Nephrolithiasis is a major global health challenge, with oxidative stress and mitochondrial dysfunction emerging as key drivers of renal injury and stone formation. l-ergothioneine (l-Erg), a naturally occurring antioxidant transported by OCTN1, has shown promising effects in cystinuria models, preventing stone formation. Despite evidence supporting an indirect mechanism of action, key mechanistic aspects have yet to be fully clarified. This study aimed to evaluate whether l-Erg can prevent stone progression in cystinuria and in other types of lithiasis, such as calcium oxalate nephrolithiasis, and to further elucidate its mechanistic basis.
Results:
Using mouse models, l-Erg significantly reduced cystine stone growth and renal inflammation, and its combination with d-penicillamine enhanced stone dissolution and mitigated drug-related toxicity. In calcium oxalate nephrolithiasis, l-Erg decreased crystal deposition, preserved renal architecture, normalized glutathione levels, and restored mitochondrial respiration. Transcriptomic analysis revealed downregulation of immune pathways and activation of cell cycle genes, suggesting attenuation of inflammation and promotion of tubular repair.
Innovation:
This study is the first to demonstrate that l-Erg exerts renoprotective effects through combined antioxidant and mitochondrial mechanisms in two major forms of nephrolithiasis and introduces a dual therapeutic approach combining an antioxidant with a cystine-solubilizing agent.
Conclusion:
By targeting oxidative stress and mitochondrial dysfunction, l-Erg represents a promising therapeutic strategy for nephrolithiasis, either alone or as an adjunct to current treatments. Antioxid. Redox Signal. 44, 878–891.
Nephrolithiasis represents a major global health challenge, with increasing prevalence and high recurrence rates that impose a significant burden on health care systems and patient quality of life (Abufaraj et al., 2021; Youssef et al., 2020). Among the different stone types, cystinuria stands out as one of the most severe forms, associated with early onset, lifelong recurrence, and progressive renal damage (Modersitzki et al., 2014; Streeper et al., 2017). This rare genetic disorder, caused by mutations in SLC3A1 and SLC7A9 genes, disrupts the reabsorption of cystine and dibasic amino acids in the renal proximal tubule (Calonge et al., 1994; Feliubadaló et al., 1999), leading to excessive urinary cystine excretion. Because cystine has low solubility at physiological urine pH, it precipitates and aggregates into crystals, which progressively form stones in the urinary tract (Königsberger et al., 2000). Clinical data indicate that up to 75% of patients with cystinuria develop bilateral calculi, and repeated episodes of obstruction and surgical interventions often result in chronic kidney disease (CKD) and hypertension (Prot-Bertoye et al., 2015; Strologo et al., 2002). Health-related quality of life is markedly impaired in cystinuria compared with other stone formers, underscoring the need for improved therapies.
Innovation
This study introduces l-Erg as a novel therapeutic strategy for nephrolithiasis by targeting oxidative stress and mitochondrial dysfunction, two central mechanisms in stone pathophysiology. We demonstrate that l-Erg alone can reduce stone growth in cystinuria models and, when combined with DP, achieves an additive effect. Furthermore, in CaOx nephrolithiasis, l-Erg decreases crystal deposition and preserves renal architecture. Mechanistically, l-Erg normalizes glutathione homeostasis, restores antioxidant enzyme expression, and improves mitochondrial respiration. These findings reveal a dual mechanism of renoprotection and support the clinical potential of l-Erg for diverse forms of nephrolithiasis.
Current management strategies aim to reduce cystine precipitation through conservative measures such as increased hydration, dietary modifications, and urine alkalinization (Servais et al., 2021). When these approaches fail, thiol-based drugs like d-penicillamine (DP) and tiopronin are prescribed to dissolve stones by forming mixed disulfides with cysteine, which are more soluble than cystine (Halperin et al., 1981; Pak et al., 1986). However, these treatments are frequently discontinued due to severe adverse effects, including dermatological, hematological, and autoimmune complications (Prot-Bertoye et al., 2019). This therapeutic gap underscores the urgent need for safer and more effective alternatives.
Beyond cystinuria, oxidative stress and mitochondrial dysfunction have emerged as common features in kidney stone pathophysiology, including calcium oxalate (CaOx) lithiasis, the most prevalent stone type worldwide (Cao et al., 2004; Chaiyarit and Thongboonkerd, 2020; Hirose et al., 2010; Xu et al., 2025, 2024). Crystal interaction with tubular epithelial cells triggers reactive oxygen species (ROS) production, lipid peroxidation, and mitochondrial damage, which induce inflammation and apoptosis (Alelign and Petros, 2018). These processes not only compromise renal homeostasis but also create a favorable environment for crystal nucleation and aggregation. Therefore, targeting oxidative and mitochondrial defects represents a promising strategy to prevent stone formation and progression (Liu et al., 2023; Marhoume et al., 2021; Niimi et al., 2014; Sharma et al., 2016; Wu et al., 2021).
l-ergothioneine (l-Erg) is a naturally occurring thiol antioxidant obtained from the diet and actively transported into tissues by OCTN1, a transporter highly expressed in the kidney (Gründemann et al., 2022). l-Erg accumulates in organs under oxidative stress and has demonstrated cytoprotective, antioxidant, and anti-inflammatory effects in renal tissue across mouse models of CKD, diabetic nephropathy, and drug-induced nephrotoxicity (Dare et al., 2021; Deiana et al., 2004; Peng et al., 2025; Salama et al., 2021). Its unique properties, long half-life, ability to scavenge ROS and reactive nitrogen species, and metal-chelating capacity make it an attractive candidate for renal protection (Akanmu et al., 1991; Aruoma et al., 1997; Paul and Snyder, 2010; Stoffels et al., 2017). In a previous study, we showed that l-Erg prevents cystine lithiasis formation or delays its onset in Slc7a9−/− cystinuria mouse model by increasing urinary cystine solubility and restoring mitochondrial respiratory capacity (Mayayo-Vallverdú et al., 2023). In addition, the loss of OCTN1 in a cystinuria mouse (Slc7a9−/− Slc22a4−/−) resulted in a significant increase in stone development, highlighting l-Erg’s protective role in cystine lithiasis (de Heredia et al., 2021). Moreover, when the double knockout (KO) model (Slc7a9−/− Slc22a4−/−) was treated with l-Erg, no effect on stone formation was observed, further confirming that l-Erg internalization is essential for its stone-preventing effect (Mayayo-Vallverdú et al., 2023). These findings indicate that l-Erg does not act directly in the urine but rather exerts its beneficial effects at the intracellular level as an antioxidant, cytoprotective, and anti-inflammatory molecule. However, the precise mechanisms underlying its protective effect and its potential applicability in other lithiasis in which oxidative stress, mitochondrial function, and renal homeostasis are altered beyond cystinuria remain unclear.
Oxidative stress and mitochondrial dysfunction are central contributors to stone pathophysiology. Therefore, correcting these defects represents a promising therapeutic strategy. In this context, the present study was designed to explore the therapeutic potential of l-Erg in kidney stone disease. Specifically, we aimed to assess its effect in cystinuria models once the stone is formed, alone and in combination with DP, and to investigate whether its benefits could be extended to CaOx lithiasis. Furthermore, we sought to elucidate the molecular pathways underlying l-Erg renoprotection, focusing on mitochondrial function, redox signaling, and transcriptomic changes.
Results
Combined l-Erg and d-penicillamine therapy halts stone progression and sustains effect post-treatment
We first assessed the efficacy of l-Erg, DP, and the combined l-Erg + DP treatment in mitigating the progression of cystine stones. Daily stone growth rate was assessed by in vivo X-ray imaging at baseline, during treatment, and after discontinuation. Figure 1A shows that, although no significant differences were observed between groups prior to treatment (pretreatment), all three interventions significantly slowed stone progression compared with controls. l-Erg administration decreased stone growth by more than 50% (from 2 to 0.77 mg/day), while DP and the combined treatment successfully halted stone progression or even reduced stone size in some mice (from 1.6 to −0.17 mg/day and from 2.1 to −0.25 mg/day, respectively) (Fig. 1A). Although differences between DP alone and the combined treatment were not statistically significant, a trend toward an additive effect was observed. Similar results were obtained in both Slc7a9−/− and Slc3a1D140G cystinuria mouse models (Supplementary Fig. S1), indicating that the treatment effect was independent of the underlying genetic defect.
Treatment effect on stone progression, cystine precipitation, and urinary cystine levels.(A) Timeline of treatment schedule and stone growth rate in control, l-Erg-treated, DP-treated, and l-Erg + DP-treated mice during 1 month before treatment (PreTreatment) and 1 month of treatment (Treatment). Gray dashed lines connect measurements from the same mouse across time points. (B) Timeline of treatment schedule and stone growth rate in S-Met-treated, l-Erg-treated, DP-treated, and l-Erg + DP-treated mice during PreTreatment, Treatment, and 1 month after treatment discontinuation (PostTreatment). Gray dashed lines indicate individual mouse trajectories. (C) Relative cystine precipitate in urine from control, S-Met-treated, l-Erg-treated, DP-treated, and l-Erg + DP-treated mice. Values normalized to control levels. (D) Urinary cystine levels in control, l-Erg-treated, DP-treated, and l-Erg + DP-treated mice. Cystine concentration normalized by creatinine. Each dot represents an individual mouse. #p < 0.1, *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001. Cntrl, control; l-Erg, l-ergothioneine; DP, D-penicillamine; S-Met, S-methyl- l-ergothioneine; 3M, 3 months of age; 4M, 4 months of age; 5M, 5 months of age; 6M, 6 months of age.
As l-Erg has a pharmacokinetic half-life of about 1 month (Tang et al., 2018), we conducted a second cycle of treatment to assess whether its therapeutic effects persisted after discontinuation (Fig. 1B). We also evaluated whether the main metabolite of l-Erg, S-Met, has any effect on stone progression, but no differences were observed between pre-treatment, treatment, and post-treatment conditions (Fig. 1B). Regarding l-Erg, DP, and l-Erg + DP treatments, comparisons between pre-treatment and treatment confirmed our previous results, with stone growth rates decreasing from 3.34 to 1.72 mg/day (l-Erg), from 1.47 to −0.22 mg/day (DP), and from 2.36 to 0.16 mg/day (l-Erg + DP) (Fig. 1B). Interestingly, in mice treated with l-Erg and l-Erg + DP, the beneficial effects persisted for at least 1 month after treatment discontinuation, whereas DP-treated mice exhibited a post-treatment stone growth rate similar to pre-treatment levels (Fig. 1B). These findings indicate that l-Erg prolongs the effect of DP, reducing stone size even after therapy cessation. These results indicate that l-Erg enhances cystine solubility and prolongs DP efficacy through sustained antioxidant activity.
In our previous study, we reported that although urinary cystine concentration in l-Erg-treated mice was comparable with controls, l-Erg reduced cystine precipitate by 50% (Mayayo-Vallverdú et al., 2023). To compare treatment outcomes in the current study, we measured cystine precipitation and urinary cystine levels under different treatment conditions (Fig. 1C, D). In l-Erg-treated mice, cystine precipitate decreased by more than 60%, with no significant changes in urinary cystine levels compared with controls (Fig. 1C, D), confirming previous observations. DP-treated mice exhibited a greater reduction in cystine precipitate (>70%) and significantly lower urinary cystine levels compared with controls, consistent with its known cystine-chelating capacity (Fig. 1C, D). Notably, in l-Erg + DP-treated mice, an additive effect was evident, with cystine precipitate decreasing by more than 80% (Fig. 1C). Regarding S-Met treatment, although higher levels of S-Met were achieved in urine after 1 month of treatment (Supplementary Fig. S2), no differences were observed in stone growth or cystine precipitate, and therefore, its potential role in cystinuria was excluded (Fig. 1B, C).
l-Erg treatment ameliorates renal injury
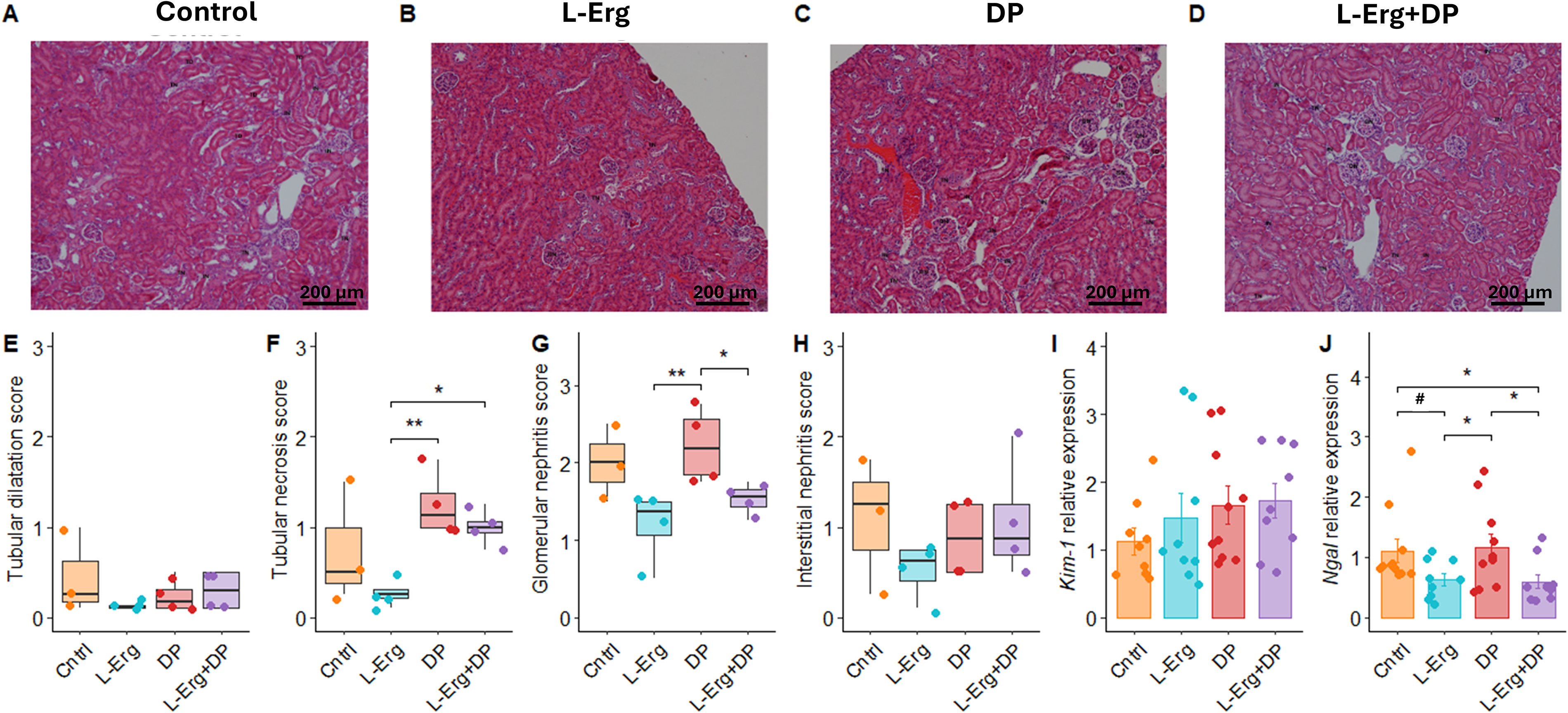
Renal physiology is affected by the presence of cystine crystals and stones, as well as by the harmful effects of DP, such as nephrotic syndrome and membranous glomerulonephritis (Habib et al., 2006). In cystinuria mice treated with DP, glomerular alterations were also observed (Font-Llitjós et al., 2007). To investigate these impacts, we evaluated histological parameters including tubular dilatation, tubular necrosis, glomerular nephritis, and interstitial nephritis (Fig. 2). l-Erg-treated mice showed a better-preserved renal morphology, with significant improvement in tubular necrosis and glomerular nephritis compared with DP-treated mice (Fig. 2E–H). In addition, l-Erg administration ameliorated glomerular nephritis in the l-Erg + DP group, counteracting a known side effect of DP treatment (Fig. 2G) (Font-Llitjós et al., 2007).
l-Erg alleviates renal injury caused by cystine crystals and DP toxicity. Representative H&E-stained kidney sections from control (A), l-Erg-treated (B), DP-treated (C), and l-Erg + DP-treated (D) groups. Histological damage score for tubular dilatation (E), tubular necrosis (F), glomerulonephritis (G), and interstitial nephritis (H), rated as 0 = no injury, 1 = <25%, 2 = <50%, and 3 = >50% injury. Renal expression of Kim-1(I) and Ngal(K) assessed by qPCR. Gene expression normalized to Gapdh and expressed as fold change relative to controls. Each dot represents an individual mouse. #p < 0.1, *p < 0.05, **p < 0.01. TD, tubular dilatation; TN, tubular necrosis; GN, glomerular nephritis; IN, interstitial nephritis.
Furthermore, we assessed the expression of kidney injury molecule-1 (Kim-1) and neutrophil gelatinase-associated lipocalin (Ngal), two biomarkers of renal injury (Fig. 2I, J). While no differences were detected in Kim-1 expression, a significant decrease in Ngal expression was observed in the l-Erg and l-Erg + DP groups compared with both control and DP-treated mice. Ngal appears to be a more robust biomarker in urolithiasis (Lasota et al., 2023), as Kim-1 shows more heterogeneous results among studies, which may explain the differences observed in our model. Altogether, these findings highlight the capacity of l-Erg to alleviate renal damage and mitigate DP-related toxicity.
l-Erg treatment modulates the inflammatory response and activates cell cycle gene expression
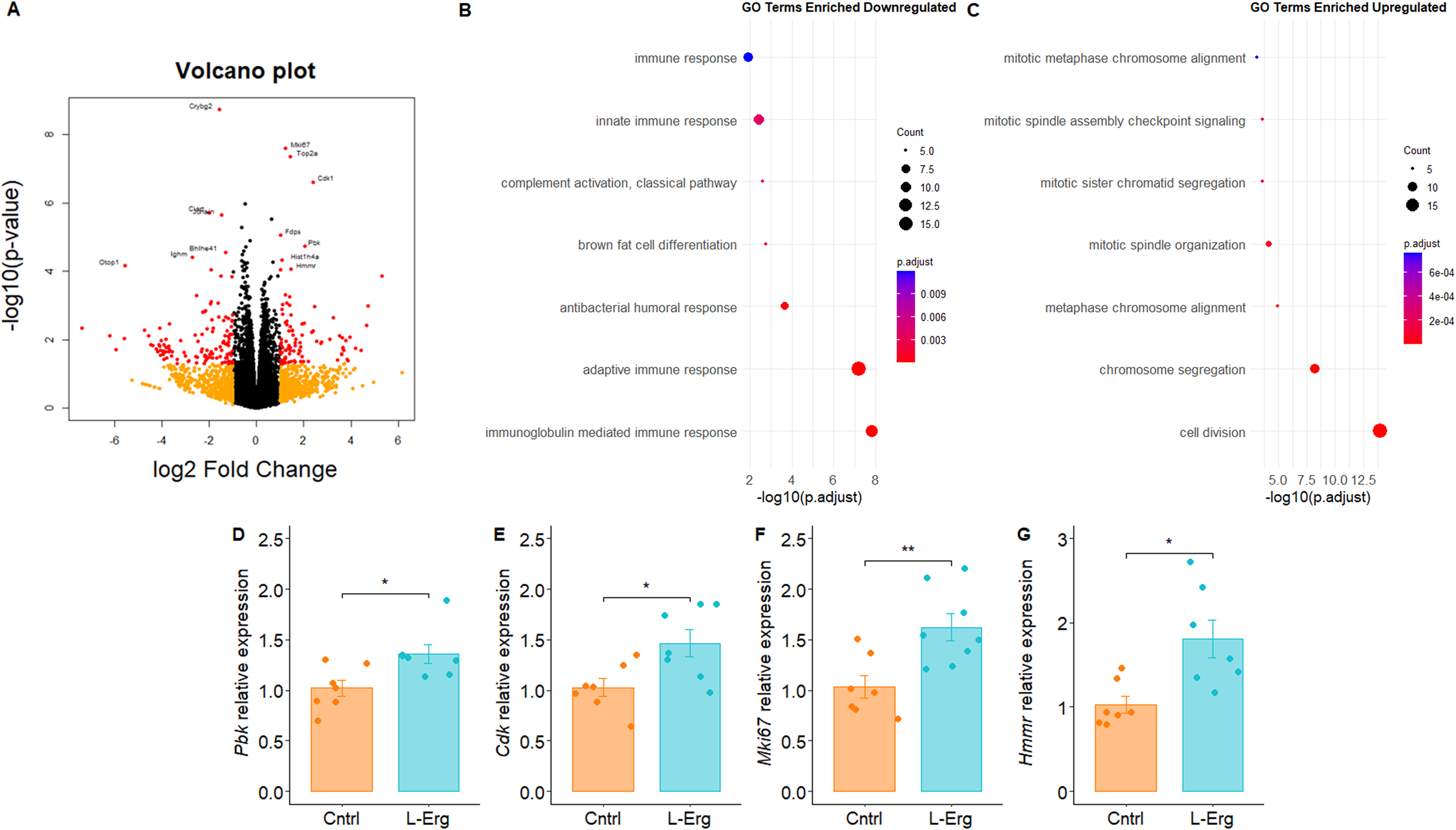
To provide new insights about the mechanism by which l-Erg alleviates renal damage and reduces cystine kidney stones, we performed RNA sequencing analysis to compare gene expression profiles between l-Erg-treated and untreated cystinuric kidneys. Differentially expressed genes (DEGs) were considered when p values were <0.05 and |log2 fold change| >1. Applying these thresholds, we identified 125 downregulated and 96 upregulated genes (Fig. 3A). Using the Database for Annotation, Visualization and Integrated Discovery (DAVID), we assessed enriched biological pathways. For the downregulated genes found in l-Erg-treated kidneys, the top enriched biological processes were related to immune response (Fig. 3B), whereas for the upregulated genes, the most enriched processes were associated with cell cycle and division (Fig. 3C). Transcriptomic analysis indicates that l-Erg modulates immune pathways (consistent with previous reports; Paul and Snyder, 2010) and promotes tubular repair via cell cycle activation.
RNA-seq analysis of kidneys from untreated and l-Erg-treated mice.(A) Volcano plot showing differentially expressed genes (DEGs). Genes with p < 0.05 and |log2 fold change| >1 are shown in red; genes with p > 0.05 and |log2 fold change| >1 in orange; and non-DEGs in black. Functional enrichment analysis of downregulated (B) and upregulated (C) DEGs using Gene Ontology (GO) terms. Dot plots display the top seven enriched GO categories based on adjusted p value calculated by DAVID. Dot size indicates the number of genes per term; color represents statistical significance (–log10-(p-adjust)). (D–G) Validation of selected DEGs by qPCR. Gene expression normalized to Gapdh and expressed as fold change relative to controls. *p < 0.05, **p < 0.01.
When focusing on the top 10 upregulated genes (Supplementary Table S1), we identified Mki67, a well-known proliferation marker (Pieters et al., 2019), as well as the kinases Cdk1 and Pbk, whose loss of function has been linked to kidney injury (Wei et al., 2025) and human kidney stone disease (Nettuwakul et al., 2020), respectively. In addition, Hmmr (hyaluronan-mediated motility receptor) was upregulated, a gene involved in cell growth and inflammation regulation (Tavianatou et al., 2019). These four DEGs identified in the RNA-seq analysis were validated by qPCR (Fig. 3D–G). Similarly, other studies investigating l-Erg in liver disease have reported attenuation of oxidative damage, inflammation, and apoptosis (Lv et al., 2024), supporting the pleiotropic protective role of l-Erg.
l-ergothioneine mitigates calcium oxalate-induced renal injury via redox and mitochondrial regulation
This section evaluated whether l-Erg can counteract acute CaOx-induced oxidative and mitochondrial injury. In previous studies (Mayayo-Vallverdú et al., 2023), we described that l-Erg does not interact with cystine or decrease urinary cystine levels to prevent cystine stones, indicating that its mechanism of action is indirect. This evidence opens up a range of possibilities for the potential effect of l-Erg in other types of lithiasis, such as CaOx lithiasis, the most common type, in which oxidative, inflammatory, and mitochondrial defects have been widely described (Xu et al., 2025, 2024). To explore this hypothesis, l-Erg (16 mg/kg/day) was administered to wild-type (WT) mice starting 1 week before CaOx crystal induction by daily glyoxylate injections (100 mg/kg/day for 5 consecutive days) and continued until the last day of injections.
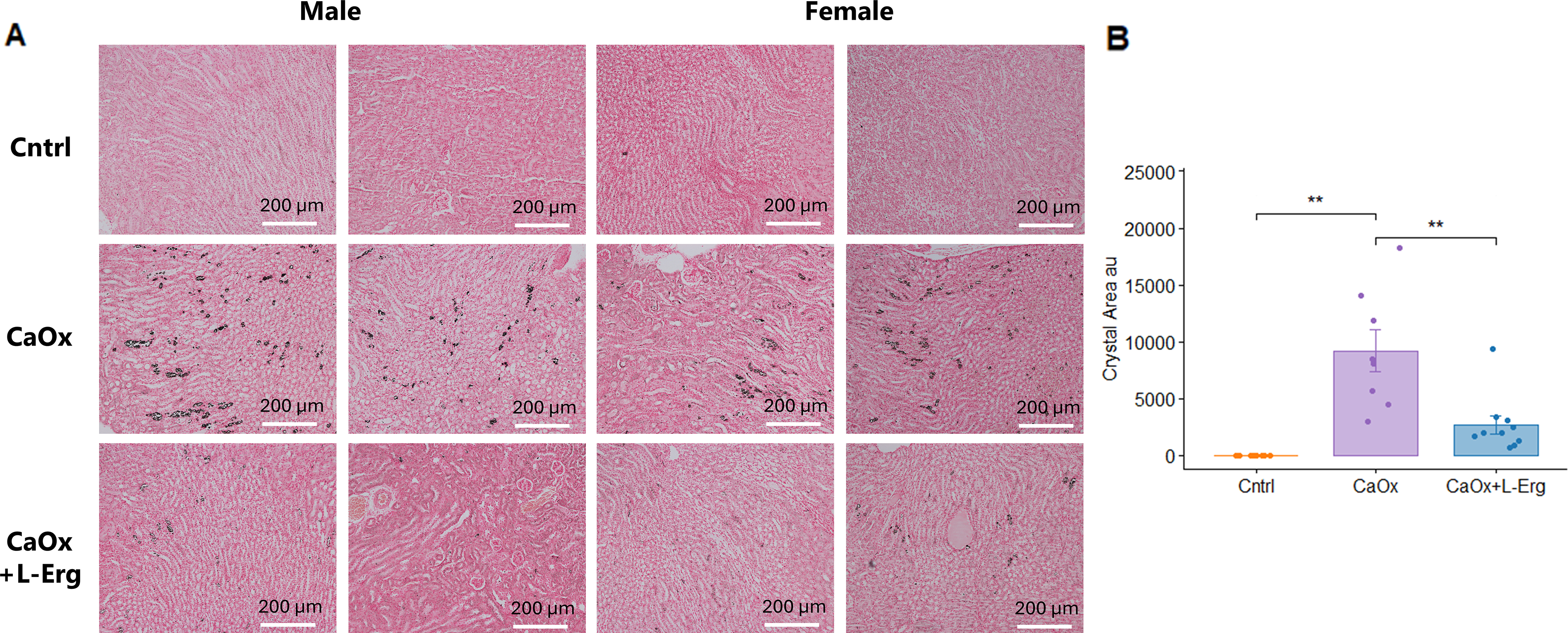
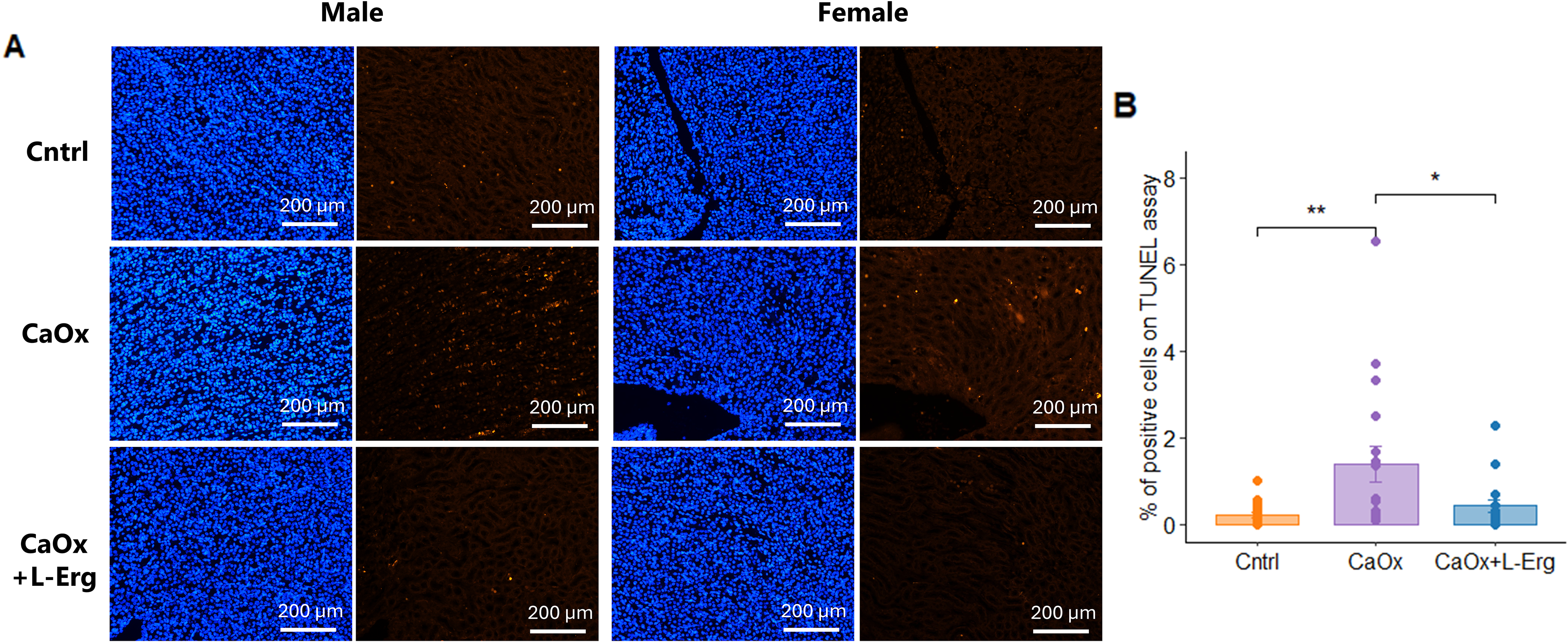
Histological analyses confirmed CaOx crystal formation and revealed that l-Erg reduced crystal burden and tubular injury. First, Von Kossa staining confirmed CaOx crystal formation after glyoxylate injections and revealed that l-Erg treatment reduced CaOx crystal deposition by more than 50% (Fig. 4A, B). Next, Hematoxylin & Eosin (H&E) staining showed that CaOx crystals induced severe tubular lesions, characterized by dilatation and cell injury compared with controls (Fig. 5A–E). l-Erg treatment significantly decreased tubular dilatation and necrosis (Fig. 5A–C). In addition, CaOx crystal induction markedly increased Kim-1 and Ngal expression, which was notably attenuated by l-Erg treatment (Fig. 5F, G). Finally, TUNEL assay confirmed that CaOx crystals triggered renal apoptosis, and l-Erg treatment mitigated this damage (Fig. 6A, B). Altogether, these findings demonstrate that l-Erg treatment reduces CaOx crystal burden and preserves renal homeostasis.
l-ergothioneine treatment reduces CaOx crystal formation in mouse kidneys.(A) Representative Von Kossa-stained kidney sections from Cntrl, CaOx-induced (CaOx), and CaOx-induced mice treated with l-Erg (CaOx + l-Erg). (B) Quantification of mean crystal area across four kidney sections per mouse. Each dot represents an individual mouse (**p < 0.01). CaOx, calcium oxalate.
l-ergothioneine treatment prevents renal damage after CaOx crystal induction.(A) Representative H&E-stained kidney sections from Cntrl, CaOx-induced mice, and CaOx-induced mice treated with l-Erg (CaOx + l-Erg). Quantification of histological damage scores for tubular dilatation (B), tubular necrosis (C), interstitial nephritis (D), and glomerular damage (E). Four kidney sections per mouse were analyzed; scores were rated as 0 = no injury, 1 = <25%, 2 = <50%, and 3 = >50% injury. Renal expression of Kim-1(F) and Ngal(G) genes assessed by qPCR. Gene expression normalized to Gapdh and expressed as fold change relative to control mice. Each dot represents an individual mouse. *p < 0.05, **p < 0.01, ***p < 0.001.
l-ergothioneine treatment reduces apoptosis induced by CaOx crystals.(A) Representative TUNEL-stained kidney sections from Cntrl, CaOx-induced mice, and CaOx-induced mice treated with l-Erg (CaOx + l-Erg). (B) Quantification of TUNEL-positive cells across four kidney sections per mouse. Each dot represents an individual mouse. *p < 0.05, **p < 0.01.
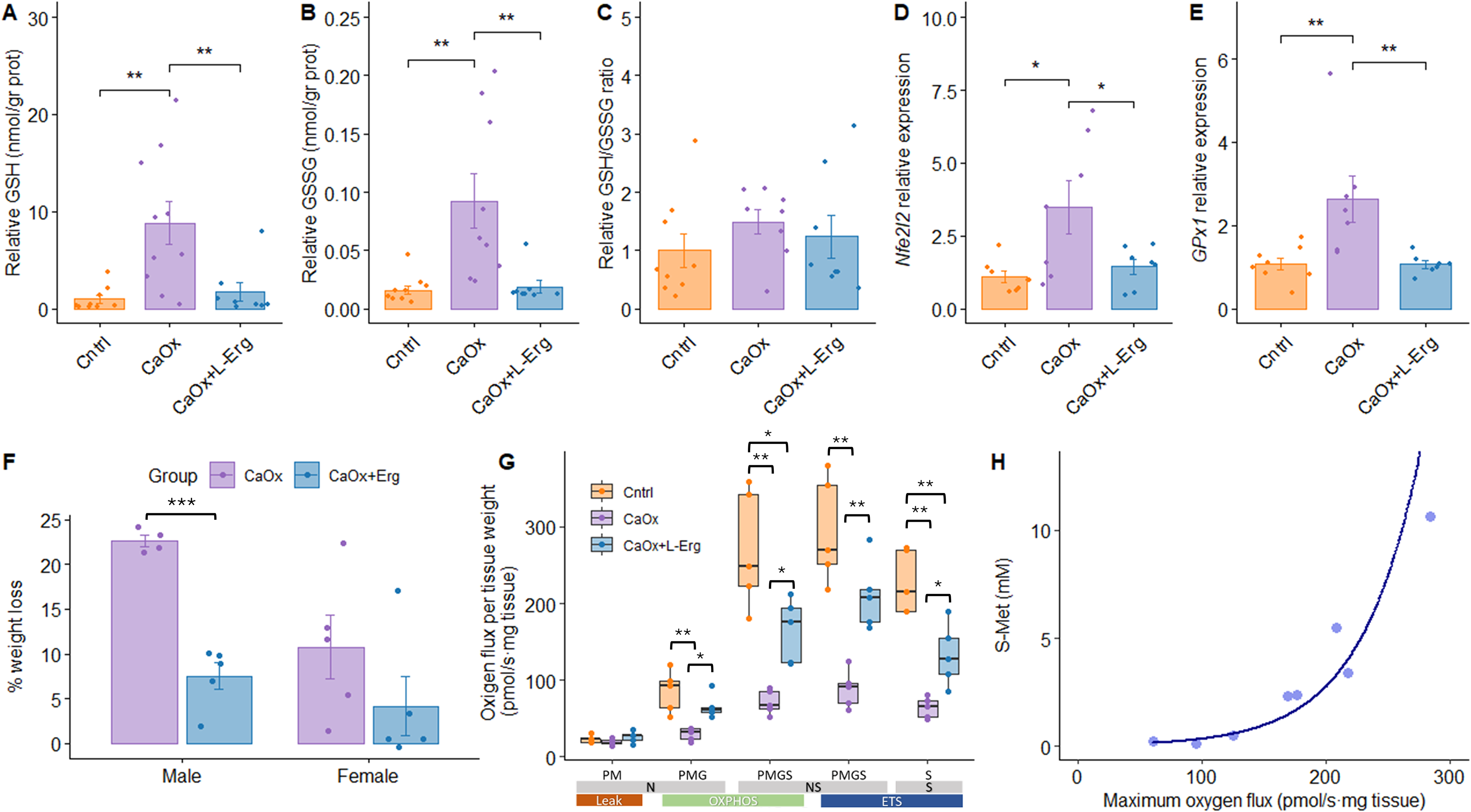
As shown in Supplementary Figure S3, 12 days of l-Erg administration were sufficient to increase urinary l-Erg and S-Met levels, indicating saturation of l-Erg uptake and activation of its metabolism. To interpret these findings, we next examined whether changes in glutathione homeostasis paralleled alterations in mitochondrial function. Assessment of kidney glutathione levels revealed that, in the CaOx group, both reduced (GSH) and oxidized (GSSG) forms were significantly increased compared with controls and CaOx + l-Erg mice, while the GSH/GSSG ratio remained similar across all groups (Fig. 7A–C). Considering that GSH synthesis is tightly regulated by antioxidant transcriptional pathways, we next evaluated the expression levels of the nuclear factor erythroid 2-related factor 2 (Nfe2l2—Nrf2) and glutathione peroxidase 1 genes, which catalyze the oxidation of GSH to GSSG. Both genes were upregulated in the CaOx group, and l-Erg treatment could restore its expression to control levels (Fig. 7D, E). These results suggest that 5 days of glyoxylate injections induce oxidative stress sufficient to trigger GSH biosynthesis, but this compensatory increase is unlikely to be sustained, eventually leading to glutathione depletion. l-Erg treatment mitigates this response, normalizing glutathione levels to control values by attenuating oxidative damage and reducing compensatory GSH synthesis. Glyoxylate induced significant weight loss, which was attenuated by l-Erg treatment (Fig. 7F).
l-ergothioneine treatment restores redox balance, mitigates weight loss, and improves mitochondrial function after CaOx crystal induction. Kidney intracellular levels of GSH (A), GSSG (B), and GSH/GSSG ratio (C) in Cntrl, CaOx, and CaOx + l-Erg mice. Values normalized to kidney total protein content and expressed relative to controls. Renal expression of Nfe2l2(D) and GPx1(E) genes assessed by qPCR. Gene expression normalized to Gapdh and expressed as fold change relative to the control group. (F) Percentage of body weight loss during 5 days of glyoxylate injections, calculated as [(initial weight − final weight)/(initial weight)] × 100. (G)Ex vivo mitochondrial respiration in saponin-permeabilized kidney biopsies. N-Leak respiration after adding pyruvate and malate (PM); N-OXPHOS capacity after adding ADP and glutamate (PMG); NS-OXPHOS capacity after succinate (PMGS); NS-ETS capacity after FCCP; S-ETS capacity after rotenone. Oxygen flux normalized by tissue mass (mg). N = 6 per group. (H) Correlation between maximum oxygen flux and urinary S-Met levels in CaOx and CaOx + l-Erg groups. Each dot represents an individual mouse. *p < 0.05, **p < 0.01, ***p < 0.001. N, NADH-linked pathway; NS, NADH- and succinate-linked pathways; S, succinate-linked pathway; OXPHOS, oxidative phosphorylation; ETS, electron transport system; FCCP, carbonyl cyanide 4-(trifluoromethoxy) phenylhydrazone.
Mitochondrial respiration is a major source of ROS and is closely linked to glutathione-dependent antioxidant defenses. Next, we evaluated whether l-Erg restored mitochondrial respiratory function using high-resolution respirometry, given its known role in oxidative stress pathways. CaOx mice exhibited significantly lower mitochondrial respiration capacity at all stages, which was consistently restored by l-Erg treatment, although not to control levels (Fig. 7G). Furthermore, maximum oxygen flux per mass correlated exponentially with urinary S-Met levels in the CaOx and CaOx + l-Erg groups (Fig. 7H), suggesting that increased l-Erg metabolism is associated with greater improvement in mitochondrial function. Both males and females showed similar response patterns to CaOx induction and l-Erg treatment across all evaluated outcomes, as detailed in Supplementary Figure S4. Collectively, these results indicate that l-Erg alleviates CaOx-induced renal damage by restoring redox balance and mitochondrial function.
Discussion
This study builds on previous evidence of l-Erg’s antioxidant effect in cystinuria (Mayayo-Vallverdú et al., 2023) and expands its therapeutic scope. The central finding of this study is that l-Erg exerts a dual antioxidant and mitochondrial protective effect in two mechanistically distinct forms of kidney stone disease. First, we demonstrate that l-Erg not only delays stone onset but also mitigates the progression of already-formed cystine stones. This is clinically relevant because most patients present with established stones, and current treatments are limited by toxicity. To our knowledge, this is the first study to propose a dual treatment combining an antioxidant with a cystine-chelating compound (DP) to target cystine stones. The combined treatment not only further decreased urinary cystine precipitation compared with DP treatment alone but also ameliorated markers of renal damage, such as glomerulonephritis, a known side effect of DP (Font-Llitjós et al., 2007). Given the lifelong and recurrent nature of cystinuria, preserving renal function is a primary concern to prevent CKD and renal failure (Sadiq and Cil, 2022). Although cystine lithiasis arises from a primary genetic defect in cystine transport, several studies have demonstrated that cystinuric kidneys also exhibit oxidative stress, mitochondrial dysfunction, and impaired antioxidant defenses (Mayayo-Vallverdú et al., 2023). These shared downstream alterations, which are also prominent in CaOx lithiasis, support the concept that targeting redox imbalance and mitochondrial injury represents a convergent therapeutic strategy across different stone diseases.
Our findings reveal a novel mechanistic dimension of l-Erg, linking redox signaling and mitochondrial protection to renal repair. Transcriptomic analysis revealed a pronounced downregulation of immune-related pathways together with an upregulation of cell cycle genes, suggesting attenuation of inflammation and promotion of tubular repair. These findings are consistent with previous reports describing the anti-inflammatory and cytoprotective properties of l-Erg in other models of kidney injury and liver disease (Lv et al., 2024). In addition, the decrease in Ngal expression and tubular necrosis observed after l-Erg treatment corroborates the attenuation of tubular injury, as Ngal is a sensitive marker of tubular damage (Han et al., 2012). Proteomic studies in patients with cystinuria have identified urinary signatures enriched in inflammatory and oxidative stress proteins (Bourderioux et al., 2015; Kovacevic et al., 2019, 2015), supporting the relevance of these pathways in disease progression. Our data suggest that l-Erg may counteract these processes, contributing to renal homeostasis.
To determine whether the protective effects of l-Erg extended beyond cystinuria, we evaluated its impact in an acute CaOx model. The extension of l-Erg benefits to CaOx lithiasis is particularly relevant, as this is the most common stone type worldwide. In this model, l-Erg reduced crystal deposition, alleviated tubular injury, maintained GSH levels, and improved mitochondrial respiration, reinforcing the concept that oxidative and mitochondrial defects are shared mechanisms in different forms of nephrolithiasis. Beyond l-Erg’s well-described antioxidant effects, emerging evidence indicates that it directly influences mitochondrial function. Recent studies have shown that l-Erg accumulates in mitochondria (Fong et al., 2024), where it preserves structural integrity (Tng et al., 2025) and activates key enzymes involved in energy metabolism. For instance, l-Erg binds to and activates 3-mercaptopyruvate sulfur transferase, enhancing mitochondrial respiration (Sprenger et al., 2025), and can be metabolized by cystathionine gamma-lyase to stimulate increasing NAD+ levels (Petrovic et al., 2025). These mechanisms may explain the improvement in mitochondrial function observed in our study and highlight a novel dimension of l-Erg biology beyond ROS scavenging.
Our findings also underscore the importance of redox balance in stone pathophysiology. CaOx crystal induction triggered a compensatory increase in both reduced (GSH) and oxidized (GSSG) glutathione forms, reflecting oxidative stress. l-Erg treatment normalized these levels, suggesting attenuation of oxidative damage and dampening of compensatory glutathione synthesis. This effect, together with improved mitochondrial respiration, positions l-Erg as an antioxidant capable of restoring cellular homeostasis under lithogenic conditions. Taken together, these results integrate both transcriptional and functional evidence, supporting a model in which l-Erg confers renoprotection through shared downstream pathways.
From a translational perspective, l-Erg possesses several favorable properties that support its potential as a therapeutic option for nephrolithiasis. It is actively transported by OCTN1, enabling its accumulation in the kidney (Gründemann et al., 2022), and has a favorable safety profile, classified as “Generally Recognized As Safe” by the Food and Drug Administration and approved by the European Food Safety Authority for use in vulnerable populations (Turck et al., 2017). These features, combined with its long half-life and pleiotropic effects, make l-Erg an attractive candidate for clinical trials. These preclinical results strongly support early-phase clinical trials to evaluate l-Erg as a therapeutic option for nephrolithiasis.
However, some limitations should be acknowledged. First, our study was conducted in mouse models, and extrapolation to humans requires caution. Second, although l-Erg improved mitochondrial function and reduced inflammation, the precise molecular targets remain to be fully elucidated. Finally, the optimal dose and treatment schedule for human use need to be established, considering potential variability in OCTN1 transporter activity (Taubert et al., 2005; Toh et al., 2013).
In conclusion, our data support the hypothesis that targeting oxidative stress and mitochondrial dysfunction represents a common therapeutic strategy for different types of kidney stones. Our work identifies a conserved therapeutic mechanism centered on redox balance and mitochondrial protection, positioning l-Erg as a promising candidate for different forms of nephrolithiasis, either as a standalone treatment or as an adjunct to current therapies, to improve renal outcomes and reduce stone recurrence. Future clinical studies should validate these benefits and establish optimal dosing strategies for personalized nephrolithiasis management.
Materials and Methods
Electronic laboratory notebook was not used
Animal models and experimental design
Experiments were performed in accordance with the Animal Experimentation Ethics Committee of IDIBELL (AAALAC-accredited facility, protocol B9900010). Animal experiments complied with ARRIVE guidelines. Mice were housed under specific pathogen-free conditions in ventilated racks with controlled temperature, humidity, and a 12-h light/dark cycle, with ad libitum access to water and VRF1 chow (Special Diets Services, UK). All analyses were conducted blinded to treatment groups.
To assess the in vivo effects of l-Erg and DP on cystine stone formation, we used homozygous Slc7a9−/− (Feliubadaló et al., 2003) and Slc3a1D140G (Peters et al., 2003) mice on a C57Bl/6J background. At 3 months of age, mice with radiographically confirmed stones were randomized into four experimental groups (N = 10, 5 males and 5 females): control, l-Erg (16 mg/kg/day) (Mayayo-Vallverdú et al., 2023), DP (450 mg/kg/day) (Font-Llitjós et al., 2007), and combined l-Erg and DP. Treatments were administered via drinking water for 5 weeks, with weekly dose adjustments based on body weight and water intake. l-Erg dose was selected based on previous efficacy and safety data (Mayayo-Vallverdú et al., 2023); DP dose was chosen according to established murine protocols (Font-Llitjós et al., 2007). l-Erg was provided by Tetrahedron SAS; DP was obtained from Sigma-Aldrich (Burlington, MA, USA). An additional cohort (N = 5, males) was used to evaluate treatment persistence and the effect of S-methyl-l-ergothioneine (S-Met) after discontinuation.
For CaOx lithiasis, 2-month-old WT C57Bl/6J mice were assigned to three groups (N = 10, 5 males and 5 females): control, CaOx, and CaOx + l-Erg. CaOx crystals were induced by daily intraperitoneal injection of glyoxylate (100 mg/kg/day; Sigma-Aldrich, Burlington, MA, USA) for 5 consecutive days (Oh et al., 2011). In the CaOx + l-Erg group, l-Erg (16 mg/kg/day) was supplied via drinking water starting 1 week before glyoxylate injections and continued until sacrifice.
Imaging and sample collection
Cystine stone progression was monitored by in vivo X-ray imaging (IVIS Lumina XR Series III, Caliper LifeScience) as previously described (de Heredia et al., 2021) at baseline, pre-treatment, and post-treatment. X-ray images were analyzed using Living Image® software, which quantifies total density counts for each region of interest. In all images, a calibration curve composed of stones with known weights was placed next to the mice. Regions corresponding to both the calibration curve and renal calculi were manually delineated. Stone weight was extrapolated from the calibration curve. Stone growth rate was subsequently calculated from two consecutive images acquired 1 month apart using the equation Δweight/Δtime.
Urine was collected in metabolic cages over 24 h. At the end of experiments, mice were euthanized by intracardiac puncture under anesthesia, followed by cervical dislocation. Kidneys and stones were collected; half of one kidney was fixed in 4% paraformaldehyde for histology, the other half for mitochondrial analysis, and the remainder was snap-frozen for molecular analysis.
Cystine precipitation assay
A cystine precipitation assay was performed as detailed (Zee et al., 2017). Urine pools from each treatment were incubated with a supersaturated l-cystine solution (4 mM) at 4°C for 96 h. Samples were centrifuged (4000 rpm, 20 min, RT), and pellets were resuspended in ultrapure water. Cystine concentrations in urine and pellets were quantified by Ultra-Performance Liquid Chromatography (UPLC)-Mass spectrometry (MS)/MS using internal standards (Casado et al., 2018).
Histology and apoptosis
Kidneys were paraffin-embedded and sectioned (5 µm). H&E; Dako, Santa Clara, CA, USA), Von Kossa (Abcam, Cambridge, MA, USA), and TUNEL (Roche Life Science, Basel, CHE) staining were performed following manufacturer protocols. Images were acquired at 10× and 20× magnification using a Nikon Eclipse 80i microscope (Leica Microsystems, Wetzlar, DE). Histological scoring was performed blinded to treatment (0 = no injury; 1 = <25%; 2 = <50%; 3 = >50%).
RNA sequencing and qPCR
Total RNA was extracted from frozen kidney tissue using TRIzol Reagent (Ambion®, Thermo Fisher Scientific, MA, USA) and a Polytron PT-1200 homogenizer (Kinematica, Switzerland) and purified with Direct-zol™ RNA MiniPrep Plus Kit (Zymo Research, CA, USA) following manufacturer instructions. RNA quantity and purity were assessed using a Nanodrop One spectrophotometer, and integrity was verified with RNA ScreenTape assay on the Tapestation 4200 platform (Agilent Technologies, Santa Clara, USA). Only samples with RNA Integrity Number (RIN) ≥7 were processed for sequencing.
Libraries were prepared using the Illumina® Stranded Total RNA Prep with Ribo-Zero Plus kit and dual-indexed adapters (UD Indexes Set A, Illumina). Ten PCR amplification cycles were applied to all libraries. Sequencing was performed on a NovaSeq6000 S4 flow cell (Illumina) in paired-end mode (150 nt reads), generating ≥30 million paired-end reads per sample. Raw reads were quality-checked using FastQC and trimmed with Trimmomatic before alignment to the mouse reference genome (GRCm39) using STAR aligner (v2.7). Gene counts were obtained with featureCounts.
Differential expression analysis was conducted using DESeq2 in R (v3.6.0). Genes with p value <0.05 and |log2 fold change| >1 were considered differentially expressed. Functional enrichment analysis of DEGs was performed using DAVID (v6.8) (Huang et al., 2009; Sherman et al., 2022) and Gene Ontology (GO) categories. Selected genes were validated by quantitative PCR (qPCR) using SYBR® Green Master Mix (NZYTECH, Lisbon, Portugal) on a LightCycler 480 Real-Time PCR System (Roche, Basel, Switzerland). Primers were designed with Primer-BLAST (see Supplementary Table S2 for primers used) and verified for efficiency (90%–110%). Relative expression was calculated using the ΔΔCt method, normalizing to Gapdh as the reference gene.
l-Erg and S-Met determination
Urinary concentrations of l-Erg and S-Met were quantified by LC-MS/MS as described (Mayayo-Vallverdú et al., 2023), using an Agilent 1290 Infinity II UHPLC system (Agilent Technologies, Santa Clara, USA) coupled to a Sciex 6500 QTRAP mass spectrometer equipped with an Ion Drive Turbo V ion source (Sciex, Framingham, MA, USA). Isotopically labeled internal standards were used for absolute quantification. Data were normalized to urinary creatinine concentration.
Glutathione analysis
Reduced (GSH) and oxidized (GSSG) glutathione levels were measured in kidney homogenates. Pulverized tissue was homogenized in PBS-N-ethylmaleimide buffer (11.9 mM), and protein precipitation was done with perchloric acid (4% final concentration). Samples were centrifuged at 15,000 g for 15 min at 4°C, and supernatants were analyzed by UPLC-MS/MS as described (Escobar et al., 2016). Results were normalized to total protein content determined by the Bicinchoninic Acid assay (BCA) Protein Assay Kit (Thermo Scientific, MA, USA).
Mitochondrial respirometry
High-resolution respirometry was performed on freshly isolated kidney biopsies permeabilized with saponin (0.05 mg/mL) using an Oxygraph-2k (Oroboros® Instrument GmbH Corp) at 37°C as described (Mayayo-Vallverdú et al., 2023). The SUIT protocol included sequential addition of substrates and inhibitors: leak state was analyzed by adding malate (2 mM) and pyruvate (10 mM); complex I-coupled respiration (oxidative phosphorylation) by adding ADP (5 mM); maximal NADH-linked pathway by adding glutamate (10 mM); and simultaneous NADH- and succinate-linked pathways by adding succinate (10 mM). Titration of carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone was performed to explore maximal uncoupled respiration and electron transfer system capacity. Finally, by adding rotenone (0.5 μM) and antimycin A (2.5 μM), complex I and complex II were inhibited, respectively, to assess residual oxygen consumption and to subtract this last measurement from the other respiratory states. Instrument calibration and background correction were performed daily.
Statistical analysis
Normality of data distribution was assessed using the Shapiro–Wilk test prior to statistical analysis. Bar graphs represent mean ± SEM, whereas boxplots display the median with interquartile range. For two-group comparisons, Student’s t-test was applied; for comparisons involving more than two groups, one-way ANOVA followed by Tukey’s post hoc test was used. Histological injury scores (0 = no injury; 1 = <25%; 2 = <50%; 3 = >50%) were treated as ordinal variables; thus, the Kruskal–Wallis test followed by Dunn’s post hoc with Holm correction for multiple testing was applied. Correlations were evaluated using Pearson’s coefficient. Significance was set at p ≤ 0.05. All analyses and visualizations were performed in R (v3.6.0) using packages ggplot2, dplyr, ggpubr, corrplot, and DESeq2.
Authors’ Contributions
Conceptualization: C.M.-V., R.E., and V.N.; Methodology: C.M.-V., R.E., and V.N.; Formal analysis: C.M.-V., M.V.-P., and E.P.; Investigation: C.M.-V., M.V.-P., E.P., S.C.-T., A.O., R.A., F.V.P., M.A.A., and P.M.G.-R.; Data curation: C.M.-V. and M.V.-P.; Writing—original draft: C.M.-V., M.V.-P., and R.E.; Writing—review and editing: V.N., S.L., and M.P.; Funding acquisition: V.N., M.P., R.E., F.V.P., and P.M.G.-R.; Resources: V.N., M.P., and R.E.; Supervision: V.N., S.L., M.P., and R.E.; Project administration: V.N. and R.E. All authors have read and approved the final version of the article.
Footnotes
Acknowledgments
In memory of Professor Virginia Nunes. We acknowledge the CCITUB and Dr. O. Jauregui for her guidance in LC-MS/MS analysis; the IRB Barcelona Functional Genomics Core Facility; and Lola Mulero, Jose Antonio Llamas, and Mireia Torres from the histology unit of IDIBELL.
Availability of Data and Materials
The RNA-seq data presented in the study are deposited in the GEO repository, accession number GSE311516. The datasets used and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Author Disclosure Statement
The authors declare the following financial interests/personal relationships that may be considered potential competing interests: C.M.-V., R.E., and V.N. received support funded by AVEROA SAS.
Funding Information
This work was funded by the Instituto de Salud Carlos III through project PI20/00200 to V.N. (cofunded by the European Regional Development Fund [ERDF], a way to build Europe), and by La Marató de TV3 through projects 202025-30 to V.N. and C.M.-V., 202025-31 to M.P., and 202025-32 to F.V.P. The MICINN projects PID2022-138537OB-I00 to P.M.G-R., PID2021-126246NB-I00, and PID2024-159674NB-100 cofunded by MICIU/AEI/10.13039/501100011033 and FEDER/UE to R.E. and the Generalitat de Catalunya (grant numbers 2021SGR052). S.L. is sponsored by the Researchers Consolidation Program (SNS/Generalitat de Catalunya) (CES09/020). In addition, we thank the CERCA Program/Generalitat de Catalunya for their institutional support.
Supplemental Material
Supplemental Material
Supplemental Material
Abbreviations
References
1.
AbufarajM, XuT, CaoC, et al.Prevalence and trends in kidney stone among adults in the USA: Analyses of national health and nutrition examination survey 2007–2018 Data. Eur Urol Focus, 2021; 7(6):1468–1475; doi: 10.1016/J.EUF.2020.08.011
2.
AkanmuD, CecchiniR, AruomaOI, et al.The antioxidant action of ergothioneine. Arch Biochem Biophys, 1991; 288(1):10–16; doi: 10.1016/0003-9861(91)90158-F
3.
AlelignT, PetrosB. Kidney stone disease: An update on current concepts. Adv Urol, 2018; 2018:3068365; doi: 10.1155/2018/3068365
4.
AruomaOI, WhitemanM, EnglandTG, et al.Antioxidant action of ergothioneine: Assessment of Its ability to scavenge peroxynitrite. Biochem Biophys Res Commun, 1997; 231(2):389–391; doi: 10.1006/BBRC.1997.6109
5.
BourderiouxM, Nguyen-KhoaT, ChhuonC, et al.A new workflow for proteomic analysis of urinary exosomes and assessment in cystinuria patients. J Proteome Res, 2015; 14(1):567–577; doi: 10.1021/PR501003Q
6.
CalongeMJ, GaspariniP, ChillarónJ, et al.Cystinuria caused by mutations in RBAT, a gene involved in the transport of cystine. Nat Genet, 1994; 6(4):420–425; doi: 10.1038/NG0494-420
7.
CaoLC, HoneymanTW, CooneyR, et al.Mitochondrial dysfunction is a primary event in renal cell oxalate toxicity. Kidney Int, 2004; 66(5):1890–1900; doi: 10.1111/J.1523-1755.2004.00963.X
8.
CasadoM, SierraC, BatlloriM, et al.A targeted metabolomic procedure for amino acid analysis in different biological specimens by ultra-high-performance liquid chromatography-tandem mass spectrometry. Metabolomics, 2018; 14(6):76; doi: 10.1007/S11306-018-1374-4
9.
ChaiyaritS, ThongboonkerdV. Mitochondrial dysfunction and kidney stone disease. Front Physiol, 2020; 11:566506; doi: 10.3389/FPHYS.2020.566506
10.
DareA, ChannaML, NadarA. L-Ergothioneine and Its combination with metformin attenuates renal dysfunction in type-2 diabetic rat model by activating Nrf2 antioxidant pathway. Biomed Pharmacother, 2021; 141:111921; doi: 10.1016/J.BIOPHA.2021.111921
11.
DeianaM, RosaA, CasuV, et al.L-Ergothioneine modulates oxidative damage in the kidney and liver of rats in vivo: Studies upon the profile of polyunsaturated fatty acids. Clin Nutr, 2004; 23(2):183–193; doi: 10.1016/S0261-5614(03)00108-0
12.
EscobarJ, Sánchez-IllanaÁ, KuligowskiJ, et al.Development of a reliable method based on ultra-performance liquid chromatography coupled to tandem mass spectrometry to measure thiol-associated oxidative stress in whole blood samples. J Pharm Biomed Anal, 2016; 123:104–112; doi: 10.1016/J.JPBA.2016.02.007
13.
FeliubadalóL, ArbonésML, MañasS, et al.Slc7a9-Deficient mice develop cystinuria non-I and cystine urolithiasis. Hum Mol Genet, 2003; 12(17):2097–2108; doi: 10.1093/HMG/DDG228
14.
FeliubadalóL, FontM, PurroyJ, et al.; International Cystinuria Consortium. Non-Type I cystinuria caused by mutations in SLC7A9, encoding a subunit (Bo,+AT) of RBAT. Nat Genet, 1999; 23(1):52–57; doi: 10.1038/12652
15.
FongZW, TangRMY, CheahIKM, et al.Ergothioneine and mitochondria: An important protective mechanism? Biochem Biophys Res Commun, 2024; 726:150269; doi: 10.1016/J.BBRC.2024.150269
16.
Font-LlitjósM, FeliubadalóL, EspinoM, et al.Slc7a9 knockout mouse is a good cystinuria model for antilithiasic pharmacological studies. Am J Physiol Renal Physiol, 2007; 293(3):F732–F740; doi: 10.1152/AJPRENAL.00121.2007
17.
GründemannD, HartmannL, FlögelS. The ergothioneine transporter (ETT): Substrates and locations, an inventory. FEBS Lett, 2022; 596(10):1252–1269; doi: 10.1002/1873-3468.14269
18.
HabibGS, SalibaW, NashashibiM, et al.Penicillamine and nephrotic syndrome. Eur J Intern Med, 2006; 17(5):343–348; doi: 10.1016/J.EJIM.2006.03.001
19.
HalperinEC, ThierSO, RosenbergLE. The use of D-Penicillamine in cystinuria: Efficacy and untoward reactions. Yale J Biol Med, 1981; 54(6):439–446.
20.
HanM, LiY, LiuM, et al.Renal neutrophil gelatinase associated lipocalin expression in lipopolysaccharide-induced acute kidney injury in the rat. BMC Nephrol, 2012; 13(1):25; doi: 10.1186/1471-2369-13-25
21.
de HerediaML, MuñozL, CarruC, et al.S-Methyl-L-Ergothioneine to L-Ergothioneine Ratio in Urine Is a Marker of Cystine Lithiasis in a Cystinuria Mouse Model. Antioxidants (Basel, Switzerland), 2021; 10(9); doi: 10.3390/ANTIOX10091424
22.
HiroseM, YasuiT, OkadaA, et al.Renal tubular epithelial cell injury and oxidative stress induce calcium oxalate crystal formation in mouse kidney. Int J Urol, 2010; 17(1):83–92; doi: 10.1111/J.1442-2042.2009.02410.X
23.
HuangDW, ShermanBT, LempickiRA. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat Protoc, 2009; 4(1):44–57; doi: 10.1038/NPROT.2008.211
24.
KönigsbergerE, WangZ, KönigsbergerL-C. Solubility of L-Cystine in NaCl and artificial urine solutions. Monatshefte Für Chemie/Chem Mon, 2000; 131(1):0039–0045; doi: 10.1007/s007060050004
25.
KovacevicL, CarusoJA, LuH, et al.Urine proteomic profiling in patients with nephrolithiasis and cystinuria. Int Urol Nephrol, 2019; 51(4):593–599; doi: 10.1007/S11255-018-2044-1
26.
KovacevicL, LuH, GoldfarbDS, et al.Urine proteomic analysis in cystinuric children with renal stones. J Pediatr Urol, 2015; 11(4):217.e1-217–e6; doi: 10.1016/J.JPUROL.2015.04.020
27.
LasotaA, WasilewskaA, Rybi-SzumińskaA. Current status of protein biomarkers in urolithiasis—a review of the recent literature. J Clin Med, 2023; 12(22):7135; doi: 10.3390/JCM12227135
28.
LiuJ, HuangJ, GongB, et al.Polydatin protects against calcium oxalate crystal-induced renal injury through the cytoplasmic/mitochondrial reactive oxygen species-NLRP3 inflammasome pathway. Biomed Pharmacother, 2023; 167:115621; doi: 10.1016/J.BIOPHA.2023.115621
29.
LvX, NieC, ShiY, et al.Ergothioneine ameliorates metabolic dysfunction-associated steatotic liver disease (MASLD) by enhancing autophagy, inhibiting oxidative damage and inflammation. Lipids Health Dis, 2024; 23(1):395; doi: 10.1186/S12944-024-02382-9
30.
MarhoumeFZ, AboufatimaR, ZaidY, et al.Antioxidant and polyphenol-rich ethanolic extract of rubia tinctorum L. prevents urolithiasis in an ethylene glycol experimental model in rats. Molecules, 2021; 26(4):1005; doi: 10.3390/MOLECULES26041005
31.
Mayayo-VallverdúC, López de HerediaM, PratE, et al.The antioxidant l-ergothioneine prevents cystine lithiasis in the Slc7a9-/- Mouse model of cystinuria. Redox Biol, 2023; 64:102801; doi: 10.1016/J.REDOX.2023.102801
32.
ModersitzkiF, PizziL, GrassoM, et al.Health-Related quality of life (HRQoL) in cystine compared with non-cystine stone formers. Urolithiasis, 2014; 42(1):53–60; doi: 10.1007/S00240-013-0621-4
33.
NettuwakulC, SawasdeeN, PraditsapO, et al.A novel loss-of-function mutation of PBK associated with human kidney stone disease. Sci Rep, 2020; 10(1):10282; doi: 10.1038/S41598-020-66936-4
34.
NiimiK, YasuiT, OkadaA, et al.Novel effect of the inhibitor of mitochondrial cyclophilin D activation, N-Methyl-4-Isoleucine cyclosporin, on renal calcium crystallization. Int J Urol, 2014; 21(7):707–713; doi: 10.1111/IJU.12425
35.
OhSY, KwonJK, LeeSY, et al.A comparative study of experimental rat models of renal calcium oxalate stone formation. J Endourol, 2011; 25(6):1057–1061; doi: 10.1089/END.2010.0386
36.
PakCYC, FullerC, SakhaeeK, et al.Management of cystine nephrolithiasis with alpha-mercaptopropionylglycine. J Urol, 1986; 136(5):1003–1008; doi: 10.1016/S0022-5347(17)45188-3
37.
PaulBD, SnyderSH. The unusual amino acid l-ergothioneine is a physiologic cytoprotectant. Cell Death Differ, 2010; 17(7):1134–1140; doi: 10.1038/CDD.2009.163
38.
PengJ, ChenJ, WuM, et al.Proteomic analysis reveals the potential mechanism of ergothioneine in preventing acute kidney injury to chronic kidney disease transition. Arch Biochem Biophys, 2025; 772:110534; doi: 10.1016/J.ABB.2025.110534
39.
PetersT, ThaeteC, WolfS, et al.A mouse model for cystinuria type I. Hum Mol Genet, 2003; 12(17):2109–2120; doi: 10.1093/HMG/DDG189
40.
PetrovicD, SladeL, PaikopoulosY, et al.Ergothioneine improves healthspan of aged animals by enhancing CGPDH activity through CSE-Dependent persulfidation. Cell Metab, 2025; 37(2):542–556.e14; doi: 10.1016/J.CMET.2024.12.008
41.
PietersTT, FalkeLL, NguyenTQ, et al.Histological characteristics of acute tubular injury during delayed graft function predict renal function after renal transplantation. Physiol Rep, 2019; 7(5):e14000; doi: 10.14814/PHY2.14000
42.
Prot-BertoyeC, LebbahS, DaudonM, et al.; French Cystinuria Group. CKD and its risk factors among patients with cystinuria. Clin J Am Soc Nephrol, 2015; 10(5):842–851; doi: 10.2215/CJN.06680714
43.
Prot-BertoyeC, LebbahS, DaudonM, et al.; French Cystinuria Group. Adverse events associated with currently used medical treatments for cystinuria and treatment goals: Results from a Series of 442 Patients in France. BJU Int, 2019; 124(5):849–861; doi: 10.1111/BJU.14721
44.
SadiqS, CilO. Cystinuria: An overview of diagnosis and medical management. Turk Arch Pediatr, 2022; 57(4):377–384; doi: 10.5152/TurkArchPediatr.2022.22105
45.
SalamaSA, Abd-AllahGM, MohamadinAM, et al.Ergothioneine mitigates cisplatin-evoked nephrotoxicity via targeting Nrf2, NF-ΚB, and apoptotic signaling and inhibiting γ-Glutamyl Transpeptidase. Life Sci, 2021; 278:119572; doi: 10.1016/J.LFS.2021.119572
46.
ServaisA, ThomasK, Dello StrologoL, et al.; Metabolic Nephropathy Workgroup of the European Reference Network for Rare Kidney Diseases (ERKNet) and eUROGEN. Cystinuria: Clinical practice recommendation. Kidney Int, 2021; 99(1):48–58; doi: 10.1016/J.KINT.2020.06.035
47.
SharmaM, KaurT, SinglaSK. Role of mitochondria and nadph oxidase derived reactive oxygen species in hyperoxaluria induced nephrolithiasis: Therapeutic intervention with combinatorial therapy of n-acetyl cysteine and apocynin. Mitochondrion, 2016; 27:15–24; doi: 10.1016/J.MITO.2016.01.002
48.
ShermanBT, HaoM, QiuJ, et al.DAVID: A web server for functional enrichment analysis and functional annotation of gene lists (2021 Update). Nucleic Acids Res, 2022; 50(W1):W216–W221; doi: 10.1093/NAR/GKAC194
49.
SprengerH-G, MittenbühlerMJ, SunY, et al.Ergothioneine controls mitochondrial function and exercise performance via direct activation of MPST. Cell Metab, 2025; 37(4):857–869.e9; doi: 10.1016/J.CMET.2025.01.024
50.
StoffelsC, OumariM, PerrouA, et al.Ergothioneine stands out from hercynine in the reaction with singlet oxygen: Resistance to glutathione and TRIS in the generation of specific products indicates high reactivity. Free Radic Biol Med, 2017; 113:385–394; doi: 10.1016/J.FREERADBIOMED.2017.10.372
51.
StreeperNM, WertheimML, NakadaSY, et al.Cystine stone formers have impaired health-related quality of life compared with noncystine stone formers: A case-referent study piloting the Wisconsin stone quality of life questionnaire among patients with cystine stones. J Endourol, 2017; 31(S1):S48–S53; doi: 10.1089/END.2016.0564
52.
StrologoLD, PrasE, PontesilliC, et al.Comparison between SLC3A1 and SLC7A9 Cystinuria Patients and Carriers: A Need for a New Classification. J Am Soc Nephrol, 2002; 13(10):2547–2553; doi: 10.1097/01.ASN.0000029586.17680.E5
53.
TangRMY, CheahIKM, YewTSK, et al.Distribution and accumulation of dietary ergothioneine and its metabolites in mouse tissues. Sci Rep, 2018; 8(1):1601; doi: 10.1038/S41598-018-20021-Z
54.
TaubertD, GrimbergG, JungN, et al.Functional role of the 503F variant of the organic cation transporter OCTN1 in Crohn’s disease. Gut, 2005; 54(10):1505–1506; doi: 10.1136/GUT.2005.076083
55.
TavianatouAG, CaonI, FranchiM, et al.Hyaluronan: Molecular size-dependent signaling and biological functions in inflammation and cancer. Febs J, 2019; 286(15):2883–2908; doi: 10.1111/FEBS.14777
56.
TngTJW, LeowDMK, GohG, et al.Ergothioneine treatment ameliorates the pathological phenotypes of Parkinson’s disease models. J Neurochem, 2025; 169(7):e70168; doi: 10.1111/JNC.70168
57.
TohDSL, CheungFSG, MurrayM, et al.Functional analysis of novel variants in the organic cation/ergothioneine transporter 1 identified in Singapore populations. Mol Pharm, 2013; 10(7):2509–2516; doi: 10.1021/MP400193R
58.
TurckD, BressonJL, BurlingameB, et al.; EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Statement on the safety of synthetic l-ergothioneine as a novel food - supplementary dietary exposure and safety assessment for infants and young children, pregnant and breastfeeding women. EFS2, 2017; 15(11); doi: 10.2903/J.EFSA.2017.5060
59.
WeiQ, ShenY, TianY, et al.Diagnostic potential of CDK1 and STAT1 in acute kidney injury associated with gastrointestinal cancers: A bioinformatics-based study. Front Mol Biosci, 2025; 12:1522246; doi: 10.3389/FMOLB.2025.1522246
60.
WuY, XunY, ZhangJ, et al.Resveratrol attenuates oxalate-induced renal oxidative injury and calcium oxalate crystal deposition by regulating TFEB-Induced autophagy pathway. Front Cell Dev Biol, 2021; 9:638759; doi: 10.3389/FCELL.2021.638759
61.
XuY, LiG, GeD, et al.Mitochondrial dysfunction in kidney stones and relief of kidney stones after reducing MtROS. Urolithiasis, 2024; 52(1):117; doi: 10.1007/S00240-024-01614-Y
62.
XuY, YouJ, YaoJ, et al.Klotho alleviates oxidative stress and mitochondrial dysfunction through the Nrf2/HO-1 pathway, thereby reducing renal senescence induced by calcium oxalate crystals. Urolithiasis, 2025; 53(1):61; doi: 10.1007/S00240-025-01734-Z
63.
YoussefRF, MartinJW, SakhaeeK, et al.Rising occurrence of hypocitraturia and hyperoxaluria associated with increasing prevalence of stone disease in calcium kidney stone formers. Scand J Urol, 2020; 54(5):426–430; doi: 10.1080/21681805.2020.1794955
64.
ZeeT, BoseN, ZeeJ, et al.α-Lipoic acid treatment prevents cystine urolithiasis in a mouse model of cystinuria. Nat Med, 2017; 23(3):288–290; doi: 10.1038/NM.4280
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.