
Abstract
Evolutionary criminology is an approach to the understanding of crime and criminality that is based in part on key aspects of evolutionary psychology. The approach allows for a renewed examination of traditional criminological assumptions and can serve to further enhance theoretical viewpoints on antisocial behavior. The recently developed evolutionary taxonomy theory is an example of such an approach. Relying on the tenets of life history theory, the evolutionary taxonomy was proposed as a theoretical scaffolding for Moffitt's developmental taxonomy of offending. While recent tests of the evolutionary taxonomy have been informative, lacking from the existing literature is an assessment of the extent to which measures of life history theory can predict classification into offending groups based on Moffitt's developmental taxonomy. The current study provided a partial test of classification predictions using the National Longitudinal Study of Adolescence to Adult Health study (n = 12,012). Results of multivariable regression analyses indicated that measures associated with somatic effort and aspects of the developmental environment were predictive of group classification, but measures associated with reproductive effort were not. Implications for evolutionary criminology and traditional criminology are discussed.
Keywords
Introduction
Criminological examinations of the etiologies of antisocial behavior have tended to focus on sociological factors, including but not limited to parenting, cultural institutions, and social bonds (Gottfredson & Hirschi, 1990; Hirschi, 1969). While these factors and others derived from a traditional criminological approach have illuminated aspects of why people engage in criminal offending, they often leave a great deal of explained variance unaddressed (Weisburd & Piquero, 2008). Relatively recently, criminologists have incorporated approaches from disciplines outside of sociology to help bolster their understanding of criminal activity. Perhaps the most obvious area in criminology where this infusion of interdisciplinary approaches has taken place is in biosocial (aka, biopsychosocial) criminology.
Relying on a vast array of theoretical and methodological approaches from disciplines such as behavior genetics, neuroscience, psychology, and evolutionary biology, biosocial criminologists have produced a considerable amount of research that has both challenged and supported aspects of a traditional criminology. Nonetheless, even within biosocial criminology there are areas that are relatively underexplored. One such area is the topic of the current special issue: evolutionary criminology. While biosocial criminology has provided increases in our understanding of some proximal causes of individual differences in antisocial behaviors, there is a gap in terms of our understanding of ultimate explanations of such phenotypic variation (Mealy, 2000; Wiebe, 2012). A potential argument for why biosocial criminology has lacked in terms of ultimate causes is its perception at times as an atheoretical approach to studying crime and criminality (Walsh, 2011). Evolutionary criminology, however, provides a theoretical scaffolding on which much of biosocial criminology—and, indeed, traditional criminology—can unify and progress (Boutwell et al., 2015a).
Key to an evolutionary approach to criminal conduct is the conceptualization of antisocial behavior—not as a collection of actions relative to legal statutes, but rather as a social strategy employed to potentially obtain resources that help increase the likelihood of survival and reproduction. Wiebe (2012) put it succinctly: “Crime is a way to get what one wants . . . [but] Why do we want what we want?” (p.348; emphasis in original). As evolutionary biologists (and more recently evolutionary psychologists) have long pointed out, organisms are presented with various adaptive problems the solutions to which aid in survival and reproduction (Buss, 1991). Thus, the things we want, on average and over evolutionary time, help solve various adaptive problems. Seen from an evolutionary point of view then, crime and criminality are simply potential behavioral strategies that individuals can rely upon to obtain what they want. However, as control theorists in criminology (e.g., Hirschi, 1969) have pondered, why then doesn’t everyone engage in crime?
While historical criminological responses to the question of why everyone isn’t constantly engaged in crime have focused on variation in individual-level traits (e.g., self-control; Gottfredson & Hirschi, 1990) and/or (in)formal social institutions (e.g., parenting or involvement in after-school activities; Hirschi, 1969) an evolutionary criminology response focuses on behavior as an extension of adaptive strategies. More specifically, given that ours is a highly social species, survival and reproduction was, on average, heavily reliant on cooperation. Thus, most people avoid criminal involvement as a strategy of cooperation. Of course, within a social network where cooperation is the norm social cheaters can benefit by exploiting such cooperative tendencies (Dawkins, 2006; Mealy, 1995). However, as researchers have illustrated (e.g., Axelrod, 1984), such strategies tend to reach a maximum saturation point before they are no longer beneficial in the long term, yet such strategies will always be present in a social environment based on cooperation (Dawkins, 2006). Thus, the reason everyone doesn’t commit crime is because it wouldn’t benefit everyone: there are more optimal strategies that exist in given environments.
Who then benefits from engaging in cheating as manifested by criminal behavior? Daly and Wilson (1998, 2005) and later Daly (2016) argued that criminal conduct, especially serious violent conduct, arises in those situations where competition for scarce resources is most severe (see also Mishra & Lalumière, 2008). Arguing against the idea that violent conduct is a pathological response to horrible conditions, these authors contend that such conduct is a function of the intense cost of not competing intensely—indeed, they argue that violent conduct should instead be viewed as a rational response to pathological social conditions (Daly, 2016). Thus, strategies to approach adaptive problems are expected to manifest as antisocial primarily in social conditions that provide corresponding environmental cues. In essence, this process describes some of the main pillars of life history theory.
Briefly, in life history theory strategies for solving adaptive problems can be conceptualized as falling along a continuum whereby faster life history is exemplified by strategies emphasizing short-term gains and slower life history is exemplified by strategies that accentuate long-term gains (Brumbach et al., 2009; Kruger et al., 2015). The approach predicts that within-species variation in strategies related to maturation, overall health and well-being, sexual behaviors, risk-taking, and parental investment will manifest as a function the stability or predictability, among other cues, of the developmental environment. In general, based in part on environmental cues strategic tradeoffs can take place between somatic efforts (growth, health, and well-being) and reproductive efforts (acquiring mates and producing offspring; Roff, 2002). While environmental cues provide information to direct the allocation of strategies, researchers have also highlighted that individual differences beyond external cues can also influence behavioral repertoires (e.g., Figueredo et al., 2005). A key assumption in life history theory is that life history strategies are not consciously followed or resistant to alterations (i.e., phenotypic plasticity; Belsky et al., 1991). Furthermore, a key claim of the approach is that organisms have species-typical strategies. For example, most humans exhibit a slow life history strategy given their relatively long lifespan. However, variation can manifest as a function of environmental cues and individual differences which guide the biological and behavioral strategies individuals can manifest. Thus, life history theory can guide predictive assessments of an evolutionary approach to understanding crime and criminality.
In terms of how criminology can benefit from an infusion of life history theory, it is often most instructive to apply evolutionary ideas and principles to existing theoretical approaches on antisocial behavior. One of the most well-known and well-supported approaches to understanding antisocial behavior over the life course is Moffitt's developmental taxonomy (Moffitt, 1993, 1994). What follows is a review of Moffitt's key ideas and an example of an application of evolutionary principles to Moffitt's ideas with the recently developed evolutionary taxonomy.
Moffitt's Developmental Taxonomy
Readers of the current journal may not be as familiar with Moffitt's developmental taxonomy as those in criminology. Thus, a brief review is necessary. Moffitt's initial proposition (Moffitt, 1993) and subsequent work (Moffitt, 1994) outlined the main tenets of her theory. Her ideas were driven by a thorough review of the existing empirical literature on antisocial behavior over the life course and a recognition that theoretical siloes and the resulting blind spots had prevented the consilience necessary to provide a sufficient understanding of the observed patterns. Moffitt proposed a dual-taxonomy of antisocial behavior that accounted for two primary well-established empirical patterns: the age-crime curve and variability in relative stability in antisocial behavior over the life course. The age-crime curve refers to the considerable increase in offending that tends to accompany adolescence and is later followed by desistance with entry into young adulthood. Such a pattern of offending behaviors resulted in generations of criminologists focused solely on the period of adolescence in a search for etiological factors. Moffitt argued, however, that without a life course approach—where behavioral patterns are observed from childhood to adulthood—it was impossible to distinguish those whose antisocial behaviors were limited to adolescence from those who offended throughout all periods of their lives. Moffitt proposed two types of offending groups: adolescence-limited (AL) and life course persistent (LCP). She argued that AL offenders lacked consistent displays of antisocial behavior in childhood and pre-adolescence but upon entry into adolescence delinquency was quite common, though the offending tended to be relatively minor. LCP offenders, however, were characterized by patterns of antisocial behaviors early in life and that continued into adolescence and well into adulthood. Additionally, Moffitt argued that the offending behaviors exhibited by LCP offenders were much more serious than ALs, especially during the adolescent years.
In addition to proposing the dual-taxonomy, Moffitt also put forth etiological claims that differentiated LCP from AL offenders. For LCPs Moffitt's claim was a biosocial one: the interactive effect of neuropsychological deficiencies and adverse developmental environments led to persistent and serious antisocial behaviors throughout the life course. Given the relative rarity of the etiological factors, Moffitt argued that about 5–10% of most populations would exhibit LCP offending behavioral patterns (Moffitt, 1993). Alternatively, given that AL offending was the expected norm Moffitt's etiological claims centered not on factors that were atypical but rather those that were commonly experienced.
Moffitt's claims were no less biosocial, however, as they relied on recognition of evolutionary processes and socio-cultural factors. Moffitt argued that AL offending patterns were a result of a strain caused by entrance into a period of the life course that was historically, including in our evolutionary past, one wherein youth transitioned into adulthood and the social freedoms thereof. However, modern societies instead have imposed cultural expectations that youth remain in places of constraint: at home, living with parents or guardians who dictate several aspects of youths’ lives and at school, where rule following and time management are imposed on youth. Layered into this period of expected freedom is the fact that in adolescence youth reach puberty and are therefore biologically fully mature (i.e., they can reproduce). Thus, a conflict exists between biological maturity (along with the expected associated freedoms) and the social constraints placed upon youth. Moffitt termed this disjunction between biological and social maturity the maturity gap. She further argued that to address the psychosocial discomforts associated with the maturity gap, youth sought cues from their environments for how to gain more freedom. One essential cue was the observation of the apparent freedom of LCP offenders. These similar-aged peers ostensibly possessed the social freedoms sought by those experiencing a maturity gap and so were followed in a behavioral pattern that Moffitt called social mimicry. This explanation of AL offending also conformed to a well-established observation in the criminological literature of the consistent impact of delinquent peers on offending patterns. The social mimicry process, however, is not one that lasts beyond adolescence when youths’ social maturity eventually matches the biological maturity (i.e., the maturity gap is no longer present). Thus, with no source of psychosocial strain those in early adulthood are free to follow their desired acts of freedom and not engage in rebellious offending. The desistence process takes place and the drop in the aggregated age-crime curve manifests.
Moffit also drew on curious observations from the literature that pointed to a group of people who never engaged in offending behaviors: abstainers. This group, Moffitt argued, represented a small segment of the population (about 10% or so) and experienced a life trajectory that was divergent from the traits and experiences of either LCPs or ALs. Moffitt suggested that while abstainers lacked the neuropsychological deficits and adverse rearing environments, they may possess certain personality traits that prevent interactions with peer groups. Additionally, she suggested that abstainers avoided experiencing the maturity gap and didn’t exhibit AL offending patterns due to several potential reasons. For example, they may come of age in developmental environments that required early maturational activities (e.g., rural farms) and so there was no experience of a maturity gap. Alternatively, they may have delayed pubertal development such that once they are biologically mature they are also closer to social maturity than AL offenders.
While empirical evidence of Moffitt's theory hasn’t been unequivocal (Chen & Adams, 2010; Nedelec et al., 2016), several reviews have indicated that her claims—especially those related to LCP and AL offending—have received considerable support (see Moffitt, 2018). Some authors, however, have noted that while Moffitt's developmental taxonomy provides excellent proximal explanations for observed patterns in offending behaviors it lacks a framework which addresses the more distal, or evolutionary, underpinnings for her theoretical claims (Boutwell et al., 2013; Wiebe, 2012). Recently, Boutwell and colleagues (2013, 2015a, 2015b) have proposed that Moffitt's theory can be extended with the infusion of evolutionary theory, more specifically life history theory, to better contextualize the etiological claims. The following section briefly outlines these ideas as they relate to the current study.
The Evolutionary Taxonomy
In a series of papers, Boutwell and colleagues (Boutwell et al., 2013, 2015a, 2015b) advanced the evolutionary taxonomy as a unified crime theory. Extending the work of Rushton (1985) and Ellis (1988), the evolutionary taxonomy was proposed to provide an overarching theoretical scaffolding for Moffitt's developmental taxonomy. In applying an evolutionary framework, it was argued, Moffitt's etiological claims can be enhanced with ultimate explanations. For example, while LCP offending patterns can be proximally explained by neuropsychological deficiencies and adverse rearing environments it's not clear why such offending patterns should manifest from these causes. The evolutionary taxonomy, however, argues that the offending patterns could result due to the environmental cues (e.g., shortened time horizon) that initiate the LCP behavioral pattern. Additionally, psychopathologies observed in LCP populations could also serve to hinder their recognition of the benefits of cooperation and over-emphasize the potential benefits of strategies laden with antisocial conduct (i.e., fast life history).
Thus, far tests of the evolutionary taxonomy have primarily focused on illustrating the ways in which it expands understanding of LCP offending patterns (e.g., Boutwell et al., 2015a, 2015b). Absent in the literature are tests of how components of the evolutionary taxonomy are associated with classification into all the groups proposed in Moffitt's theory. The current study partially addressed this gap by testing how measures tapping life history strategies (a pillar of the evolutionary taxonomy) predicted offending classification groups using a longitudinal sample of American youth who were followed into mid-adulthood.
Hypotheses
Based on the predictions outlined in the evolutionary taxonomy, the following hypotheses were tested in the current study. First, it was expected that measures of somatic effort would be negatively associated with classification in the offending groups: poorer overall physical and mental well-being in adolescence was expected to predict more offending over the life course. Second, it was expected that measures of reproductive effort would be positively associated with classification in offending groups such that those who directed more effort to reproduction in adolescence would also report more offending over the life course. Third, it was hypothesized that measures of the developmental environment in adolescence would be predictive of classification in the offending groups. More specifically, cases indicating a riskier, more unpredictable, and/or anticipating a more negative time horizon (i.e., shorter anticipated lifespan) would also report more offending over the life course. To test these expectations, the current study employed data from the restricted version of the National Longitudinal Study of Adolescence to Adult Health (Add Health; Harris et al., 2009).
Methods
Sample
The Add Health began with two stages of data collection at Wave 1 in 1994 to 1995. First, an in-school questionnaire was administered when participants were in grades 7 to 12 (n = 90,118). Second, in-home data collection occurred for a subset of the overall school samples (n = 20,745). Participants from the in-home sample were re-interviewed in 1996 (Wave 2; n = 14,738), which was about a year later for most respondents. The third wave of data took place in 2001 to 2002 (n = 15,197) when most of the respondents were entering early adulthood. Wave 4 data collection occurred in 2007 to 2008 (n = 15,701) and a fifth wave of data was recently collected beginning in 2016 and ended in 2019 (n = 12,300). The current study uses data from the first four waves. For more information about the Add Health study, including sampling strategy see Harris et al. (2009).
The analytical sample for the current study was formed in a multi-step process. First, to ensure that all life history measures occurred in adolescence we reduced the sample to only those cases who were 18 or younger at Wave 1. Second, after creation of all study measures (see below) only those cases with valid scores on the antisocial behavior indexes, the life history measures, and the demographic variables were kept in the sample, leaving an analytical sample of n = 12,012. 1
The analytical sample includes participants who were about 16 years old at Wave 1 (X̄ = 15.84, SD = 1.59; min., max: 12, 18), which serves as adolescence in the current study. During Wave 3 (early adulthood) and Wave 4 (adulthood) respondents were, on average, 22 (SD = 1.67; min., max: 18, 25) and 29 (SD = 1.60; min., max: 25, 32) years old, respectively. The analytical sample was comprised of 55% of cases who self-reported as female (45% male) and 64% who self-reported their race as white (34% Person of Color). All data preparation and analyses were conducted using Stata SE 17 (StataCorp, 2021).
Measures 2
Developmental Taxonomic Groups
To identify cases that exhibited offending patterns consistent with Moffitt's taxonomic groups, a multi-step process was followed that has been used by prior researchers (Barnes et al., 2011), with minor modifications. First, items measuring both property (e.g., damaging property, shoplifting, or burglary) and violent offending (e.g., seriously injuring someone in a fight, using or threatening to use a weapon, shooting, or stabbing someone) were obtained from waves 1 to 4. While some variation from wave to wave existed in terms of the specific offending behaviors, both property and violent offending were captured at each wave. For each wave, the items were first combined to create an average score. 3 Second, each average score from each wave was dichotomized so that 0 = no offending and 1 = some offending. Third, as noted above, any cases missing on any of the three dichotomous measures (an indicator variable for waves 1 and 2, 4 an indicator variable for Wave 3, and an indicator variable for Wave 4) were removed so that the analytical sample contained only cases with valid scores on the indicator variables used to identify the taxonomic groups. Finally, cases were assigned to taxonomic groups following conventional practices (e.g., Barnes et al., 2011). More specifically, cases that scored 0 on all three indicator variables were coded as abstainers, whereas cases that scored a 1 on the adolescence indicator variable and 0 on waves 3 and 4 were coded as adolescent-limited (AL), and cases that scored a 1 on all three indicator variables were coded as life-course persistent (LCP).
Conventionally, researchers have used just the three categories above in assessing Moffitt's developmental taxonomy with the Add Health data. 5 However, there were 3,208 cases that were not assigned to any of the three groups. To avoid dropping these cases or mis-assigning them to one of the three conventional groups, we chose to use a more comprehensive group assignment strategy, as outlined in Table 1. Cases that indicated offending at waves 1 and 3, but not Wave 4, and were older than 18 years old at Wave 3 6 , were coded as a new group called AL-extended. Cases that indicated no offending in adolescence (waves 1 and 2) but indicated offending at either or both waves 3 and/or 4 were coded as post-adolescence onset offenders (all cases in this group were older than 18 years at Wave 3). Finally, cases that indicated offending in adolescence and in Wave 4, but not Wave 3, were coded as LCP offenders. Given the 5- to 6-year difference between waves 3 and 4 we considered offending at Wave 4 to represent persistence in antisocial behavior. 7
Developmental Taxonomic Group Assignment by Offending Pattern.

Note. 0 = no offending, 1 = any offending; all AL-extended and post-adolescence onset cases were older than 18 years at Wave 3; year of data collection in parentheses below the indicated wave; all three patterns of post-adolescence onset resulted in being coded as post-adolescence onset.
Life History Traits and Behaviors in Adolescence
Measures tapping various aspects of somatic effort (i.e., energy and resources that are directed towards an organism's own health and well-being) focused on three primary areas. Item selection was informed by components of life history theory and prior research that has employed the Add Health study to measure facets of life history (e.g., Brumbach et al., 2009; Nedelec & Beaver, 2012). To ensure that these measures were asked during adolescence (i.e., aged 18 years or younger), we chose to use only items from Wave 1 and not include Wave 2 items (dropping cases older than 18 at Wave 2 would result in a loss of 1,126 cases). First, physical health was based on 24 different items some of which asked respondents their level of agreement with descriptive statements (e.g., “You are physically fit”) while others asked about the frequency of experiencing various negative symptoms (e.g., “How often have you had a sore throat or cough?”). These items were recoded so that higher scores indicated better health outcomes, were standardized, and then averaged to form the physical health index (Cronbach's α = .85). Second, given its relevance to well-being, measures of mental health were also included. Eight items, some of which were based on frequency of affect-related experience (e.g., “How often in the past week were you happy?”) and some of which were based on level of agreement (e.g., “You have a lot of good qualities”), were standardized and averaged to form the positive affect index (Cronbach's α = .81). Higher scores on the positive affect index indicate more positive general feelings of well-being. Third, negative mental health was measured using 15 items based on frequency of experience (e.g., “How often in the past week did you feel depressed?”). The items were standardized and averaged to form the negative affect index and illustrated strong internal consistency (Cronbach's α = .86). Higher scores on the negative affect index indicate more negative feelings of well-being.
Another key component of life history strategy is energy and resources that an organism directs to the acquisition of mates and the production of offspring. Collectively, these behaviors are referred to as reproductive effort. Given the average age of respondents at Wave 1 (about 16 years), variability on measures related to parental investment was minimal. Thus, reproductive effort was represented in the current study by two measures. 8 First, respondents were asked “With how many people, in total, including romantic relationship partners, have you ever had a sexual relationship?”. Responses to this question were numerical and so the number of sexual partners measure is a count variable. 9 Second, respondents were asked questions pertaining to how many romantic and/or non-romantic relationships resulted in a pregnancy. In total, six items (up to three romantic partners and up to three non-romantic partners) were summed and then dichotomized to form the pregnancy measure where 0 = no pregnancies and 1 = one or more pregnancies. 10
Developmental Environment Measures
Following the direction and nomenclature of prior researchers (e.g., Brumbach et al., 2009; Nedelec & Beaver, 2012), the current study includes three measures of environmental conditions in adolescence. First, to tap mortality risk in the developmental environment, seven items pertaining to the frequency of various threatening experiences (e.g., “During the past 12 months, how often … did you see someone shoot or stab another person; did you get into a physical fight; were you jumped?”) were used to create the standardized and averaged environment harshness index (Cronbach's α = .71). 11 Higher scores on the harshness index indicate a riskier or more dangerous developmental environment. Second, to measure unreliable and unpredictable aspects of the developmental environment five items derived from Wave 3 were employed. These retrospective items were all preceded by the statement, “By the time you started 6th grade, how often…”. Examples included, “Your parents or other adult care-givers left you home alone when an adult should have been with you”, “Your basic needs were not taken care of, such as keeping you clean or providing food or clothing”, and “Did Social Services take you out of your living situation”. 12 Two items from Wave 1 which measured whether or not the respondent had always lived at their current residence and how often they ran away from home were also included. The seven items were dichotomized (0 = never/always lived at current residence; 1 = one or more times/lived in different residences), standardized, and averaged to form the environmental unpredictability index (Cronbach's α = .55) where higher scores indicate a more unreliable or unpredictable developmental environment. Finally, another core concept in life history is the adjustment of strategies based on time horizons wherein a more negative time horizon (i.e., shorter expected life span) is more likely to result in a faster life history strategy. Thus, following the direction of past researchers (e.g., Mishra & Lalumiere, 2008; Nedelec & Beaver, 2012), three items tapping future outlook from Wave 1 were included. Respondents were asked, “What are the chances the following will happen to you…You will live to age 35 (reverse coded); You will be killed by age 21; You will get HIV or AIDS”. Responses (1 = almost no chance, 2 = some chance/probably not, 3 = a 50–50 chance, 4 = a good chance, 5 = almost certain) were standardized and the three items were averaged to create the negative time horizon measure (Cronbach's α = .57) where higher scores indicate a more negative future outlook (i.e., shorter expected life span).
Demographic Measures
Three measures of demographic information were included in the multivariable regression model as control variables. Age was measured in years and was calculated by subtracting a respondent's year of birth from the interview year at Wave 1. 13 Self-reported measures of sex (0 = female, 1 = male) and race (0 = Person of Color, 1 = White) from Wave 1 were also included.
Analytical Strategy
To test the hypotheses of the current study, a multi-step approach was employed. First, descriptive statistics of all study measures were produced. Second, to assess whether the life history measures (somatic effort, reproductive effort, and developmental environment indexes) varied across the taxonomic groups, a series of ANOVA were conducted. This assessment also included tests of effect sizes using ω2 and corrected multiple pairwise comparisons using Tukey's HSD. Third, to assess the predictive capabilities of the life history measures, a single multinomial logistic regression model was conducted to predict taxonomic group membership. A multinomial regression model was chosen over a series of logistic regression models to ensure that the resulting probabilities for group membership were in reference to a single reference group rather than in reference to all the remaining groups and to avoid incorrect estimation of standard errors based on varying samples (Weisburd et al., 2020). 14 Given its treatment as a baseline offending group in Moffitt's developmental taxonomy and that it is considered the most common type of offending group, the AL taxonomic group was used as the reference group in the current study. Finally, to interpret the findings more readily, predictive probabilities were produced based on the multinomial logistic regression model.
Results
Table 2 provides the descriptive statistics of the study variables for the analytical sample (n = 12,012). As illustrated, the standardized somatic effort measures all exhibited substantial variation. As expected, given the mean age at wave 1, the reproductive effort measures were quite skewed with the mean number of sexual partners at about 1 (median = 0) and SD = 8.25, while just over 6% of the sample indicated a pregnancy in adolescence. The developmental environment measures also exhibited substantive variation but were positively skewed as well. The distribution of cases across the taxonomic groups conformed to expectations and prior observations in the Add Health and other data: most of the sample (38%) was AL offenders, followed by abstainers (29%), AL-extended (14%), life-course persistent (12%) and post-adolescence onset (7%). As noted, the analytical sample was, on average, about 16 years old at Wave 1, close to an even split in terms of gender (55% female), and mostly White in terms of self-reported race (64%).
Summary Statistics for all Study Variables and Sample Demographics (n = 12,012).

Note. “LH”: life history; “W”: Wave; Sex: 0 = female; 1 = male; Race: 0 = Person of color; 1 = White.
The unpredictability measure was created using items from both W1 and W3.
The second step in the analyses was to assess the mean differences on all study variables across the taxonomic groups. A series of ANOVA (see Table S1 in the supplemental material) illustrated that the groups differed on average across all study measures (p < .0001), although the effect sizes (ω2) were generally weak to moderate. The corrected multiple pairwise comparisons illustrated group differences on each of the variables across a substantive proportion of all the comparisons (see Tables S2a to S2d in supplemental material). These initial findings indicated that the life history measures appeared to vary across taxonomic groups and justified the multivariable regression analyses.
The third step in the analyses included a single multinomial logistic regression model wherein a categorical variable containing the five taxonomic groups was regressed on the somatic effort measures, the reproductive measures, the developmental environment measures, and the control variables. Multinomial regression is preferred when the outcome variable is categorical and the analysis seeks to assess the contrast between three or more groups (Hasking et al., 2011). As noted above, the reference category against which the groups were compared was the AL taxonomic group. Consequently, the results of the multinomial regression analysis indicate the likelihood of group classification relative to classification in the AL group holding the other variables in the model constant.
For ease of interpretation, the results of the multinomial regression model are displayed in Figure 1 which illustrates the relative risk ratio (RRR) and associated 95% confidence intervals per covariate (complete results of the regression model are displayed in Table S3 in the supplemental material). Confidence intervals that do not overlap with the vertical reference line at 1.0 on the x-axis are considered statistically significant (i.e., p < .05). As illustrated, physical health increased the odds of classification as an abstainer (RRR = 1.61) and as a post-adolescence onset offender (RRR = 1.36), relative to being an AL offender. Thus, both abstainers and post-adolescence onset offenders were more likely to report being in better physical health in adolescence than AL offenders. However, there was no difference between the odds of classification in the AL-extended or LCP groups, relative to classification as an AL offender. The second somatic effort measure, positive affect, exhibited a similar pattern where the odds of being an abstainer and the odds of being a post-adolescence onset offender were 1.23 and 1.35 times that of being an AL offender, respectively. Again, abstainers and post-adolescence onset offenders were more likely to report higher scores on the positive affect index in adolescence relative to AL offenders. The final somatic measure, negative affect, evinced a decrease in the odds of classification as an abstainer relative to an AL offender (RRR = 0.74). Thus, abstainers were less likely to report higher scores on the negative affect index in adolescence compared to AL offenders. In terms of the reproductive measures, neither illustrated any association with the odds of classification in any group, relative to the AL group.
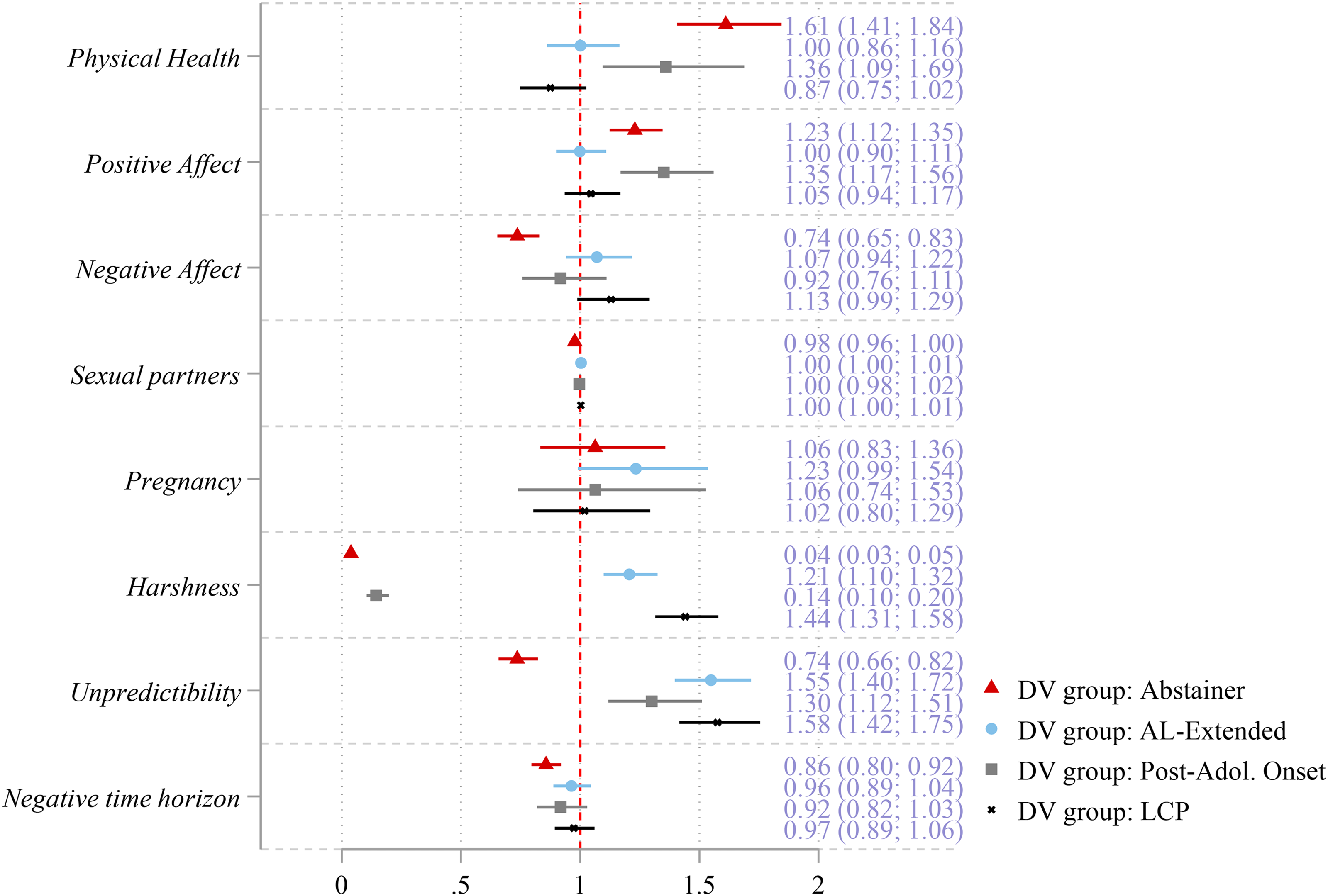
Multinomial logistic regression results predicting taxonomic group (reference group: AL offenders) using life history measures in adolescence (n = 12,012).
The three developmental environment measures were all associated with group classification, though in various ways. For instance, abstainers (RRR = 0.04) and post-adolescence onset offenders (RRR = 0.14) were much less likely to experience a harsh developmental environment relative to AL offenders. Whereas, both AL-extended (RRR = 1.21) and LCP (RRR = 1.44) groups were more likely to have experienced harsh developmental environments relative to AL offenders. Furthermore, all three offending groups (Al-extended RRR = 1.55; post-adolescence onset RRR = 1.30; LCP RRR = 1.58) were more likely to have experienced greater unpredictability in their developmental environments than AL offenders, but abstainers (RRR = 0.74) were less likely to experience unpredictability. Finally, negative time horizon evinced a decrease in the odds of being classified as an abstainer (RRR = 0.86) such that the shorter the expected lifespan a respondent perceived the less likely they were to be an abstainer than an AL offender. 15
Given the results of the multinomial logistic regression model, predicted probabilities of group classification were plotted across two of the developmental environment measures: harshness and unpredictability. These variables were chosen since they evinced the most consistent association with group membershipin the model. Figure 2 displays the predicted probabilities of classification in each taxonomic group, relative to AL offenders, across the scores on both the harshness and unpredictability indexes (from −3 SD to + 3 SD ). As illustrated in the left panel of Figure 2, as experiences of a harsh, or riskier, developmental environment increased, the likelihood of group classification, relative to AL offenders, varied for each taxonomic group. The starkest contrast illustrated are the probabilities for classification as an abstainer (relative to AL). Specifically, the probabilities change from .95 at −3 SD to .19 at the mean to less than .01 at + 3 SD . Thus, at lower ends of the harshness index, the model predicted that cases were much more likely to be classified as abstainers than AL offenders. Whereas, at the higher end of the index, the model predicted that cases were far more likely to be classified as AL offenders than abstainers.
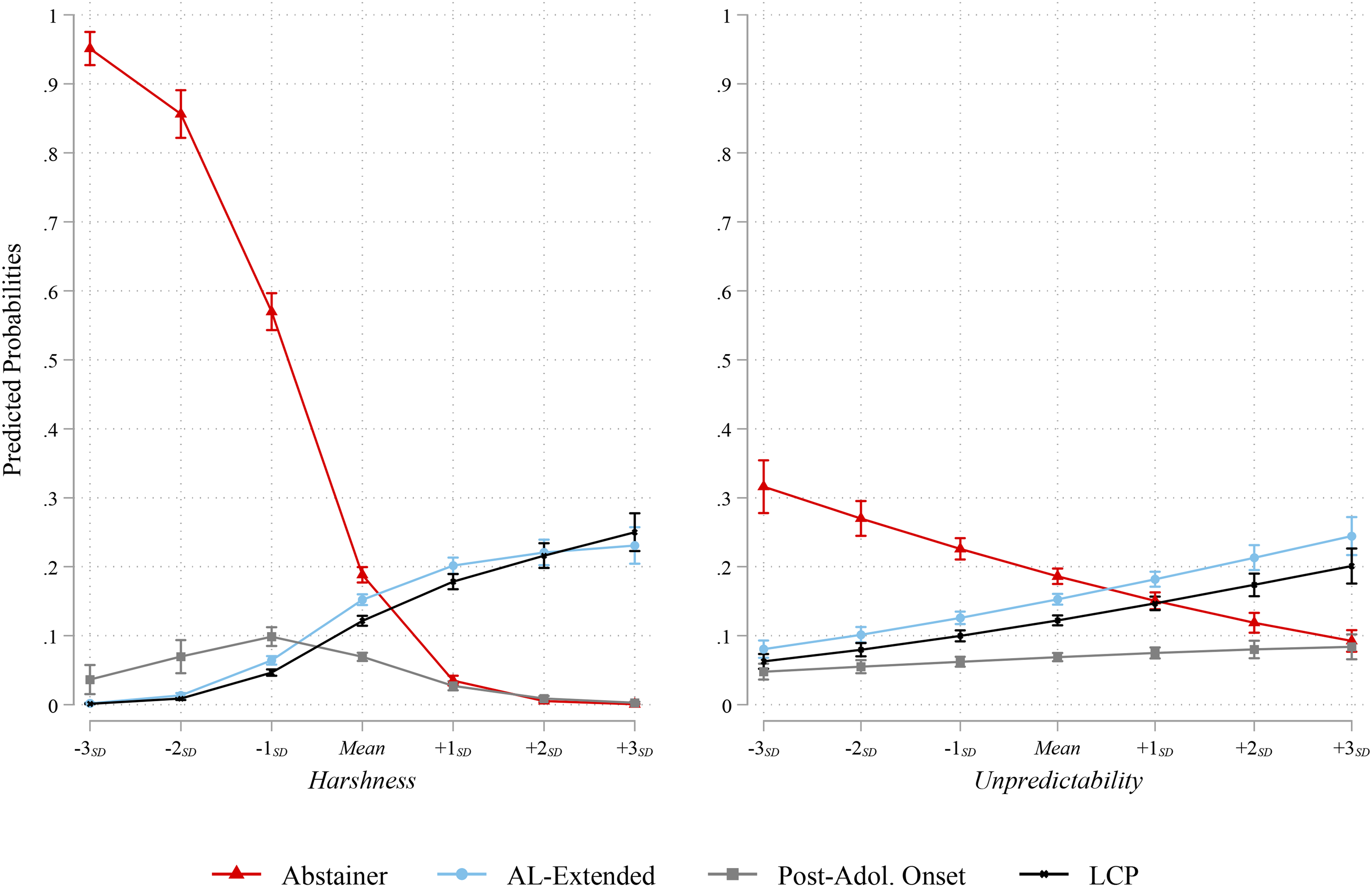
Predicted probabilities of taxonomic group membership across the developmental environment in adolescence indexes (x-axes).
The other groups evinced similar patterns to each other in terms of a steady increase in the probabilities of group classification (relative to AL offenders) from the low end of the harshness index but diverged at −1 SD . More specifically, while the AL-extended and LCP offending groups increased in a virtually identical manner across the distribution of the harshness index, the post-adolescence onset group eventually converged on the same probabilities as the abstainer group above the mean on the harshness index. Overall, these patterns illustrate that respondents who reported any offending in both adolescence and early- to mid-adulthood (i.e., waves 3 or 4) were more likely to have experienced risky or harsh developmental environments in adolescence relative to those whose offending was limited to just adolescence or just periods in adulthood.
While not as drastic as probabilities across scores on the harshness index, the predicted probabilities across the unpredictability index also illustrated variability in terms of group classification. Once again, those who were predicted by the model to be classified as abstainers were less likely to experience an unpredictable developmental environment relative to AL offenders (as the unreliability of the developmental environment increased, the likelihood of being an abstainer—relative to an AL offender—decreased substantially). Furthermore, the AL-extended and LCP groups evinced similar patterns of predicted probabilities such that the greater the unreliability of the developmental environment the greater the likelihood of classification as an AL-extended or LCP group (relative to AL offenders). Finally, although the unpredictability index indicated an increase in the odds of classification as a post-adolescence onset offender in the multinomial regression model the predicted probabilities appear to be relatively stable across the distribution of the index, though there is a slight increase from a predicted probability of .06 at −3 SD to .20 at + 3 SD .
Discussion
Evolutionary criminology can help both traditional and contemporary approaches to crime and criminality in a variety of ways. Furthermore, an infusion of criminological understanding of antisocial behavior can also enhance evolutionary perspectives on human behavior. One area in which the consilience of theoretical approaches has taken place is in the recently developed evolutionary taxonomy: a theoretical scaffold for Moffitt's developmental taxonomy based primarily on the tenants of life history. While recent tests of the evolutionary taxonomy have provided some initial support, absent from the literature—and key to the approach's utility—is an assessment of how well the evolutionary taxonomy predicts classification into the offending groups derived from Moffitt's theory. The current study partially addressed this gap in the literature by examining how well measures of somatic effort, reproductive effort, and components of the developmental environment in adolescence predicted offending group classification using a large sample of youth who were followed into mid-adulthood. Overall, the findings provide some initial tentative support for the evolutionary taxonomy though some predictions were not supported.
First, in general, the results indicate that measures representative of a faster life history strategy (e.g., less somatic effort, greater unpredictability and harshness in the developmental environment) were predictive of more offending behaviors overall. Save for the reproductive measures, each life history measure predicted classification into the abstainer group such that abstainers were more likely to have reported more somatic effort in adolescence, more predictable and less harsh developmental environments, and a more positive future outlook (i.e., positive time horizon). Moffitt's (1993) original characterization of abstainers painted a picture of awkwardness and early experiences of isolation from peers. Indeed, Moffit argued that the most central explanation to the abstainer (non-) offending pattern was that they were excluded from peer groups due to their unattractive personality traits or underdeveloped biological maturity (Chen & Adams, 2010; Moffitt, 1993). While the current study didn’t assess the extent to which variation in personality affected group classification, the findings did illustrate that, on average, abstainers reported a more stable and safe developmental environment than those who reported any offending. In a recent review, Moffitt (2018) summarized findings indicating that while abstainers experienced isolation during adolescence they later reported positive outcomes such as group acceptance and socio-emotional coping. Perhaps the more stable environments reported in the current analyses influence both the non-offending and the more positive outcomes after adolescence. Future research is encouraged to focus on this possibility.
In terms of the taxonomic group that tends to receive the most attention in the literature—LCP offenders—only the developmental environment measures of harshness and unpredictability predicted classification as LCP, relative to AL. This observation conforms not only to Moffitt's (1993, 1994) original theoretical predictions it also aligns with much of the empirical literature that has tested her ideas (see Moffitt, 2018). Additionally, it adds further support for the evolutionary taxonomy's assertion that LCP offending patterns are a node in the adaptive strategy repertoire characterized by a fast life strategy. However, the lack of predictive utility of the other life history measures for LCP group classification—relative to AL classification—indicates a lack of support for the evolutionary taxonomy and counters prior examinations. One possible explanation for why the current study conflicts with prior studies is that while prior studies of the evolutionary taxonomy have examined how specific aspects of a fast life history (e.g., number of sexual partners) predict classification as an LCP they have done so in reference to all cases who were not LCP (e.g., Barnes et al., 2011). The analysis in the current study, however, was more isolated in terms of the comparisons given that all group classification was in reference to just AL offenders. As noted in the Analytical Strategy section, this limit was intentional to ensure the use of a consistent reference category and sample. Our contention is that the analytical approach in the current study is more methodologically appropriate and therefore future research is encouraged to employ similar approaches in assessing the predictions of offending group classification.
A final observation centers on our choice of operationalization of the offending groups. As outlined in Table 1, we employed a more comprehensive identification of offending taxonomies relative to prior researchers who have employed the Add Health data. This comprehensive approach resulted in the addition of two offending groups beyond AL and LCP: the AL-extended group and the post-adolescence onset group. The identification of groups beyond Moffitt's dual-taxonomy has been observed in other samples (e.g., Wiesner & Capaldi, 2003). Additionally, prior researchers using the Add Health (e.g., Barnes et al., 2011) have recognized the existence of cases that weren’t neatly classified into one of the abstainer, AL, or LCP groups. It would further appear that the identified groups differ in terms of their potential etiological factors. For example, while the AL-extended group (cases that offended in adolescence and in Wave 3 but were older than 18 at Wave 3) mimicked the LCP group in terms of the associations in the model (indeed, the 95% confidence intervals for each point estimate for each covariate overlapped between the two groups), the post-adolescence onset group aligned with the associations between the somatic effort measures and the abstainer group but were mixed in terms of the developmental environment measures. The starkest contrast appeared in terms of the association with unpredictability which was negatively associated with the odds of being classified as an abstainer but was positively associated with the odds of being classified as post-adolescence onset. To the extent that these observations are unique to the Add Health remains an empirical question, but authors are encouraged to employ comprehensive group operationalizations when assessing Moffitt's developmental taxonomy to avoid masking potentially meaningful variation and etiological examinations.
Limitations
The observations of the current study should be tempered by at least four limitations. First, the measures used to tap aspects of life history, though related, were not included in the Add Health study to specifically measure components of life history strategies. Additionally, several domains of life history were not included in the analyses due to a lack of items in the data. Future research is encouraged to assess the results of the current study with broader inclusion of life history measures to further test the evolutionary taxonomy. Second, although the group classification strategy employed in the current study was meant to be more comprehensive than prior studies using the Add Health, it may lack the precision of other methods used to identify groups (e.g., latent class or group-based trajectory modeling). It remains an empirical question whether the results of the current study would vary if these other more sophisticated methods were employed. Third, although confidence in the causal order is increased as a function of the longitudinal nature of the data those measures that ostensibly are external to the respondent (i.e., developmental environment) may in fact not be, due to the self-reported nature of the data. Thus, the findings may be confounded by common source bias (Nielsen, 2019). Future research is encouraged to employ multiple-source data to further test the predictive utility of life history measures on offending group classification. Finally, virtually every measure included in the current study has been shown to have non-zero heritability in various studies (e.g., Barnes et al., 2011; Polderman et al., 2015). Thus, to the extent that genetic factors impact any of the observed associations confounding is a potential concern. Future research is encouraged to assess the predictions of the evolutionary taxonomy employing genetically informed methodologies (e.g., Boutwell et al., 2015b).
Conclusion
With the aforementioned limitations in mind, the current study has illustrated a potential way in which evolutionary criminology can help inform and expand traditional criminological approaches. Of additional importance is the way in which evolutionary approaches can benefit from an infusion of criminological concepts like Moffitt's developmental taxonomy. While the current study represented the first substantive test of the evolutionary taxonomy's predictive utility, it certainly represents an initial modest step. Future research is encouraged to further examine the tenets of the evolutionary taxonomy and evolutionary criminology, in general, using disparate data and methods that address the limitations outlined herein. Our understanding of crime, criminality, and antisocial behavior will only benefit.
Supplemental Material
sj-docx-1-evp-10.1177_14747049231212356 - Supplemental material for Predicting Moffitt's Developmental Taxonomy of Antisocial Behavior Using Life History Theory: A Partial Test of the Evolutionary Taxonomy
Supplemental material, sj-docx-1-evp-10.1177_14747049231212356 for Predicting Moffitt's Developmental Taxonomy of Antisocial Behavior Using Life History Theory: A Partial Test of the Evolutionary Taxonomy by Joseph L. Nedelec and Francesco DiRienzo in Evolutionary Psychology
Footnotes
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article
Supplemental Material
Supplemental material for this article is available online.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.