
Abstract
Aims:
Liver ischemia-reperfusion injury (IRI) is a sterile inflammatory process that contributes significantly to graft rejection following liver transplantation. Although SET domain bifurcated histone lysine methyltransferase 1 (SETDB1) is known to preserve genomic stability and restrain inflammation under oxidative stress, its immunoregulatory function in myeloid cells during liver IRI has not been elucidated. This study aimed to investigate the role and mechanism of SETDB1 in regulating macrophage-driven inflammatory responses in liver IRI.
Results:
Myeloid-specific SETDB1 knockout (SETDB1 cKO) mice exhibited exacerbated liver injury, increased infiltration of pro-inflammatory macrophages and neutrophils, and amplified inflammatory responses compared with SETDB1fl/fl controls. Depletion of macrophages alleviated liver damage, reduced neutrophil infiltration and hepatocyte apoptosis, and eliminated the excessive injury observed in SETDB1 cKO mice. Mechanistically, SETDB1 suppressed the expression of purinergic receptor P2X7 (P2RX7). Pharmacological inhibition of P2RX7 with oxidized adenosine triphosphate significantly attenuated liver injury and macrophage infiltration in SETDB1 cKO mice. In vitro assays confirmed that SETDB1 inhibited the P2RX7/Caspase-1/Gasdermin D (GSDMD) pathway in macrophages, thereby limiting pyroptosis and inflammation.
Innovation:
This study identifies SETDB1 as a previously unrecognized regulator of macrophage pyroptosis during liver IRI. By linking epigenetic regulation to suppression of the P2RX7/Caspase-1/GSDMD pathway, our findings provide novel mechanistic insight into how SETDB1 protects against sterile liver inflammation.
Conclusion:
SETDB1 plays a pivotal role in protecting the liver from IRI by restraining macrophage-mediated pyroptosis and inflammation. These findings suggest that targeting the SETDB1/P2RX7/Caspase-1/GSDMD axis may represent a promising therapeutic strategy for mitigating liver IRI and improving transplant outcomes. Antioxid. Redox Signal. 45, 133–148.
Introduction
Liver ischemia-reperfusion injury (IRI) significantly contributes to the high mortality rates observed following hepatectomy and liver transplantation, often exacerbating liver damage and failure (George et al., 2024; Peralta et al., 2013). The pathogenesis of liver IRI involves a series of inflammatory responses and oxidative stress induced by the abrupt restoration of blood flow and oxygen supply to the ischemic liver (George et al., 2024). This cascade of events is central to the progression of liver damage, highlighting the importance of elucidating the immune cell dynamics and molecular pathways involved in IRI. A deeper understanding of these mechanisms is essential for developing effective therapeutic strategies to protect the liver and improve transplantation outcomes.
In the early stages of IRI, macrophages are swiftly activated and polarized toward the pro-inflammatory M1 phenotype. These M1 macrophages release a range of pro-inflammatory cytokines in the injured area, including tumor necrosis factor-alpha (TNF-α), interleukin-1beta (IL-1β), interleukin-6 (IL-6), reactive oxygen species (ROS), nitric oxide (NO), complement proteins, and chemokines (Colletti et al., 1990; Kato et al., 2002; Wang et al., 2021). These inflammatory factors further aggravate hepatocyte damage and trigger infiltration of peripheral immune cells, thereby amplifying the inflammatory response (Dong et al., 2007). This pro-inflammatory cascade is a key driver of liver IRI and represents one of the central mechanisms underlying liver injury (Bamboat et al., 2010; Zhou et al., 2018). Given the critical role of macrophages in the initiation and propagation of the inflammatory response, targeting macrophage regulatory pathways to modulate IRI is a promising therapeutic approach.
SET domain bifurcated histone lysine methyltransferase 1 (SETDB1), a histone lysine methyltransferase, catalyzes the trimethylation of histone H3 at lysine 9 (H3K9), thereby regulating chromatin silencing, gene expression, and genome stability (Schultz et al., 2002; Strepkos et al., 2021). Recent studies have highlighted SETDB1’s important role in cellular responses to damage, inflammation, and oxidative stress (Chen et al., 2023; Južnić et al., 2021; Park et al., 2023; Wang et al., 2020; Xia et al., 2023). Through its H3K9 methylation activity, SETDB1 suppresses the expression of pro-inflammatory genes in M1 macrophages, such as TNF-α, IL-1β, and IL-6, and promotes the expression of anti-inflammatory factors such as IL-10 in M2 macrophages, thus limiting the production of inflammatory mediators and enhancing macrophage-driven tissue repair and anti-inflammatory functions (Eames et al., 2012; Hachiya et al., 2016). Additionally, SETDB1 has been implicated in the regulation of antioxidant enzymes, such as superoxide dismutase (SOD) and catalase (CAT), which mitigate elevated ROS levels and oxidative damage in macrophages (He et al., 2015). Despite these insights, the specific role of SETDB1 in macrophages during liver IRI remains insufficiently explored.
The purinergic receptor P2X7 (P2RX7), a member of the purinergic ATP-gated ion channel family, is widely expressed in almost all immune cells and plays a pivotal role in regulating inflammasome activation, cancer-associated inflammation, immune defense against pathogens, T cell differentiation, neurological diseases, and musculoskeletal diseases (Adinolfi et al., 2015; Baricordi et al., 1999; Bartlett et al., 2014; Csóka et al., 2015; Di Virgilio et al., 2017). In macrophages, stimulation by high extracellular ATP concentrations leads to prolonged activation of P2RX7, resulting in pore formation, ionic flux, and the activation of multiple downstream pathways including the NOD-like receptor family pyrin domain-containing protein 3 (NLRP3) inflammasome, culminating in IL-1β and IL-18 secretion and inflammatory cell death. Therefore, P2RX7 serves as a crucial sensor of danger-associated molecular patterns (DAMPs) and a central regulator of inflammation in various pathological contexts (Muñoz-Planillo et al., 2013; Ousingsawat et al., 2015; Qian et al., 2021; Savio et al., 2018; Walev et al., 1995). oxidized adenosine triphosphate (OATP) has long been recognized as an irreversible antagonist of P2RX7 through covalent modification of its ATP-binding site, thereby inhibiting receptor activation and P2RX7-dependent inflammatory signaling. However, subsequent studies have demonstrated that OATP also interacts with additional molecular targets beyond P2RX7, suggesting a broader mechanism of action. Despite this, OATP remains a valuable pharmacological tool to explore the functional contribution of P2RX7 signaling in inflammation and tissue injury. Collectively, these findings underscore the complex yet central role of P2RX7 receptor activity in immune regulation and its modulation by OATP (Di Virgilio, 2003).
In this study, we generated myeloid-specific SETDB1 knockout (SETDB1 cKO) mice to investigate the function of SETDB1 in macrophages during liver IRI. Liver IRI models were established in SETDB1fl/fl and SETDB1 cKO mice. Intensified liver damage and enhanced infiltration of pro-inflammatory macrophages were observed in SETDB1 cKO mice, while depletion of macrophages alleviated the exacerbated liver injury. Experimental evidence from in vitro models indicates that SETDB1 negatively regulates Casp1-p20 and GSDMD-N expression and reduces the secretion of inflammatory mediators. Mechanistically, we demonstrate that SETDB1 in macrophages negatively regulates the P2RX7/Caspase-1/GSDMD signaling pathway, thereby reducing macrophage pyroptosis and inflammatory responses, ultimately protecting the liver from IRI.
Results
Myeloid SETDB1 knockout exacerbates liver IRI
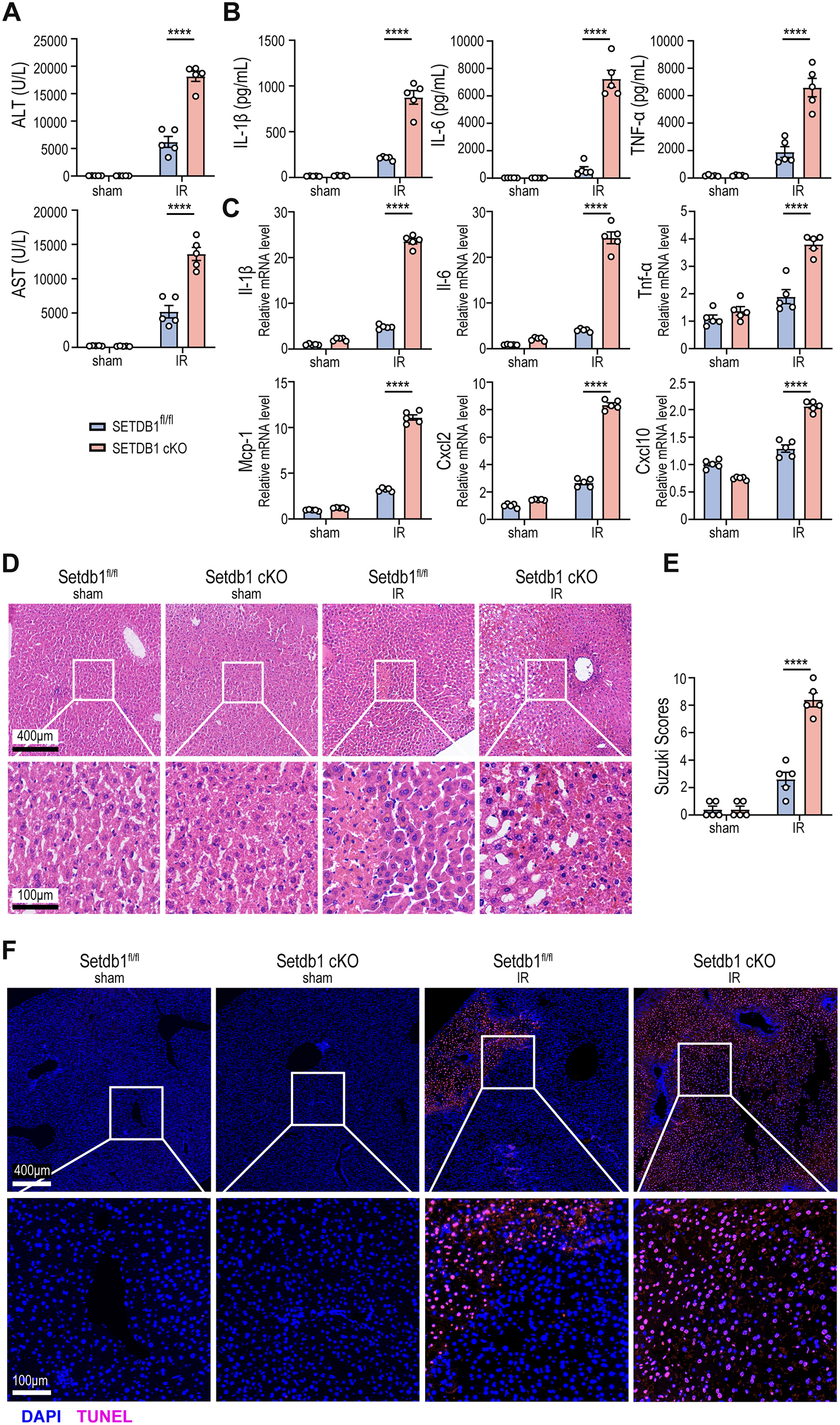
To investigate the role of myeloid SETDB1 in regulating hepatocyte injury and sterile inflammation in liver IRI, we generated myeloid-specific SETDB1 cKO mice using CRISPR/Cas9 technology. The experimental mice were subjected to 40 min of liver ischemia followed by 6 h of reperfusion, while the sham group underwent abdominal surgery without liver treatment. Compared to SETDB1fl/fl mice, SETDB1 cKO mice displayed elevated serum levels of alanine aminotransferase (ALT) and aspartate aminotransferase (AST), as well as increased serum secretion level of pro-inflammatory cytokines IL-1β, IL-6 and TNF-α (Fig. 1A and B). In liver tissue from IRI models, SETDB1 cKO mice exhibited elevated mRNA levels of inflammatory factors and chemokines, including IL-1β, IL-6, TNF-α, monocyte chemoattractant protein-1 (MCP-1), C-X-C motif chemokine ligand 2 (CXCL2), and CXCL10 (Fig. 1C). hematoxylin and eosin (H&E) staining revealed more severe liver tissue injury in SETDB1 cKO mice compared to SETDB1fl/fl mice, characterized by increased hepatocyte edema, congestion, vacuolar degeneration and extensive hepatocyte necrosis, consistent with Suzuki’s histological score for liver IRI (Fig. 1D and E). TUNEL staining also showed a higher number of apoptotic hepatocytes in SETDB1 cKO mice (Fig. 1F). These results indicate that SETDB1 cKO mice exhibit more severe hepatocyte injury and aseptic inflammation following liver IRI. Importantly, Setdb1 expression was significantly elevated in the IRI group compared with the control group, suggesting that hepatic IRI induces the upregulation of SETDB1 in liver immune cells (Supplementary Fig. S1). These findings indicate that SETDB1 may participate in the epigenetic regulation of immune responses during hepatic IRI.
Myeloid SETDB1 knockout increases myeloid immune cell infiltration in liver IRI
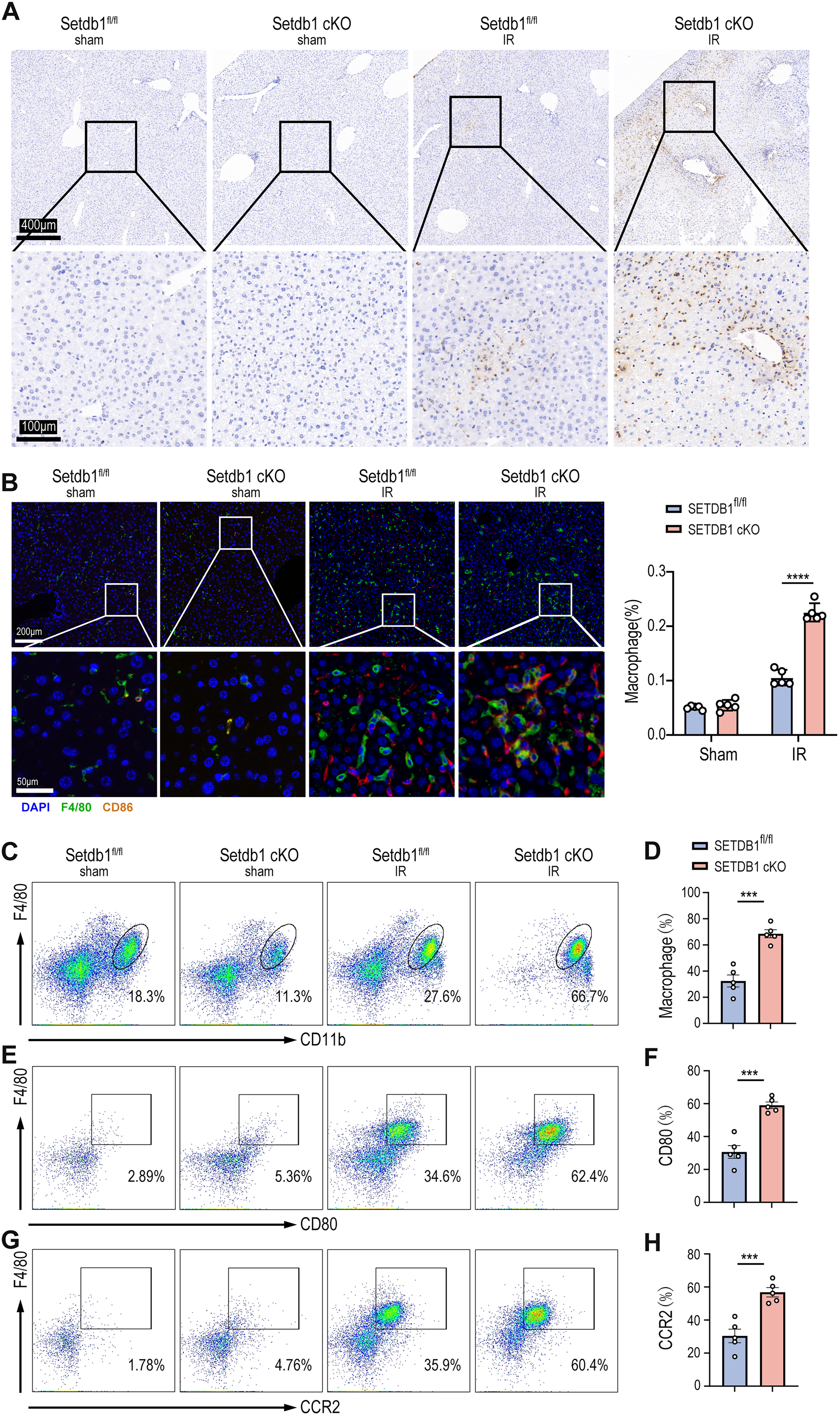
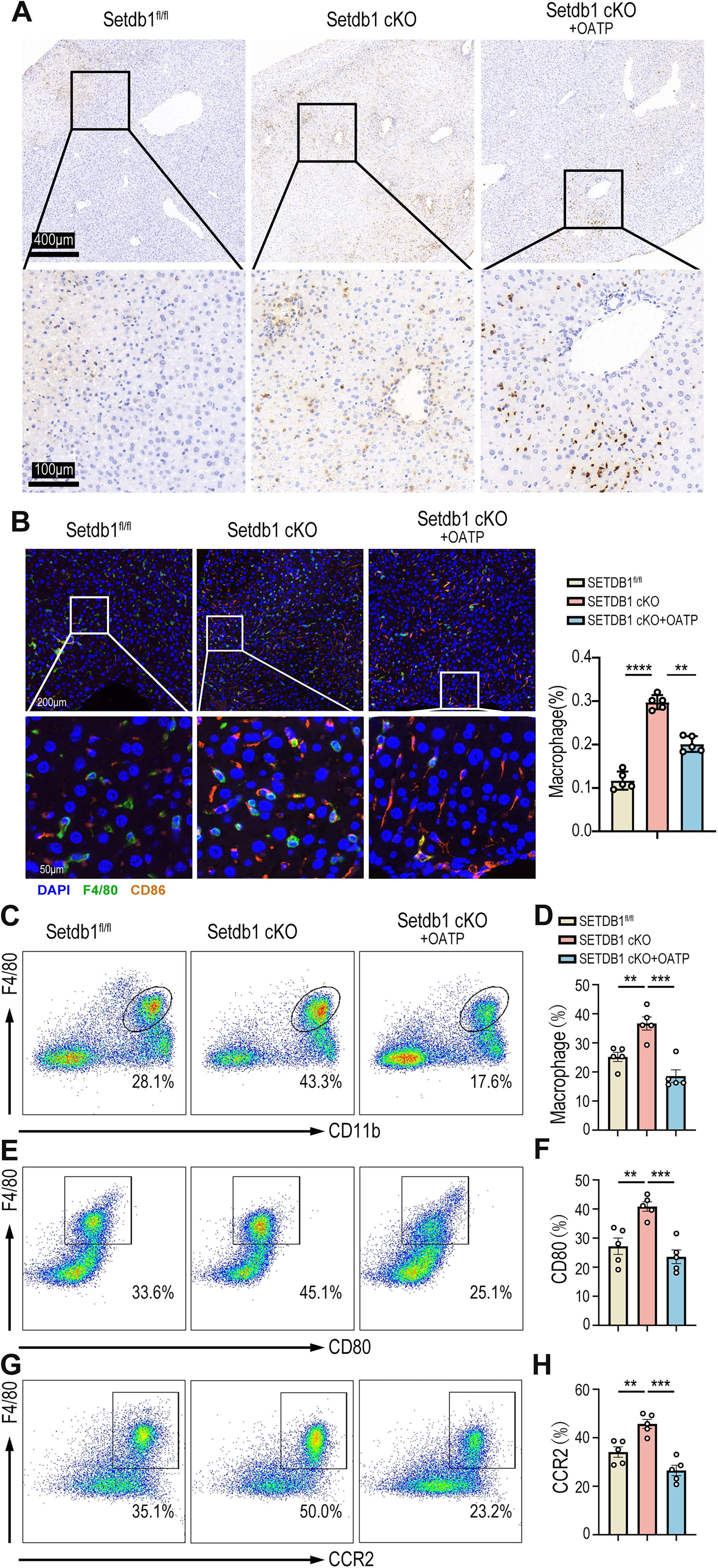
The activation of innate immune cells, particularly neutrophils and macrophages, plays an indispensable role in aseptic inflammation during liver IRI (van Golen et al., 2012). To assess the impact of myeloid SETDB1 knockout on immune cell infiltration during liver IRI, we performed pathological staining of liver tissues from both SETDB1fl/fl and SETDB1 cKO mice subjected to IRI. Neutrophil infiltration was evaluated using myeloperoxidase (MPO) staining, revealing significantly increased neutrophil infiltration in liver tissue of SETDB1 cKO mice compared to SETDB1fl/fl mice (Fig. 2A). To examine the infiltration of pro-inflammatory macrophages, we performed co-localization and quantitative analysis of macrophage surface markers F4/80 and CD86 through immunofluorescence staining, showing enhanced pro-inflammatory macrophage infiltration in SETDB1 cKO liver tissue (Fig. 2B). Flow cytometric analysis further confirmed that SETDB1 cKO mice exhibited significantly greater infiltration of peripheral mononuclear macrophages in the liver post-IRI, with increased proportions of pro-inflammatory macrophages and CC motif Chemokine Receptor 2 (CCR2+) macrophages (Fig. 2C–H). These results confirm that myeloid SETDB1 knockout exacerbates liver IRI by recruiting more pro-inflammatory neutrophils and macrophages to the liver. To investigate whether hypoxia-inducible factor (HIF) play a role in the IRI model, we performed immunohistochemical staining. The results showed no significant difference in HIF expression between the SETDB1fl/fl IRI and SETDB1 CKO IRI groups, suggesting that HIF does not contribute to SETDB1-mediated IRI (Fig. S2A).
Macrophages mediate SETDB1 cKO-Accelerated liver IRI
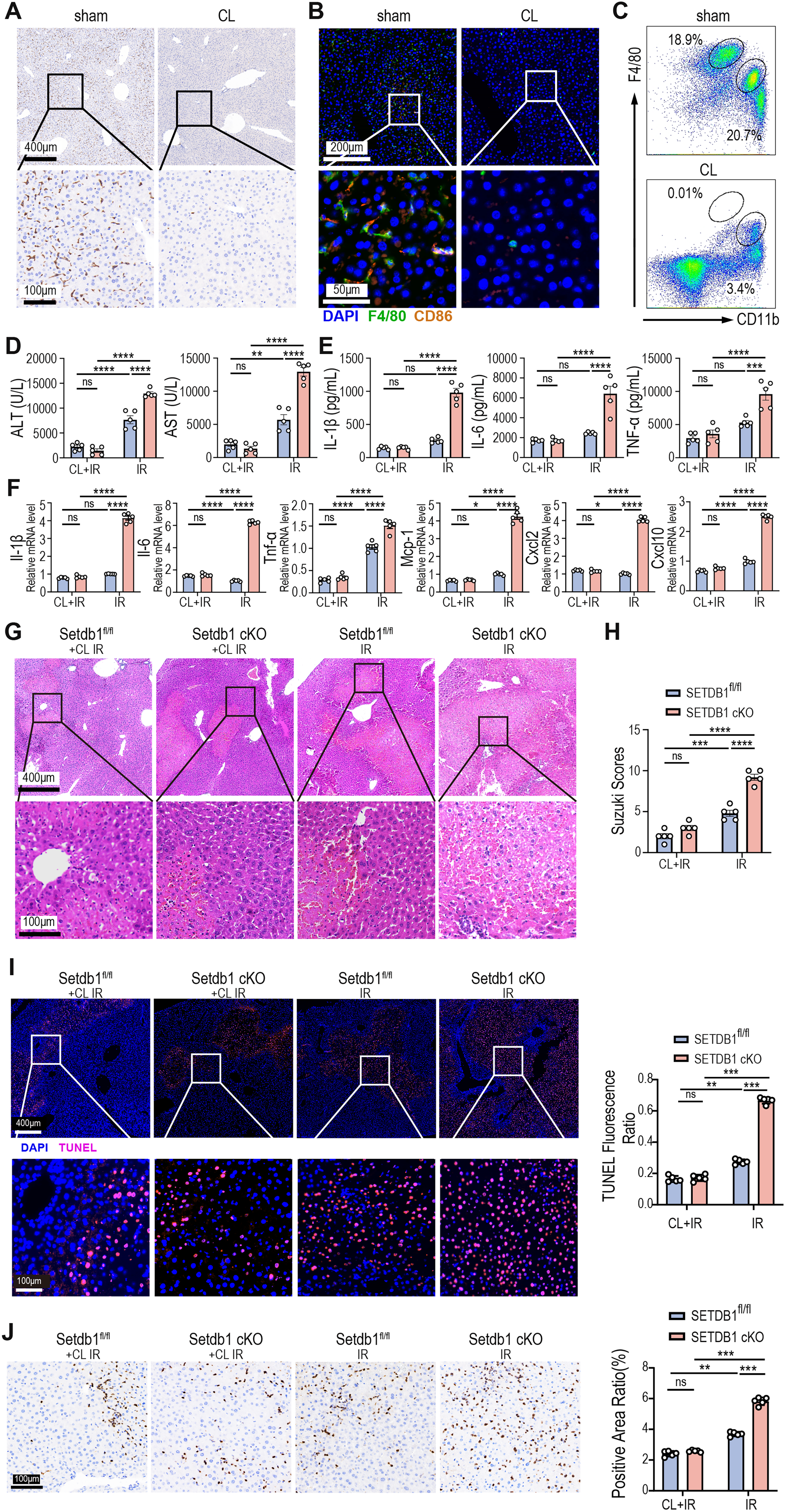
Macrophages play a pivotal role in triggering and modulating the inflammatory response during liver IRI and are considered more important than other immune cells (Roohani and Tacke, 2021; van Golen et al., 2012). To assess the role of macrophages in SETDB1 cKO-exacerbated liver IRI, we depleted macrophages using clodronate liposomes (CL) via intraperitoneal injection. After 24 h, liver tissues were analyzed for macrophage depletion efficiency via immunohistochemistry, immunofluorescence, and flow cytometry. Immunohistochemistry and immunofluorescence staining confirmed the depletion of liver macrophages (Fig. 3A and B), and flow cytometry revealed a significant reduction in the proportion of both Kupffer cells and mononuclear macrophages in liver tissue (Fig. 3C). Following macrophage depletion, SETDB1fl/fl and SETDB1 cKO mice underwent liver IRI. As expected, compared to the non-depleted macrophage groups, both SETDB1fl/fl and SETDB1 cKO mice with depleted macrophages showed decreased serum ALT and AST levels, as well as the secretion of serum inflammatory factors and inflammation-related genes and chemokines (Fig. 3D–F). Notably, depletion of macrophages alleviated the enhanced liver injury and inflammation observed in SETDB1 cKO mice relative to SETDB1fl/fl controls, including reductions in liver enzyme levels (Fig. 3D), cytokine secretion (Fig. 3E), and inflammation-related genes expression (Fig. 3F). H&E staining further indicated that macrophage depletion significantly alleviated liver injury, reduced congestion and vacuolar degeneration, and lowered Suzuki’s histological score (Fig. 3G and H). TUNEL staining suggested a decrease in apoptotic hepatocytes after macrophage depletion (Fig. 3I). Moreover, the depletion of macrophages reduced neutrophil infiltration, with no significant difference between SETDB1fl/fl and SETDB1 cKO mice (Fig. 3J). These findings confirm that macrophages mediate the exacerbation of liver IRI in SETDB1 cKO mice.
Inhibition of P2RX7 receptor mitigates SETDB1 cKO-Accelerated liver IRI
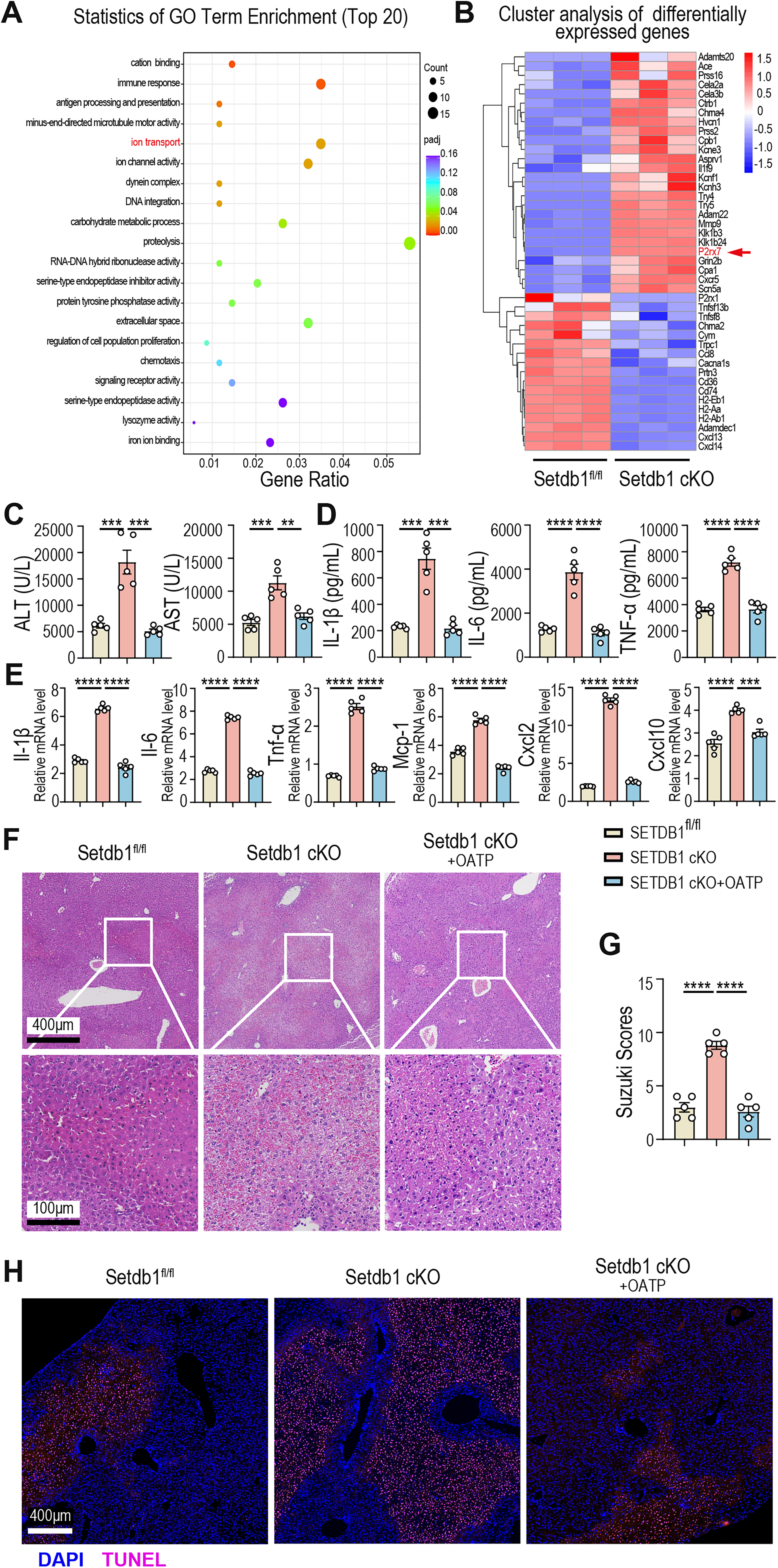
To further explore the mechanisms underlying the aggravated liver injury and pro-inflammatory macrophage infiltration observed in SETDB1 cKO mice, we performed RNA-seq analysis on liver-infiltrated macrophages after 40 min of ischemia and 6 h of reperfusion in SETDB1fl/fl and SETDB1 cKO mice. Gene Ontology analysis revealed that differential gene expression in macrophages from SETDB1fl/fl and SETDB1 cKO mice was primarily enriched in proteolysis, ion transport, and immune response pathways (Fig. 4A). Notably, P2RX7, an ATP receptor, serves as a crucial mediator linking inflammasome activation and cell pyroptosis, and accumulating evidence indicates its pivotal involvement in the inflammatory cascade associated with IRI. It was also highly expressed in macrophages from SETDB1 cKO mice (Fig. 4B). P2RX7 is a key regulator of macrophage immune responses (Junger, 2011). To investigate the role of P2RX7 in liver IRI, we administered the P2RX7 receptor inhibitor OATP to SETDB1 cKO mice prior to liver IRI. Results demonstrated that OATP treatment significantly reduced serum liver enzyme activity and decreased the secretion of pro-inflammatory cytokines IL-1β, IL-6, and TNF-α (Fig. 4C and D). Moreover, liver tissue expression of inflammatory factors and chemokines such as IL-1β, IL-6, TNF-α, MCP-1, CXCL2 and CXCL10 was lower in OATP-treated SETDB1 cKO mice (Fig. 4E). H&E and TUNEL staining indicated reduced hepatocyte necrosis, congestion, vacuolar degeneration, and apoptosis in OATP-treated SETDB1 cKO mice, in line with a decrease in Suzuki’s histological score (Fig. 4F–H). Therefore, inhibition of the P2RX7 receptor by OATP mitigated liver IRI in SETDB1 cKO mice.
Inhibition of P2RX7 receptor decreases macrophage infiltration in SETDB1 cKO-Accelerated liver IRI
Then, we further explored the impact of P2RX7 receptor inhibition on immune cell infiltration during liver IRI. MPO staining suggested that neutrophil infiltration was reduced in OATP-treated SETDB1 cKO mice (Fig. 5A). Additionally, immunofluorescence staining showed a significant reduction in pro-inflammatory macrophage infiltration (co-localized F4/80 and CD86) (Fig. 5B). Flow cytometric analysis further confirmed these findings, showing a decreased proportion of mononuclear macrophages infiltrating the liver and a reduction in the proportions of pro-inflammatory macrophages and CCR2+ macrophages after OATP treatment (Fig. 5C–H). In conclusion, inhibition of P2RX7 receptor reduces the infiltration of hepatic pro-inflammatory macrophages and the secretion of inflammatory factors in SETDB1 cKO mice, thus alleviating liver IRI.
SETDB1 protects BMDM from pyroptosis via the P2RX7/caspase-1/GSDMD pathway in vitro
P2RX7 activation triggers pyroptosis in macrophages by inducing NLRP3 inflammasome activation and caspase-1 cleavage (Bidula et al., 2019). This form of pyroptosis plays a crucial role in both acute and chronic inflammatory responses and is closely associated with various diseases (Robinson et al., 2019). To investigate whether SETDB1 is involved in the pyroptosis signaling pathway, we isolated bone marrow cells from SETDB1fl/fl and SETDB1 cKO mice and induced them into macrophages. These bone marrow-derived macrophages (BMDMs) were then stimulated with lipopolysaccharide (LPS) (500 ng/mL) for 8 h and ATP (4 mM) for 2 h to induce pyroptosis. The results indicated that BMDMs from SETDB1 cKO mice exhibited higher levels of inflammatory cytokines and chemokines IL-1β, IL-6, TNF-α, MCP-1, CXCL2, and CXCL10, and increased secretion of inflammatory factors IL-1β, IL-6, and TNF-α compared to BMDMs from SETDB1fl/fl mice (Fig. 6A and B). Western blot analysis revealed increased expression of Casp1-p20, GSDMD-FL, and GSDMD-N in BMDMs from SETDB1 cKO mice (Fig. 6C–F). These findings suggest that SETDB1 protects against macrophage pyroptosis and the associated inflammatory response by negatively regulating P2RX7 expression and the Caspase-1/GSDMD pathway.

Discussion
SETDB1 has emerged as a crucial epigenetic regulator in controlling inflammatory responses and maintaining tissue homeostasis. In this study, we identified a protective role of macrophage SETDB1 in liver IRI. The suppression of the P2RX7/Caspase-1/GSDMD signaling cascade by SETDB1 highlights a novel link between histone methylation and the regulation of pyroptosis, suggesting that SETDB1 may serve as a molecular brake to restrain excessive inflammasome activation during sterile inflammation.
Our findings further suggest that the absence of SETDB1 amplifies macrophage pyroptotic signaling, promoting the release of pro-inflammatory cytokines and recruitment of immune cells, which in turn exacerbates liver damage. These results underscore the importance of SETDB1-mediated epigenetic control in balancing immune activation and tissue injury. Understanding this regulation provides new insight into the crosstalk between chromatin-modifying enzymes and innate immune signaling pathways during hepatic stress. Such knowledge not only deepens our understanding of liver IRI pathophysiology but also points to SETDB1 and its downstream P2RX7/Caspase-1/GSDMD axis as promising therapeutic targets for inflammation-modulating interventions.
SETDB1, an epigenetic methyltransferase, is crucial for regulating immune responses through its suppressive effects on inflammatory gene expression (Griffin et al., 2021). Tumor-related studies have demonstrated that the epigenetic silencing of immune genes by SETDB1 can promote immune evasion in cancers (Zhang et al., 2021). In the context of intestinal inflammation, SETDB1 is critical for maintaining epithelial homeostasis, supporting differentiation and survival of intestinal epithelial cells, and preserving genomic stability in intestinal stem cells, thereby preventing inflammation (Južnić et al., 2021; Wang et al., 2020). Furthermore, by suppressing transposable elements to preserve genomic stability, SETDB1 plays a critical role in preventing excessive immune activation and DNA damage-induced cell death during hypoxia (Park et al., 2023). In ischemic hypoxic conditions, SETDB1 has been shown to mitigate renal IRI by targeting the HP1β-SESN2 axis (Xia et al., 2024). In hepatocytes, SETDB1 participates in inflammatory regulation during liver IRI via the JNK/p38 pathway, targeting apoptosis-signal-regulating kinase 1 (ASK1) (Xia et al., 2023). Despite these insights, the specific role of SETDB1 in macrophages during liver IRI has remained largely unexplored. Given the central role of macrophages in liver immune responses, understanding the function of SETDB1 in this context is critical for identifying therapeutic strategies to reduce inflammation and tissue damage in liver IRI.
P2RX7, a purinergic receptor, is activated by extracellular ATP during inflammation and tissue injury, playing a pivotal immunomodulatory role through various signaling pathways (Di Virgilio et al., 2017; Ousingsawat et al., 2015). It is fundamental for macrophage functions in innate immune response, facilitating processes such as chemotaxis, migration, phagocytosis, and cell lysis (Ousingsawat et al., 2015; Surprenant et al., 1996). Notably, high levels of extracellular ATP activate P2RX7, which enhances neuronal activity and regulates ferroptosis and oxidative stress via the extracellular signal-regulated kinase (ERK)/glutathione peroxidase 4 (GPX4) signaling pathway (Li et al., 2024). P2RX7 has also been implicated in the release of inflammatory mediators and cell pyroptosis in conditions such as chronic nonbacterial osteomyelitis (Charras et al., 2024). Among the downstream pathways regulated by P2RX7, the caspase-1/GSDMD signaling pathway has emerged as a critical player in the inflammatory regulation during IRI across multiple tissues, including the liver kidney, intestine, retina, neurons, brain, and myocardium (Jia et al., 2020; Kolachala et al., 2021; Luo et al., 2022; Wang et al., 2022; Yan et al., 2023; Yanpiset et al., 2023). Here, we found that SETDB1 regulates liver IRI through the P2RX7-mediated Caspase-1/GSDMD pathway. Moreover, SETDB1 may indirectly regulate P2RX7 expression through modulating H3K9 methylation, potentially by altering chromatin accessibility and recruiting heterochromatin proteins such as HP1. Understanding these interactions could yield valuable insights into the immunomodulatory roles of SETDB1 and P2RX7 in liver IRI.
In conclusion, our findings reveal that macrophage SETDB1 plays a critical role in liver IRI. By reducing pyroptosis and limiting pro-inflammatory macrophage infiltration through the P2RX7/Caspase-1/GSDMD pathway, SETDB1 protects hepatic tissue from IRI-induced damage. These results suggest that enhancing SETDB1 activity or stability, as well as pharmacologically inhibiting the P2RX7/Caspase-1/GSDMD signaling axis, may represent promising therapeutic strategies to alleviate liver inflammation and injury. This molecular insight into the inflammatory regulation in liver IRI lays the foundation for novel therapeutic approaches aimed at reducing liver inflammation and injury triggered by ischemia-reperfusion. So further exploration of this pathway may uncover new strategies for targeted inflammation-modulating therapies.
Innovation
Research on liver IRI has mainly focused on inflammatory responses and immune cell infiltration, while the role of epigenetic regulation remains unclear. This study demonstrates that SETDB1 protects against liver injury by suppressing the P2RX7/Caspase-1/GSDMD pathway in macrophages, linking epigenetic control to pyroptosis regulation. These findings provide new mechanistic insight and suggest a potential therapeutic strategy for preventing liver transplantation complications.
Materials and Methods
Reagents
Granulocyte-macrophage colony-stimulating factor (GM-CSF, Peprotech 315-03), Collagenase type IV (STEMCELL TECHNOLOGIES 07909), Percoll (Cytiva 17-0891-09), Mouse IL-1β ELISE kit (Abclonal RK00006), Mouse IL-6 ELISE kit (Abclonal RK00008), Mouse TNF-α ELISE kit (Abclonal RK00027), CD45-APC (Biolegend 103112), F4/80-PE (BD 565410), CD11b-PE-CY7 (Biolegend 101216), CCR2-BV421 (BD 747963), CD80-FITC (Biolegend 104706), Hanks (Solarbio H1020), D-Hanks (Solarbio H1045), Casp1-p20 (Cell Signaling Technology D57A2), GSDMD (Abcam ab209845), MPO antibody (Abcam ab208670 Dilution ratio: 1:3000), HRP Goat Anti-Rabbit (Shanghai Huilan Biotechnology Co., Ltd. RCA054), Diaminobenzidine (DAB) Chromogenic Reagent Kit (Shanghai Huilan Biotechnology Co., Ltd.), TUNEL reagent (Shanghai Huilan Biotechnology Co., Ltd. RCT-50R Dilution ratio: 1:50), 4′,6-diamidino-2-phenylindole dihydrochloride (DAPI; Shanghai Huilan Biotechnology Co., Ltd. RC05), Primary antibody (F4/80) (CST 70076S Dilution ratio: 1:500), Primary antibody (CD86) (CST 19589S Dilution ratio: 1:400), HIF1a Antibody (28 b) (Santa Cruz Biotechnology Dilution ratio: 1:100), Di-Methyl-Histone H3 (Lys9) Rabbit Monoclonal Antibody (Beyotime AF2314 Dilution ratio:1:1000), Histone H3K9me3 antibody (pAb) (Active Motif Dilution ratio:1:1000), Histone H3 Rabbit pAb (ABclonal A2348 Dilution ratio:1:1000), P2RX7 Polyclonal antibody (Proteintech 28207-1-AP Dilution ratio:1:1000).
Mice
Lyz-Cre transgenic mice were purchased from GemPharmatech CO., Ltd and genotyped. The SETDB1fl/fl mice were generated by the CRISPR-Cas9 system (GemPharmatech) and were backcrossed with Lyz-Cre transgenic mice to generate mice with selective deletion of Setdb1 in myeloid cells (SETDB1 cKO). All mice were maintained under controlled conditions (22°C, 50% humidity, 12 h light/dark cycle, with lights on at 7:00 AM). The animal care and experimental protocols conducted in this study were approved by the Animal Care and Use Committee of Huazhong University of Science and Technology.
Liver IRI model
Male mice (6–8 weeks old), underwent a 70% liver ischemia/reperfusion model to induced IRI. Mice were anesthetized, and the arterial and portal venous blood supply to the cephalad lobes of the liver was interrupted with a vascular clamp, the clamp was removed after 40 min of ischemia, and the mice were euthanized by cervical dislocation after 6 h of reperfusion. The sham group underwent the same procedure without blocking the blood supply. Oxidized ATP (150 μg/mouse) or saline was administered intraperitoneally daily for one week prior to ischemia. CL (200 μL/mouse) was injected intraperitoneally 24 h before liver ischemia modeling.
Liver enzyme assay
Serum was collected from the mouse eyeball blood, and levels of ALT and AST in serum were measured using a biochemical analyzer.
Culture of BMDMs
BMDMs were cultured by isolating bone marrow cells from the femur and tibia of 6–8-week-old mice. Cells were cultured in DMEM medium supplemented with 10% fetal bovine serum (FBS) and GM-CSF (10 ng/mL) for 6 days, with medium changes on day 3. After 8 h of stimulation with LPS (500 ng/mL) and 2 h of stimulation with ATP (4 mM), cells and culture supernatants were collected for further analysis.
Isolation of primary mouse macrophages
Mice were anesthetized, and the portal vein was first injected with Hank's balanced salt solution (without calcium and magnesium) to rinse the blood in the liver. The liver was then perfused with D-Hanks digestive solution containing calcium, magnesium, and 0.5 mg/mL collagenase type IV. After harvesting, the liver was torn apart, and the collected cell suspension was filtered through a 70 μm filter. The filtered cells were centrifuged at 4° 50 g for 5 min, and the supernatant was collected for the separation of macrophages. Macrophages were separated using 25% and 50% Percoll solutions.
ELISA
The secretion of cytokines/chemokines (IL‐1β, IL‐6, TNF‐α) was measured by ELISA, following the manufacturer’s protocols.
H&E staining and histological analysis
Liver tissues were fixed in paraformaldehyde, dehydrated, embedded, sectioned, and stained with H&E. The severity of liver IRI was assessed using Suzuki’s criteria, which include scoring for follows: congestion (none = 0, minimal = 1, mild = 2, moderate = 3, severe = 4), vacuolization (none = 0, minimal = 1, mild = 2, moderate = 3, severe = 4), and necrosis (none = 0, single cell necrosis = 1, < 30% = 2, 30–60% = 3, > 60% = 4); scores for each parameter ranged from 0 to 4, with a maximum score of 12.
Immunofluorescence staining for paraffin sections
Four-micron-thick paraffin-embedded tissue sections were deparaffinized, and antigen retrieval was performed. Sections were blocked with 3% hydrogen peroxide solution and 3% bovine serum albumin (BSA). Primary antibodies with a specific dilution ratio were incubated on the tissue in a dark, humid chamber at 4°C overnight, followed by incubation with secondary antibodies for 1 h at room temperature. The newly formulated fluorescent color-developing liquid was dropped onto the tissue, washed with PBS to stop the color developing, and then the above steps were repeated to incubate another primary antibody. The slide is then fitted with DAPI to stain the nucleus and mount the slides with an anti-fade mounting medium. The slices were then viewed under an inverted fluorescence microscope, and images were captured.
Immunohistochemistry on paraffin sections
Four-micron-thick sections were prepared from paraffin-embedded tissues, following deparaffinization of the sections, and then the antigens were retrieved. Then, the tissue was blocked with 3% hydrogen peroxide solution, and the tissue was covered with diluted primary antibody and incubated in a humidified room at 4°C overnight; the corresponding secondary antibody (HRP label) was added to cover the tissue, and incubated at room temperature for 60 min. Then, the tissue was stained with freshly prepared DAB staining solution, and the nucleus was counterstained with Harris hematoxylin. Finally, the tissue slides were dehydrated and mounted.
TUNEL immunofluorescence staining
Four-micron-thick sections were prepared from paraffin-embedded tissues, following deparaffinization of the sections, and then the antigens were retrieved. Then the TUNEL reagent was added and placed flat in a dark and humid chamber at 37°C for 1 h. The DAPI staining solution was incubated in the dark at room temperature on the tissue for 10 min. Then the slides were mounted with an anti-fading mounting medium, and the sections were observed and photographed under an inverted fluorescence microscope.
Quantitative real-time PCR
RNA was extracted from tissues and reverse-transcribed into cDNA. The expression of target genes was quantified by real-time PCR using SYBR Master Mix. PCR results of β-actin were used as internal controls. The primer sequences used in this study are listed in Table 1.
Sequences of primers used for quantitative real-time PCR analysis

Flow cytometry
Cells isolated from Percoll gradients were resuspended, fixed with 0.4% paraformaldehyde for 15 min, and incubated with Fc block for 15 min. Then added flow antibody to avoid light and incubated for 30 min. After centrifugation, the precipitated cells were re-suspended with flow fixing solution for on-machine detection. The antibodies used were CD45-APC, F4/80-PE, CD11b-PE-CY7, CCR2-BV421, and CD80-FITC.
Western blots
Total protein was extracted from BMDMs stimulated with LPS and ATP, separated by SDS-PAGE, and transferred to membranes. Membranes were probed with antibodies against Casp1-p20 and GSDMD, and β-actin was used as a loading control. For densitometric analysis, the intensity of each band was quantified using Image J software. A rectangular region of interest was drawn around each band. Background intensity was not subtracted during densitometric analysis, and the raw band intensity values were normalized to β-actin. The resulting relative protein expression levels were used for statistical analysis and presented in the figures.
Cell culture
Cell line L02 was obtained from CCRID. All cell lines were maintained in DMEM containing 4.5 g/mL Glucose and
Establishment of the hypoxia/reoxygenation injury model
L02 cells in the logarithmic growth phase were seeded into culture plates at an appropriate density. After the cells adhered and grew to confluence, the original medium was discarded, and the cells were washed once or twice with PBS. To simulate ischemia-like injury, the culture medium was replaced with glucose-free DMEM, and the cells were subjected to hypoxic conditions.
Hypoxia was induced by placing the cells in a pre-equilibrated hypoxia chamber continuously flushed with a gas mixture of 95% N2 and 5% CO2. The sealed chamber was then placed in a 37°C incubator for 6 h. Following hypoxia treatment, the hypoxic medium was gently aspirated and replaced with normal high-glucose DMEM containing 10% FBS. The cells were immediately returned to the standard incubator (95% air, 5% CO2) and cultured for an additional 12 h for reoxygenation. A normoxic control group was maintained throughout the experiment in normal medium under standard culture conditions, with a medium change performed at the same time points to control for variables.
Statistics
All data are presented as means ± SEM. Statistical significance was assessed using an unpaired t-test or two-way ANOVA. p < 0.05 was considered to be statistically significant. Analysis and graphing were carried out through the GraphPad Prism software (version 9.0).
Authors’ Contributions
L.S., L.J.S., and W.Y. provided development of methodology, acquisition, analysis, and interpretation of data, and statistical analysis; L.S., J.L., and S.C. performed writing and revision of the article; S.C. and P.X.L. performed study concept, design, and review of the article. All the authors read and approved the final article.
Footnotes
Acknowledgments
Thanks to the laboratory equipment of the Experimental Medicine Center of Tongji Hospital, Tongji Medical School, Huazhong University of Science and Technology.
Ethics Approval and Consent to Participate
The mouse care and experimental protocols conducted in this study were approved by the Experimental Animal Welfare and Ethics Committee of Huazhong University of Science and Technology (approval numbers TJH-202303004).
Data Availability Statement
Data, material, and software information supporting the conclusions of this article are included within the article. Data will be made available upon request.
Author Disclosure Statement
The authors declare no competing interests.
Funding Information
This work is supported by the