP/PKA/CREB pathway and the concurrent inhibition or downregulation (downarrow) of PI3K/AKT and ROS. The metabolic breakdown of Alamandine into Alamandine-(1–5) by Peptidase is also indicated. •Classical Protective Path (Right Column): This section details the ACE2/Ang-(1–7)/Mas axis. ACE2 converts Ang II into Ang-(1–7), which binds to the Mas receptor on spherical, healthy hematopoietic cells. This axis supports cellular and niche homeostasis through the upregulation (uparrow) of NO and cGMP, facilitating anti-inflammatory and anti-fibrotic responses. Bone Marrow Microenvironment (Bottom): The base of the figure represents the stromal niche, featuring adipocytes, endothelial cells, and macrophages. These cells serve as the primary paracrine sources of local RAS components.
Overall, the alamandine–MrgD axis exhibits anti-proliferative, anti-inflammatory, and antioxidant properties.18–21 Although alamandine-induced MrgD receptor activation could induce the protective antiproliferative biological responses, the MrgD gene itself had been identified as a potential proto-oncogene as overexpressed in neoplastic cells. Research indicates that MrgD exhibits intrinsic or constitutive activity. When overexpressed, which frequently occurs in various cancerous tissues, the receptor can assume an active form even without a ligand.22–24 This activation, despite the absence of a ligand, initiates downstream oncogenic pathways such as the MAPK/ERK and PI3 K/Akt cascades, leading to anchorage-independent growth and a loss of contact inhibition.22,25 In this scenario, merely having a high receptor density might be enough to induce malignant transformation, irrespective of the presence of its protective peptide partner, alamandine.
Besides alamandine, β-alanine, which was the first ligand identified for MrgD, triggers MrgD through Gq/Gi-coupled NF-κB signaling and IL-6 release—responses that do not occur with alamandine. It has been demonstrated to promote spheroid formation and cell proliferation in cancer cells expressing MrgD.23,24,33 Angiotensin A can attach to MrgD to produce vasoconstrictive and pro-proliferative effects before being metabolized into alamandine, indicating a shift towards the “classical” harmful RAS effects under certain physiological conditions. 23 Therefore, the clinical outcome seems to depend on the balance between the ligand and receptor. Alamandine likely functions as a biased agonist that stabilizes the receptor in a protective signaling state, effectively suppressing the receptor's intrinsic oncogenic drive. When alamandine's concentration is inadequate compared to the receptor density, the axis shifts towards the constitutive or alternative ligand-mediated pro-proliferative state. Thus the balance of the ligand alamandine relative to the receptor MrgD seems to be crucial since the absence of the proper protective peptide, an overabundance of receptors might be designated by the malignant cellular proliferation.22,34
Direct head-to-head comparisons of alamandine and angiotensin-(1–7) within the bone marrow (BM) niche are currently limited. 35 The two peptides signal through distinct receptors — MrgD versus MasR — and activate divergent downstream pathways, which have been demonstrated in cardiovascular and renal models. Whether these pathway differences translate into non-redundant functions specifically within the BM microenvironment has not been directly tested. The functional distinction between these two peptides in the BM is currently based on receptor-level divergence and tissue-extrapolated data, and requires experimental validation in the BM. The comparative biochemical and functional features of the classical and alternative RAS peptides are depicted in Table 1. The distinction drawn in Table 1 is based on receptor-level divergence from Ang-(1–7)/MasR signaling extrapolated from non-hematopoietic models.
Comparative biochemical and functional profiles of classical and counter-regulatory RAS peptides.
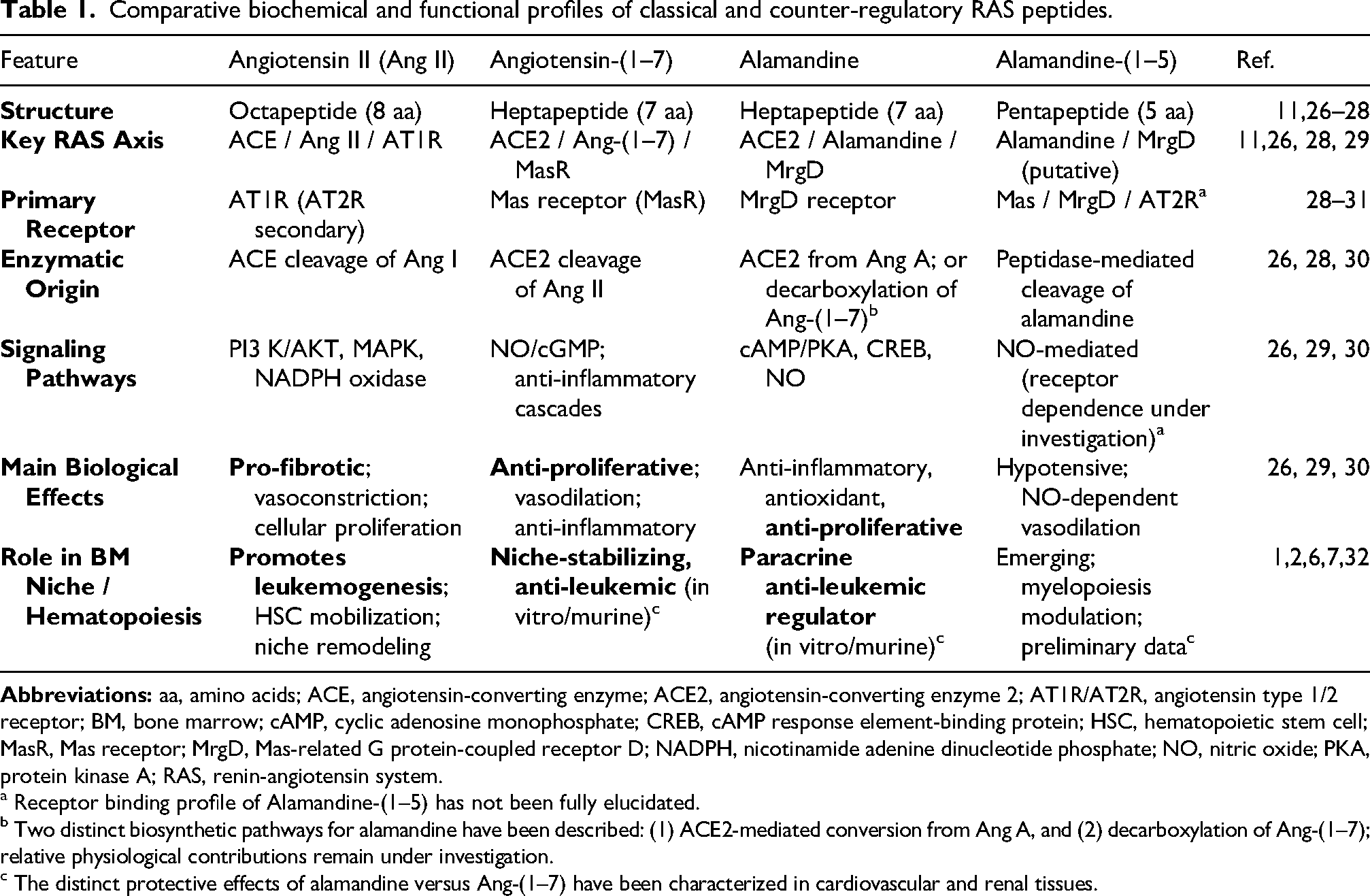
a Receptor binding profile of Alamandine-(1–5) has not been fully elucidated.
b Two distinct biosynthetic pathways for alamandine have been described: (1) ACE2-mediated conversion from Ang A, and (2) decarboxylation of Ang-(1–7); relative physiological contributions remain under investigation.
c The distinct protective effects of alamandine versus Ang-(1–7) have been characterized in cardiovascular and renal tissues.
Bone marrow RAS-associated leukemogenesis and the alamandine/MrgD paracrine axis
Molecular crosstalk between the classical and alamandine–MrgD RAS signaling in the bone marrow niche could affect the dual hematopoietic BM RAS functions counter-regulatory axis system during leukemogenesis (Figure 2). The BM ACE/Ang II/AT1R axis generate cellular proliferation where the paracrine ACE2/Ang1–7/MasR axis may act as an anti-proliferative manner. In the context of BM RAS-associated leukemogenesis, alamandine and its receptor MrgD represent a “second protective branch” that works alongside the ACE2/Ang1–7/MasR pathway to counteract the pro-leukemic effects of the classical BM RAS biopeptides.25,36,37 Alamandine supports Wnt signaling, maintaining stem cell quiescence and regenerative capacity. MrgD activation increases cAMP, leading to CREB phosphorylation and transcription of protective genes involved in stem cell maintenance.38–40
The interrelationships among the proliferative and anti-proliferative BM RAS pathways are defined based on the critical imbalances that favors leukemic progression.6,41 The classical pro-leukemogenic axis of ACE/Ang II/AT1R is typically overactivated in leukemogenesis. BM leukemic blasts significantly overexpress ACE (CD143) and renin leading to high local concentrations of Ang II.35,42–44 Ang II binds to AT1R on progenitor cells, activating signaling pathways like JAK-STAT and ERK that stimulate the proliferation of leukemic blasts via inhibiting apoptosis. ACE inhibitors and Ang II receptor blockers (ARBs) decreases the growth and colony-forming ability of leukemic blast cells.7,42,45 ACE/Ang II/AT1R axis promotes tumor angiogenesis via VEGF production and alters the BM microenvironment to support leukemic growth and metastasis. RAS genes can serve as biomarkers to predict sensitivity to anti-leukemic drugs such as Doxorubicin or Etoposide. 46 On the other hand, the anti-leukemogenic axis of ACE2/Ang1–7/MasR counteracts the proliferative effects of Ang II. ACE2 acts as a negative regulator of the classical axis by converting Ang II into Angiotensin-.1–7 Ang-1–7 binds to the Mas receptor (MasR), which exerts antiproliferative, anti-inflammatory, and anti-angiogenic effects, effectively braking the signals sent by the ACE/Ang II axis.2,6,7,47 Those RAS pathways supports normal hematopoietic recovery after injury but its activity is often suppressed or overwhelmed in the leukemic BM niche. The progression of leukemia is often driven by a shift in those RAS pathway balances.2,6,48,49 The proliferative axis of ACE/Ang II becomes dominant, removing the natural checks on cell growth provided by the anti-proliferative pathway of ACE2/Ang.1–7 High local ACE levels in leukemic microenvironment not only produce more Ang II but also accelerate the inactivation of Ang1–7 and other anti-proliferative inhibitory peptides like AcSDKP.6,8
Recently described Alamandine/MrgD paracrine axis represent a second protective branch in addition to the anti-proliferative ACE2/Ang1–7/MasR pathway in order to counteract the proliferative cellular effects of the classical ACE/Ang II/AT1R axis.50–53 Alamandine can be generated via the decarboxylation of Ang II to Angiotensin peptide followed by ACE2 processing. MrgD forms functional complexes with the Mas receptor, enabling coordinated signaling. This receptor crosstalk enhances anti-inflammatory and anti-proliferative effects, particularly in immune cells such as macrophages.26,30,54,55 Alamandine is chemically similar to Ang1–7 and is formed via the decarboxylation of Ang-1–7 or through the action of ACE2 on angiotensin peptides and acts as a functional antagonist to Ang II in BM microenvironment.27,56 The presence of in Alamandine/MrgD paracrine axis in the hematopoietic and stromal compartments suggests a role in the niche-level paracrine regulation within the BM. The co-expressions of the MrgD and MAS receptors allow for synergistic signaling, where the anti-proliferative and anti-inflammatory actions of alamandine in human macrophages and other hematopoietic progenitors.27,56 Alamandine reduces cellular proliferation and suppress the secretion of pro-inflammatory cytokines such as IL-6 and IL-1β in human macrophages. Since IL-6 and proliferative cytokines could drive leukemic blast growth, that anti-cytokine influence suggests a potential role for the Ala/MrgD axis in inhibiting leukemogenesis. Mas receptor and MrgD receptor can form heteromers (MasR–MrgDR complexes) on the cell surface. The antiproliferative effect of Alamandine actually requires the presence of both receptors. 57 The activation of the MrgD receptor can trigger signaling pathways such as AMPK/NO that directly oppose the hypertrophic and proliferative signals sent by the AT1 receptor.36,58–60 Ala/MrgD axis is novel paracrine pathway for maintaining homeostasis between the gut and BM within the context of aging and myelopoiesis.20–22 As the BM ages there is often a myelopoietic bias presenting as the increased production of myeloid progenitors, which can predispose individuals to myeloid clonal hematopoiesis. Alamandine treatment could help restore BM homeostasis and modulate this aging-related bias, potentially acting as a safeguard against the early stages of leukemogenesis.20–22
The gut-bone marrow axis as a distal regulator of leukemogenesis and the alamandine/MrgD paracrine axis
Alamandine can be generated via direct decarboxylation of Ang-(1–7) mediated by the gut microbiota.20–22 Beyond the gut, multiple organ systems have been identified as potential sources of systemic alamandine, though the cellular compartments responsible for its biosynthesis remain incompletely characterized.
The heart represents the most experimentally substantiated extra-intestinal source; isolated rat heart preparations have demonstrated local alamandine production following Ang-(1–7) perfusion, confirming the presence of the requisite enzymatic machinery within cardiac tissue. 28 Circulating alamandine has likewise been detected in human plasma, establishing its endocrine availability to distal tissues including the bone marrow.28,29
Renal dysfunction is associated with elevated alamandine levels, implicating the kidney in both its production and clearance.31,61 Adipose tissue constitutes an additional source and target. Alamandine suppresses leptin expression via MrgD in visceral adipocytes. High-glucose conditions downregulate both circulating alamandine and brown adipose tissue MrgD expression, suggesting a bidirectional metabolic–hematopoietic interface of potential relevance in obesity-associated hematopoietic dysregulation.28,32 More broadly, the tissue distribution of ACE2 encompasses the lungs, pancreas, liver, ileum, and bladder, all of which are likely biosynthetic sites, albeit without any quantitative confirmation to date.
The mechanistic pathway for the decarboxylation pathway has not been resolved yet, and there is no known human or mammalian enzyme with assigned aspartate decarboxylase activity. One hypothesis proposes that bacterial enzymes within the gut microbiome —and potentially within systemic circulation or other organ microenvironments— may catalyze this conversion, thereby linking the microbiome-RAS interface to alamandine availability across tissues.
Collectively, these findings position alamandine not merely as a locally acting paracrine mediator within the bone marrow niche, but as a peptide with multi-organ biosynthetic origins and systemic endocrine reach, whose hematopoietic implications may be shaped by the functional states of the cardiovascular, renal, metabolic, and intestinal systems.
The MrgD receptor expression is present in intestinal stem cells, vascular endothelial cells, and, most importantly, in the stem and progenitor cells of the bone marrow. There is a bidirectional gut-bone marrow axis of which intestinal microbial metabolites mediate the hematopoietic niche. Systemic inflammatory signals originated from the gut modulates bone marrow hematopoiesis. Aging is frequently associated with leaky gut characterized by increased intestinal permeability and the systemic leakage of bacterial products like lipopolysaccharides. Chronic exposure to those microbe-associated molecular patterns triggers a state of inflammaging in the bone marrow which induces a profound myeloid bias in the hematopoietic stem and progenitor cell developments. Myeloid bias results in the overproduction of pro-inflammatory monocytes and a decrease in lymphoid cells, creating a hematopoietic landscape that is highly susceptible to myeloid clonal neoplastic disorders. The alamandine/MrgD pathway plays a contributing component in maintaining the integrity of this gut-bone marrow connection.20–22 Alamandine treatment reverses intestinal permeability by regenerating the intestinal stem cell (ISC) layer through the activation of Wnt3α/β-catenin signaling in the colonic crypts. Moreover, alamandine restructures the aging gut microbiome, increasing the abundance of beneficial taxa like Bacteroides and Lactobacillus while decreasing pro-inflammatory genera such as Clostridium. The alamandine/MrgD pathway-induced shift in that microbial composition leads to a normalized metabolomic profile, with reduced levels of pro-myelopoietic metabolites in the cecum and circulation. By repairing the gut barrier and stabilizing the microbiome, alamandine prevents the distal inflammatory triggers that drive myeloid skewing and niche destabilization in the bone marrow. The modulation of the alamandine–MrgD axis 62 as a novel therapeutic target may affect leukemic cell metabolism and its integration into the gut–bone marrow axis.20–22 Whether pharmacological activation of the alamandine/MrgD axis may paradoxically upregulate classical RAS signaling through a compensatory feedback mechanism is both biologically valid and insufficiently addressed in the current literature. The RAS maintains a dynamic balance between its pro-inflammatory ACE/Ang II/AT1R and anti-inflammatory ACE2/Ang-(1–7)/Mas and alamandine/MrgD pathways.
Disruption of this equilibrium in either direction carries theoretical risks. Regarding the specific concern raised, two lines of evidence are relevant 63 : First, rather than triggering compensatory AT1R upregulation, MrgD activation by alamandine has been shown to directly antagonize AT1R activation by Ang II, suggesting that the two receptors operate in a competitive rather than a reciprocally amplifying fashion at the signaling level. This argues against a simple compensatory loop. 23 Second, however, MrgD expression has been shown to be upregulated under pathological conditions, including following Ang II treatment in cardiomyocytes, vascular smooth muscle cells, and aortic tissue, indicating that the axis is itself responsive to classical RAS activation. Whether the converse —sustained MrgD agonism leading to upregulation of AT1R-mediated signaling— occurs in a hematopoietic context remains unknown and constitutes a legitimate gap in the field. 53
Therapeutic modulation of the alamandine/MrgD axis must be considered within the broader context of RAS homeostasis, and that the net hematopoietic effect of such intervention will likely depend on the baseline activation state of the classical axis, particularly in leukemic microenvironments where Ang II/AT1R signaling is already dysregulated.
Molecular signaling perspectives for the alamandine/MrgD axis within the context of bm RAS
Molecular crosstalk between classical and alternative RAS signaling in the hematopoietic niche is essential for the homeostasis of the physiological hematopoiesis.6,46 Alamandine/MrgD axis exerts its marrow protective effects through a combination of transcriptional regulation and redox modulation (Table 2). The ability of alamandine to activate the Wnt3α/β-catenin pathway is a cornerstone of its niche-stabilizing function.21,59,64 Activation of the MrgD receptor stimulates adenylyl cyclase, leading to an increase in intracellular cAMP via the phosphorylation of the cAMP -response element-binding protein (CREB). Phospho-CREB then binds to the promoter regions of Wnt genes, inducing their expression.
Molecular crosstalks between classical pro-neoplastic RAS (ACE/ang ii/AT1R) and protective RAS (alamandine–MrgD) signaling in the bone marrow niche.

The relationship between phospho-CREB and Wnt gene induction has been established in several contexts relevant to intestinal stem cell biology and hematopoiesis. In intestinal stem cells, alamandine-mediated MrgD/cAMP/PKA signaling restores phospho-CREB and active β-catenin levels, with concomitant upregulation of Wnt3a in colon organoids — an effect blocked by adenylyl cyclase and CREB inhibitors, directly implicating the cAMP/PKA/CREB axis in Wnt3a transcriptional regulation. 66 More broadly, PKA signaling via CREB has been demonstrated to control the expression of canonical Wnt target genes including Wnt3a, Wnt1, and Wnt7a during developmental myogenesis. 67 Within the bone marrow niche specifically, multiple Wnt ligands including Wnt2b, Wnt3a, Wnt5a, and Wnt10b are expressed and regulate hematopoiesis through bone marrow stromal cells. Among these, Wnt3a promotes the expression of canonical Wnt target genes Axin2 and Ascl2 in intestinal stem cells, and its addition to aged organoid cultures restores a more youthful regenerative potential. 68
In the bone marrow microenvironment, that paracrine Wnt signal is crucial for maintaining HSC quiescence and self-renewal capacity, counteracting the inflammatory imprinting that would otherwise drive myeloid bias. Thus, Alamandine/MrgD pathway has the ability to bridge the gut-bone marrow axis further highlights its role as a key regulator of inflammaging and hematopoietic health.
The classical ACE/Ang II/AT1R axis is a major generator of reactive oxygen species (ROS) in the bone marrow, primarily through AT1R-mediated activation of the NADPH oxidases.18,52 Excessive ROS levels promote the oxidation of signaling proteins and induce DNA damage, which are hallmarks of leukemic progression and aging. Alamandine acts as a potent biological antioxidant, increasing the expression and activity of superoxide dismutase (SOD), catalase, and glutathione peroxidase. That antioxidant response reduces the ratio of BAX/BCL-2 and inhibits Caspase-3 activation, thereby protecting healthy hematopoietic progenitors from apoptosis while simultaneously disrupting the redox-sensitive growth signals in leukemic blast cells. In the context of BM RAS-driven leukemogenesis, where Ang II dominance often suppresses the production of protective heptapeptides, the resulting ligand-starved MrgD receptors may be co-opted by the malignant niche to promote survival. Consequently, therapeutic strategies aimed at restoring alamandine bioavailability or enhancing ACE2 activity may represent a rational approach to harness the potential anti-leukemic properties of this axis, warranting further experimental investigation.
Clinicopathological correlations synthesizing the autocrine/paracrine axis of the local bm RAS biopeptides
The alamandine/MrgD axis seems to be an important emerging multi-functional component of the expanded RAS that serves as a novel paracrine pathway within the bone marrow niche via providing a powerful counter-regulatory signal to the pro-leukemogenic ACE/Ang II/AT1R pathway. Beyond hematological malignancies, emerging evidence suggests that the alamandine/MrgD pathway could induce anti-neoplastic activities in other pancreatic, lung, and renal carcinomas. 65 In pancreatic and lung carcinoma cell lines, adamantine reduced tumor cell proliferation in an MgD-dependent manner. This axis caused a change in metabolism from anaerobic to aerobic glycolysis, supressed the PI3 K/AKT/mTOR pathway, and activated the pro-apoptotic transcription factor FoxO1, but did not have any anti-proliferative effect in non-tumoral cell lines. 69 In renal cell carcinoma, MrgD expression has been documented across multiple tumor subtypes. However, high MgD expression in metastatic clear cell renal cell carcinoma was paradoxically associated with worse prognosis and advanced disease stage. The oncological significance of this axis is dependent on context and may differ from its homeostatic counter-regulatory function in certain tumor microenvironmental conditions.22,70 Likewise, alamandine's inhibition of pathological retinal neovascularization via the HIF-1α/VEGF pathway hints at its potential role in managing the angiogenic complications of malignant transformations.71,72
Collectively, these findings position the alamandine/MrgD axis as a pleiotropic component of tumor biology whose anti-neoplastic potential is most clearly demonstrated in metabolically active epithelial malignancies. This axis may serve as a broader biomarker for carcinogenesis across multiple organ systems. Further mechanistic investigation is needed to determine its effect on renal and potentially other solid tumors.
The inclusion of RAS-based biomarkers for risk stratification 46 and the development of stable pharmacological agonists for the MrgD receptor 73 offer promising strategies for improving outcomes in the challenging clinicopathological management of leukemias. Future pharmacological research must target the exploration of stable alamandine analogs that can be effectively delivered to the bone marrow microenvironment to restore the niche's inherent biological defenses against leukemi development. In the era of targeted inhibitors and precision medicine, the alamandine/MrgD axis stands as a crucial, yet underutilized, resource in the fight against BM RAS-driven leukemogenesis. The key areas for future research include the development of stable alamandine analogs, clinical validation of the MrgD-targeted therapies, the integration into precision oncology frameworks and exploration in the genesis malignancies particularly leukemogenesis. Advancing this field will require translational studies focused on pharmacological activation of MrgD signaling and clinical validation in hematologic malignancies. The alamandine–MrgD axis thus represents a promising frontier in the development of microenvironment-targeted therapies in hematological neoplastic disorders.
Footnotes
Authors’ contributions
ICH conceived the idea. AY and ICH wrote the initial draft, prepared the figure and extensively edited the manuscript. All authors reviewed the final manuscript.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.