
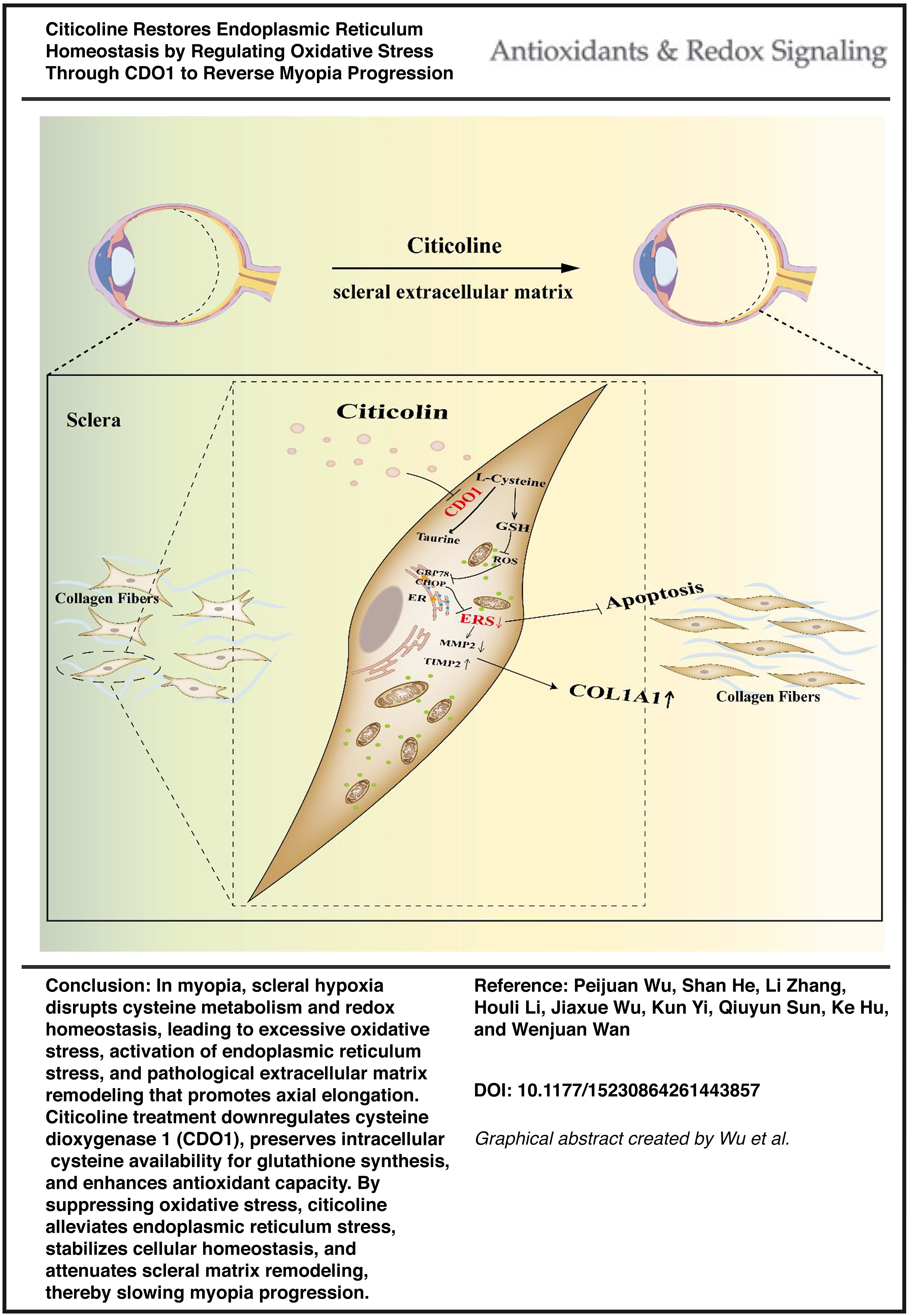
Abstract
Objective:
To investigate whether citicoline alleviates endoplasmic reticulum (ER) stress by modulating oxidative stress in myopia.
Results:
A form-deprivation myopia model in guinea pigs and a hypoxia-induced model in human scleral fibroblasts revealed that citicoline downregulated cysteine dioxygenase 1 (CDO1) expression, increased glutathione, and reduced reactive oxygen species. These changes mitigated oxidative stress, suppressed ER stress, maintained cellular homeostasis, and inhibited extracellular matrix remodeling, leading to reduced axial elongation.
Innovation:
Transcriptomic profiling identified CDO1 as a novel downstream effector of citicoline, linking cysteine metabolism to oxidative and ER stress regulation in myopia.
Conclusion:
Citicoline shows potential to delay myopia progression by alleviating ER stress through CDO1-mediated redox regulation, providing preclinical mechanistic insight into its possible role in myopia control. Antioxid. Redox Signal. 45, 113–132.
Introduction
Myopia is a common refractive error caused by an imbalance between axial length and refractive power, resulting in the focusing of light in front of the retina and blurred vision. It is the most prevalent ocular disorder worldwide, affecting ∼20%–40% of the population, with markedly higher prevalence among adolescents, thereby posing a growing public health concern (Liang et al., 2025). Holden et al. predict that by 2050, ∼4.76 billion people worldwide will be myopic, including one billion individuals with high myopia (Holden et al., 2016). High myopia significantly increases the risk of severe complications such as myopic maculopathy, cataracts, retinal detachment, and glaucoma (Ikuno, 2017). Despite its widespread prevalence and clinical significance, the underlying mechanisms of myopia remain poorly understood.
It is widely recognized that myopia develops as a multifactorial condition resulting from the interplay between genetic predisposition and environmental factors (He and Li, 2023). A hallmark of myopic progression is the persistent elongation of the ocular axis, accompanied by progressive thinning of the sclera, the primary structural tissue for maintaining the shape and biomechanical integrity of the eyeball. The sclera is primarily composed of extracellular matrix (ECM) and fibroblasts, with type I collagen (COL1A1) as the predominant structural protein (Harper and Summers, 2015; Jonas et al., 2023). During myopia development, the scleral collagenous architecture undergoes significant remodeling, driven by an imbalance between collagen synthesis and degradation. This disruption compromises the biomechanical integrity of the sclera against intraocular pressure, thereby promoting progressive axial elongation and exacerbating myopia (Sun et al., 2024). Matrix metalloproteinase-2 (MMP2), a key ECM-degrading enzyme, and its endogenous inhibitor, tissue inhibitor of metalloproteinases-2 (TIMP2), constitute a tightly regulated axis that maintains ECM homeostasis. An imbalance between MMP2 and TIMP2 disrupts collagen turnover, accelerates COL1A1 degradation, and contributes to scleral weakening and pathological tissue remodeling during myopia progression (Liu et al., 2017; Zhuang et al., 2014).
Recent evidence suggests that scleral hypoxia is a common feature of myopic eyes and plays a pivotal role in ECM remodeling (Wu et al., 2018). Hypoxia is known to induce endoplasmic reticulum (ER) stress, primarily by impairing oxygen-dependent protein folding and disrupting calcium homeostasis within the ER lumen. (Bartoszewska et al., 2022). ER stress plays a critical role in maintaining protein-folding homeostasis and overall cellular function. Previous studies have demonstrated that ER stress is involved in the development of numerous fibrotic disea (Lenna and Trojanowska, 2012). Since remodeling of the scleral ECM during myopia shares features with fibrosis, ER stress is considered a contributing factor to scleral changes associated with myopia. ER stress has been observed in both form-deprivation and lens-induced myopia models, suggesting its potential involvement in myopia progression. (Ikeda et al., 2022). In the present study, ER stress was evaluated using the canonical markers GRP78 and CHOP. Oxidative stress was assessed by measuring intracellular reduced reactive oxygen species (ROS) levels together with antioxidant and lipid peroxidation indicators, including superoxide dismutase (SOD) activity and malondialdehyde (MDA) content. Scleral ECM remodeling was monitored through the expression of COL1A1 and the regulatory balance between MMP2 and TIMP2. GRP78 (also known as BiP) is a key ER-resident chaperone involved in protein folding, whereas CHOP is a stress-induced transcription factor associated with apoptosis during prolonged ER stress (Ibrahim et al., 2019; Li et al., 2014). CHOP is considered a critical marker for the transition from protective to detrimental ER stress, and its pro-apoptotic role has been documented in various models of tissue injury (Hu et al., 2019). Under hypoxic conditions, the accumulation of misfolded proteins causes dissociation of GRP78 from ER sensors, leading to activation of the classical unfolded protein response pathways, including PERK-eIF2α, IRE1-XBP1, and ATF6 (Bolland et al., 2021). Additionally, hypoxia-induced mitochondrial dysfunction elevates ROS production, exacerbating ER stress and amplifying the cellular stress response (Aparicio-Trejo et al., 2025; Hu and Zhang, 2021).
Citicoline is an endogenous neuroprotective compound that contributes to membrane stability, neurotransmitter synthesis, and cellular energy metabolism (Qureshi et al., 2016). Citicoline has been widely used in neurodegenerative diseases such as stroke, Parkinson’s disease, and glaucoma, primarily by improving mitochondrial function, suppressing oxidative stress, and inhibiting apoptosis (Baris et al., 2023; Iulia et al., 2017; Vernazza et al., 2022). Recent studies also suggest its potential in promoting retinal and optic nerve repair (Park et al., 2005). However, it remains unclear whether citicoline exerts protective effects on the sclera under myopic stress conditions. In particular, its potential to modulate the stress response of scleral fibroblasts and attenuate myopia-related ECM remodeling has not yet been explored.
Results
Abnormal endoplasmic reticulum function was found in form-deprived myopic guinea pigs
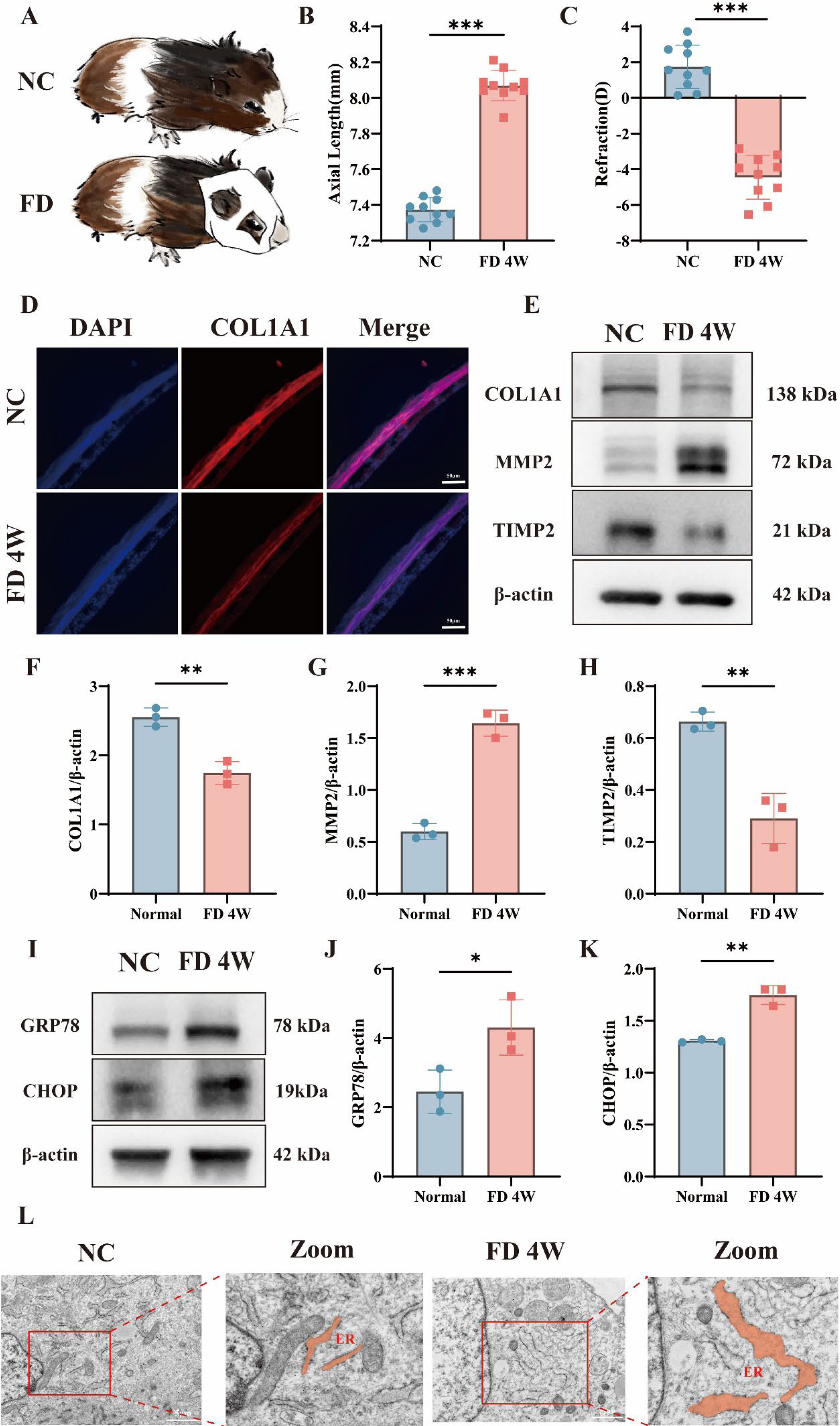
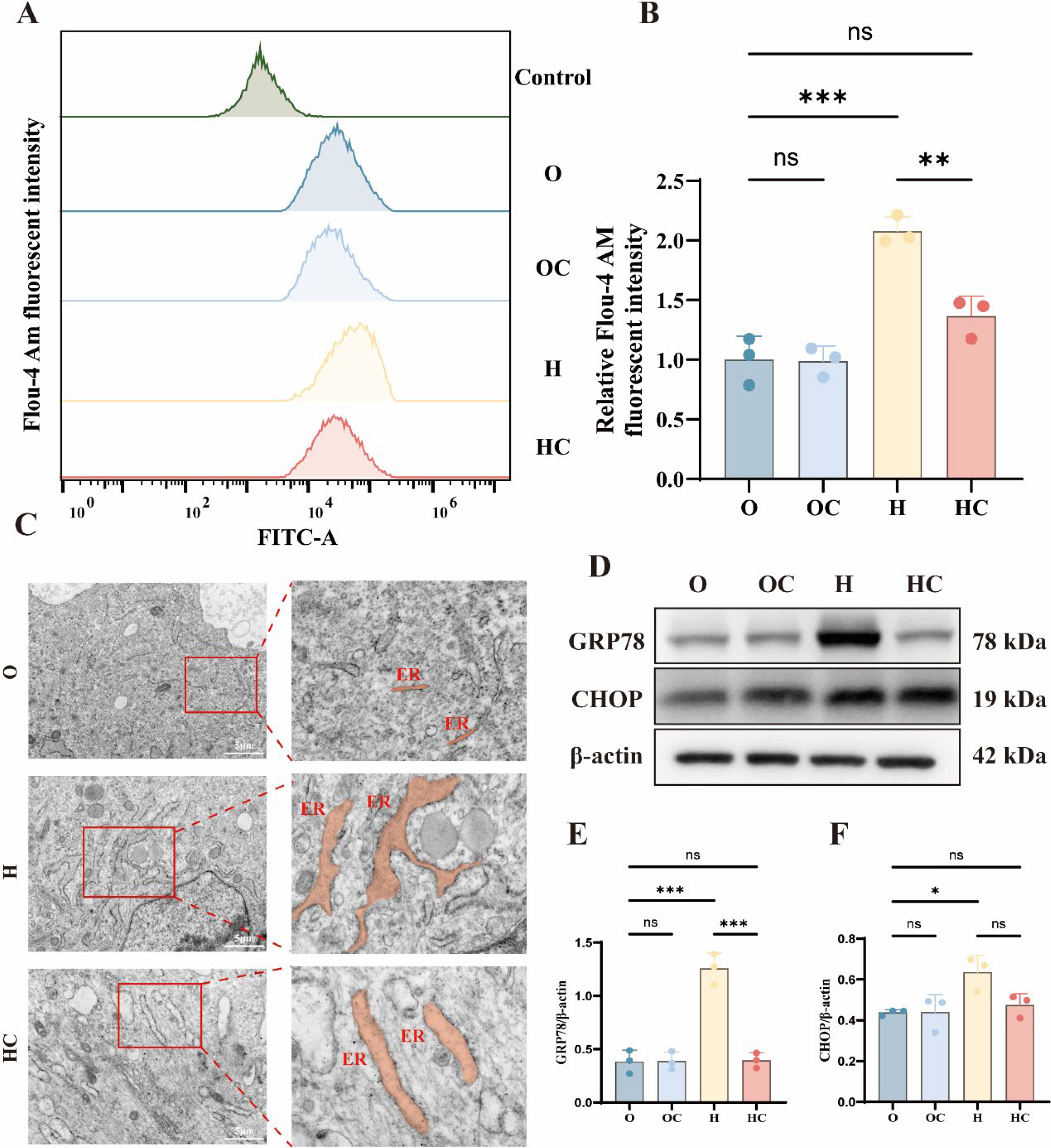
As illustrated in Figure 1A, a form-deprivation myopia model was established by monocular occlusion of the left eye using a white latex facemask. Axial length and refractive error were measured before and after induction using A-scan ultrasonography and an infrared photorefractor. After 4 weeks, the form-deprivation myopia (FD) group showed a significant increase in axial length compared with the NC group (Fig. 1B and C). The FD group also exhibited a marked myopic refractive shift (Fig. 1B and C). Immunofluorescence staining of scleral sections revealed a notable reduction in COL1A1 expression in the FD group (Fig. 1D). Compared with the NC group, the FD group showed significantly decreased COL1A1 and TIMP2 levels, while MMP2 expression was increased (Fig. 1E–H). Compared with the NC group, scleral fibroblasts from the FD group exhibited marked ER dilation and vesiculation. These changes included widened lumina, disorganized cisternae, and perinuclear ER aggregation (Fig. 1L). These morphological features are consistent with ER stress and indicate impaired ER homeostasis in the sclera of form-deprived guinea pigs. These findings indicate that 4 weeks of form deprivation induces ER stress and functional abnormalities in the sclera of guinea pigs. Together, these findings show that form deprivation induces scleral ER stress and ECM remodeling in vivo.
Citicoline attenuated Hypoxia-Induced apoptosis and modulates ECM remodeling in human scleral fibroblast
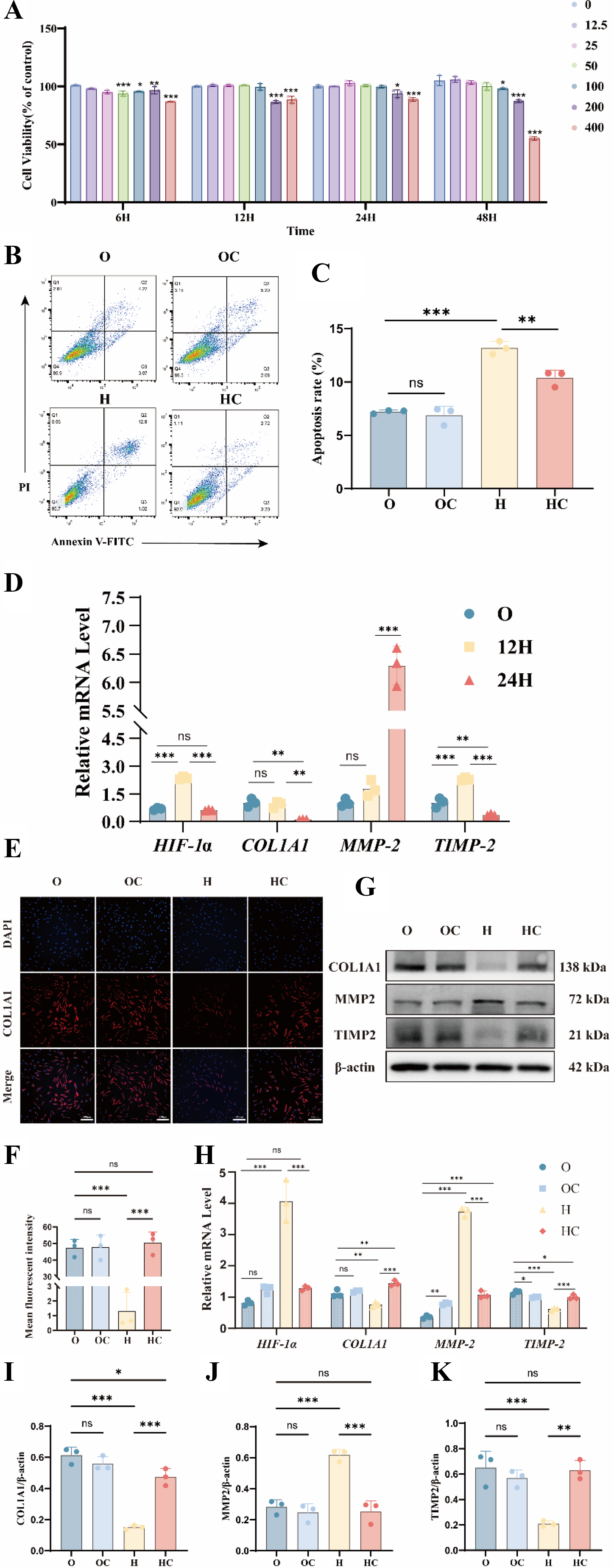
Previous studies have reported the in vitro use of citicoline in ocular or related cell types (Hurtado et al., 2005; Matteucci et al., 2014). Our CCK-8 assays showed that 12.5–200 μM citicoline was safe and biologically active. High concentrations of citicoline (400 μM) consistently reduced cell viability, regardless of treatment duration. At 6 h, moderate to low concentrations (12.5–100 μM) exerted a mild inhibitory effect on cell viability. However, with extended exposure (12–24 h), cell viability approached levels comparable to untreated controls, as shown in Figure 2A. These results indicate that the transient reduction in viability was concentration- and time-dependent. After 12 h of hypoxia, HIF-1α expression showed a slight reduction compared with normoxic controls. Modest changes were also observed in MMP2, TIMP2, and COL1A1 mRNA levels. After 24 h of hypoxia, HIF-1α expression was markedly increased. This increase was accompanied by upregulation of MMP2, decreased TIMP2, and reduced COL1A1 expression, indicating ECM remodeling under hypoxic stress (Fig. 2D). Based on these findings, subsequent experiments were conducted using 100 μM citicoline treatment for 24 h. Flow cytometric analysis showed that citicoline did not induce apoptosis under normoxic conditions in the O and OC groups. Under hypoxic conditions, citicoline significantly attenuated apoptosis (Fig. 2B,C). Under hypoxic conditions, citicoline downregulated HIF-1α and MMP2 mRNA levels. It also attenuated COL1A1 degradation, increased TIMP2 protein expression, and reduced MMP2 protein levels, as shown in Figure 2G–K. In contrast, citicoline did not significantly affect these markers under normoxic conditions. Consistent with these findings, immunofluorescence analysis showed that citicoline restored COL1A1 expression under hypoxia (Fig. 2E and F). These results show that citicoline protects scleral fibroblasts from hypoxia-induced injury and ECM dysregulation.
Citicoline delayed myopia progression and attenuated scleral endoplasmic reticulum stress in Guinea pigs
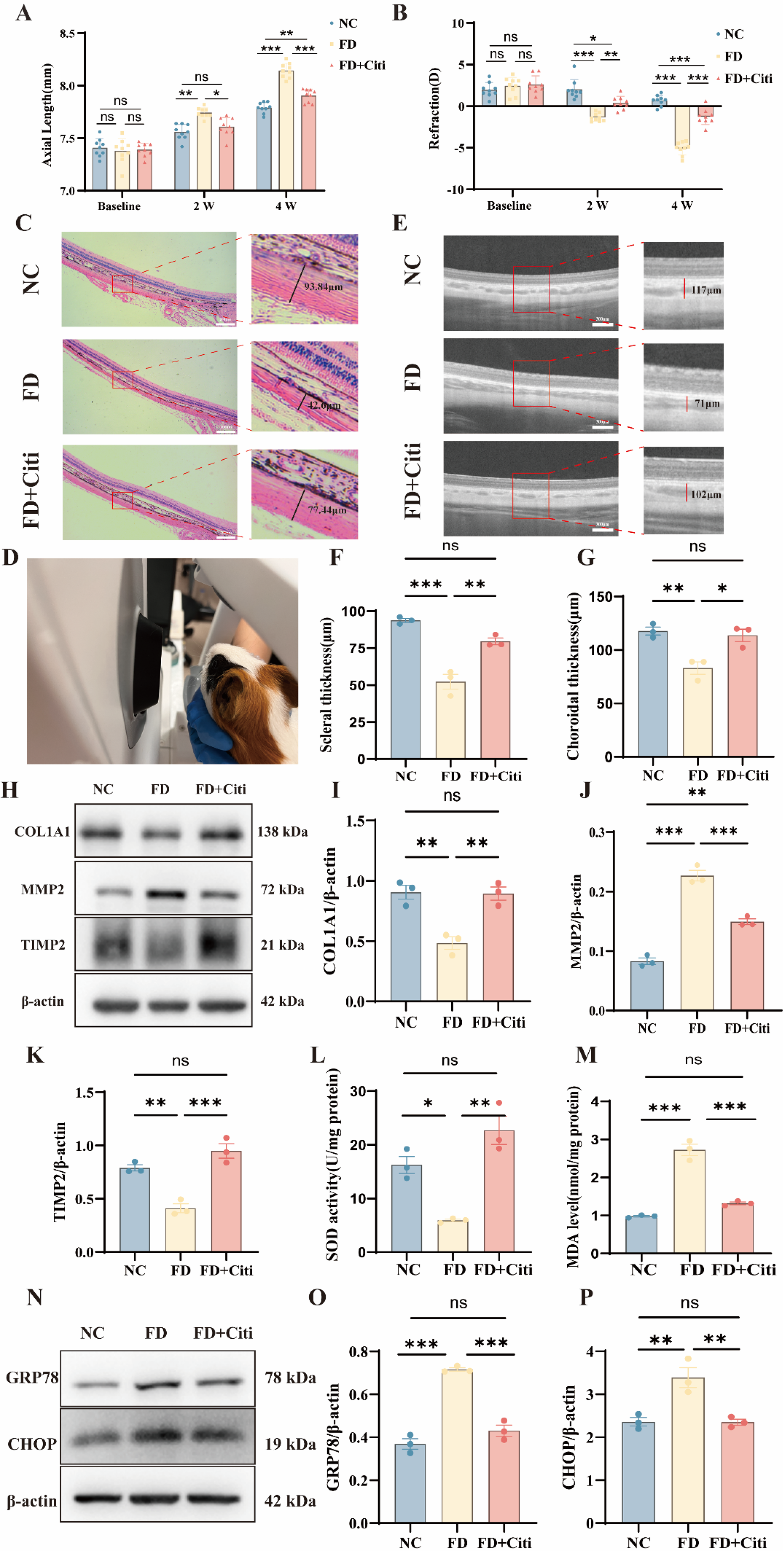
Based on previous studies showing that topically administered citicoline can penetrate ocular tissues, a concentration of 200 μM was selected for topical administration (Carnevale et al., 2019; Parisi et al., 2015; Roberti et al., 2014). This concentration was within the safe range identified in our CCK-8 assays (Fig. 2A). Topical administration of 200 μM citicoline eye drops significantly suppressed axial elongation and the myopic refractive shift during FDM development. At baseline, no significant differences in axial length or refractive error were observed among the groups. After 2 weeks of form deprivation, the FD group showed a pronounced increase in axial length compared with the NC group. The FD group also exhibited a more negative refractive error. Similar changes were observed when the deprived eyes were compared with the contralateral fellow eyes within the same animals. Notably, citicoline treatment in the FD + Citi group significantly reduced axial elongation and myopic progression compared with the FD group. Interocular differences in axial length and refractive error were also significantly reduced, indicating a protective effect. By week 4, the FD + Citi group continued to show attenuation of axial and refractive changes compared with the FD group. However, both parameters remained significantly different from those of the NC group. These findings suggest that citicoline partially delays myopia progression and alleviates form-deprivation-induced ocular changes (Fig. 3A and B and Supplementary Fig. S3). HE staining showed that scleral collagen fibers in the NC group were densely packed and regularly arranged. In contrast, the FD group exhibited scleral thinning and disorganized collagen structure. These alterations were partially alleviated in the FD + Citi group, which showed collagen architecture comparable to that of the NC group (Fig. 3C,F). No structural abnormalities were observed in the corneas of any group, indicating that topical citicoline administration was well tolerated (Supplementary Fig. S1). Optical coherence tomography (OCT) B-scan imaging showed that form deprivation resulted in blurred retinal stratification and indistinct RPE-choroid boundaries, as shown in Figure 3D and E. Choroidal thickness was reduced, and hyporeflective vacuolar signals were decreased. These alterations were markedly improved in the FD + Citi group, suggesting preservation of choroidal structure and vascular integrity (Fig. G). Compared to the FD group, citicoline significantly suppressed MMP2 expression. Citicoline also restored TIMP2 and COL1A1 protein levels (Fig. 3H–K). SOD activity and MDA levels were measured in scleral tissues. Citicoline administration increased SOD activity and decreased MDA levels compared with the FD group (Fig. 3L and M). Western blot analysis showed that citicoline downregulated GRP78 and reduced CHOP expression (Fig. 3N–P). These findings suggest that reduced oxidative stress may contribute to the ability of citicoline to mitigate ER stress during myopia progression. These in vivo findings further support a protective effect of citicoline against myopia-related structural and molecular changes.
Citicoline attenuated oxidative stress and improves mitochondrial function
Given the protective effects of citicoline observed in vitro and in vivo, we next examined whether citicoline modulates oxidative stress in scleral fibroblasts. To determine whether citicoline improves antioxidant capacity under hypoxia, we measured intracellular ROS levels and mitochondrial membrane potential in human scleral fibroblasts (HSFs). Citicoline treatment significantly reduced ROS levels and restored them toward those observed under normoxic conditions (Fig. 4A and B). JC-1 staining revealed a loss of mitochondrial membrane potential in hypoxic cells. Citicoline treatment preserved mitochondrial membrane potential under hypoxic conditions (Fig. 4C and D). These findings indicate that citicoline attenuates hypoxia-induced oxidative stress and preserves mitochondrial function. We next evaluated antioxidant status by measuring SOD activity, MDA levels, and glutathione (GSH) content in the O, OC, H, and HC groups. Under normoxic conditions, citicoline increased SOD activity and GSH levels without affecting MDA content. Under hypoxia, citicoline increased SOD activity and GSH levels and reduced MDA accumulation (Fig. 4E–G). These findings indicate that citicoline improves cellular redox balance under hypoxic conditions.

Citicoline restored endoplasmic reticulum homeostasis in hypoxia-exposed scleral fibroblasts
ER stress is associated with disruption of ER calcium homeostasis. Hypoxia significantly increased intracellular calcium levels in HSFs. Citicoline treatment restored intracellular calcium levels toward those observed under normoxic conditions (Fig. 5A and B). Transmission electron microscopy further revealed ultrastructural changes in ER morphology under hypoxic conditions. In the normoxia group, the ER displayed fine tubular networks with intact membranes and organized luminal structures, as illustrated in Figure 5C. Hypoxia induced ER dilation, membrane loosening, and structural disorganization. Citicoline treatment partially restored ER morphology and reduced these structural abnormalities (Fig. 5C). Under normoxic conditions, GRP78 and CHOP showed low basal expression in HSFs. Hypoxia markedly increased GRP78 and CHOP expression, as shown in Figure 5D–F. Notably, citicoline treatment significantly reduced hypoxia-induced GRP78 and CHOP expression (Fig. 5D–F). Real-time quantitative (RT-qPCR) analysis showed similar changes in GRP78 and CHOP mRNA expression (Supplementary Fig. S2). These observations support a link between citicoline-mediated redox regulation and preservation of ER homeostasis.
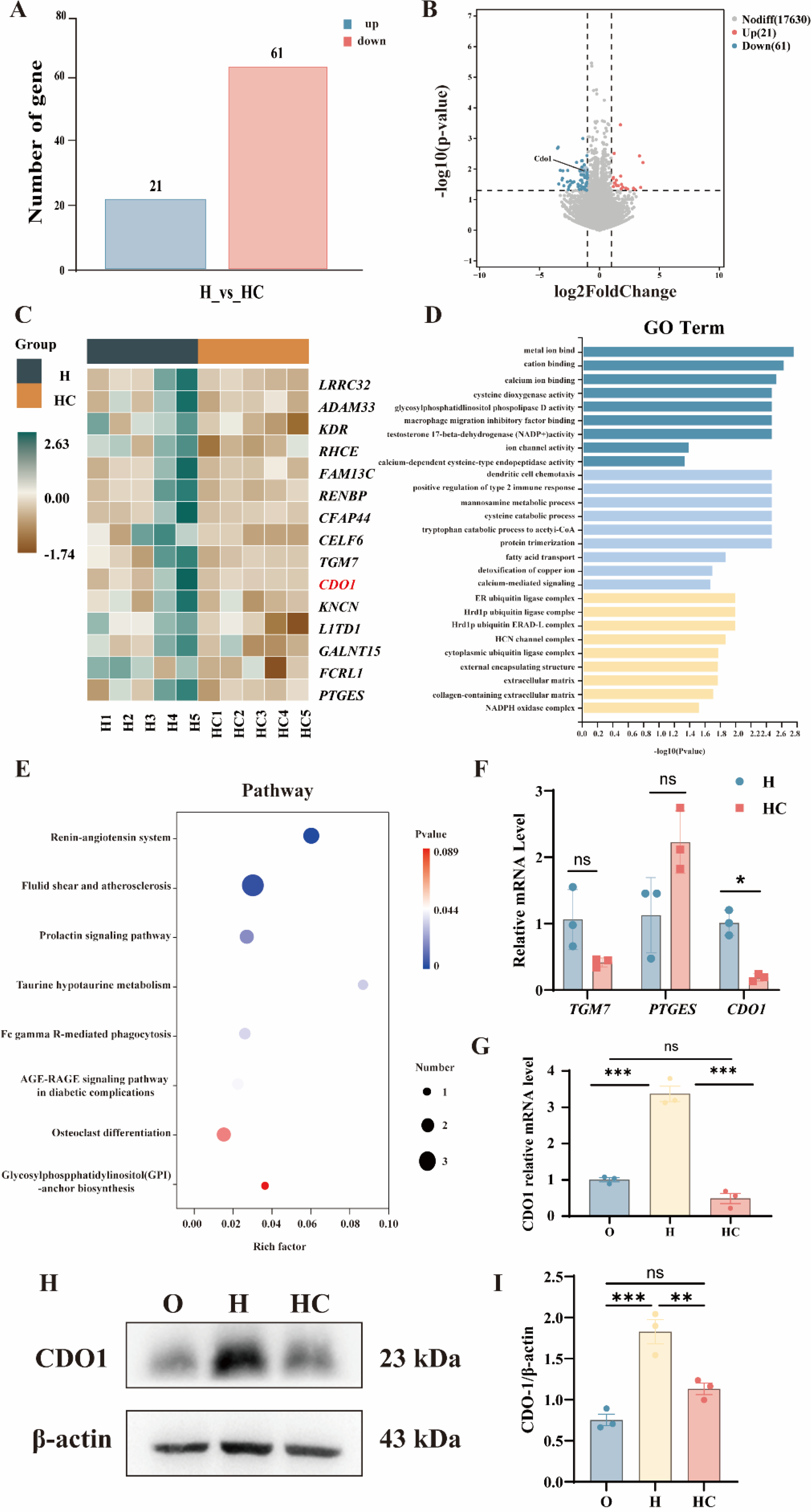
Transcriptomic analysis identified CDO1 as a potential target of citicoline
To identify potential molecular mediators underlying the protective effects of citicoline under hypoxia, we performed RNA sequencing in the H and HC groups. Differential expression analysis identified 82 differentially expressed genes (DEGs), including 21 upregulated and 61 downregulated genes (Fig. 6A). The distribution of these DEGs is illustrated in the bar chart and volcano plot (Fig. 6B). The heatmap of the top 15 DEGs demonstrates distinct expression profiles between the H and HC groups (Fig. 6C). Gene ontology (GO) analysis showed enrichment in biological processes related to cellular stress responses, including “redox process” and “protein folding” (Fig. 6D). Kyoto Encyclopedia of Genes and Genomes pathway analysis identified metabolic and signaling pathways enriched among the DEGs (Fig. 6E). RT-qPCR was performed to validate selected genes among the top 15 DEGs associated with redox processes, including TGM7, PTGES, and cysteine dioxygenase 1 (CDO1). Among these candidates, CDO1 showed the most prominent differential expression between the H and HC groups. Further validation across the O, H, and HC groups showed that citicoline suppressed hypoxia-induced CDO1 upregulation (Fig. 6F–I). Transcriptomic analysis thus identified CDO1 as a candidate mediator associated with the protective effects of citicoline under hypoxia.
CDO1 mediates the protective effects of citicoline on redox and ER stress homeostasis
To examine whether CDO1 mediates the effects of citicoline under hypoxia, we first assessed CDO1 expression across experimental groups. CDO1 expression was markedly increased in hypoxia-exposed scleral fibroblasts compared with normoxic controls. To assess the functional role of CDO1, siRNA-mediated knockdown was performed in hypoxia-exposed scleral fibroblasts. CDO1 knockdown significantly increased intracellular GSH levels compared with the hypoxia group (Fig. 7A). CDO1 knockdown was accompanied by changes in ECM and ER stress markers. Western blot analysis showed that CDO1 knockdown reduced MMP2 expression and restored COL1A1 and TIMP2 levels, as illustrated in Figure 7B–E. CDO1 knockdown also reduced the ER stress markers GRP78 and CHOP (Fig. 7D,F–H). These molecular changes were similar to those observed following citicoline treatment. Conversely, overexpression of CDO1 under hypoxia attenuated the protective effects of citicoline. After CDO1 overexpression, citicoline failed to suppress MMP2 expression or restore COL1A1 and TIMP2 levels (Fig. 7I,J–L). The inhibitory effects of citicoline on GRP78 and CHOP expression were also markedly reduced (Fig. 7I,M–O). These results indicate that CDO1 plays a key role in mediating cellular responses to citicoline under hypoxic conditions.

In vivo modulation of CDO1 influences scleral redox homeostasis and ER stress
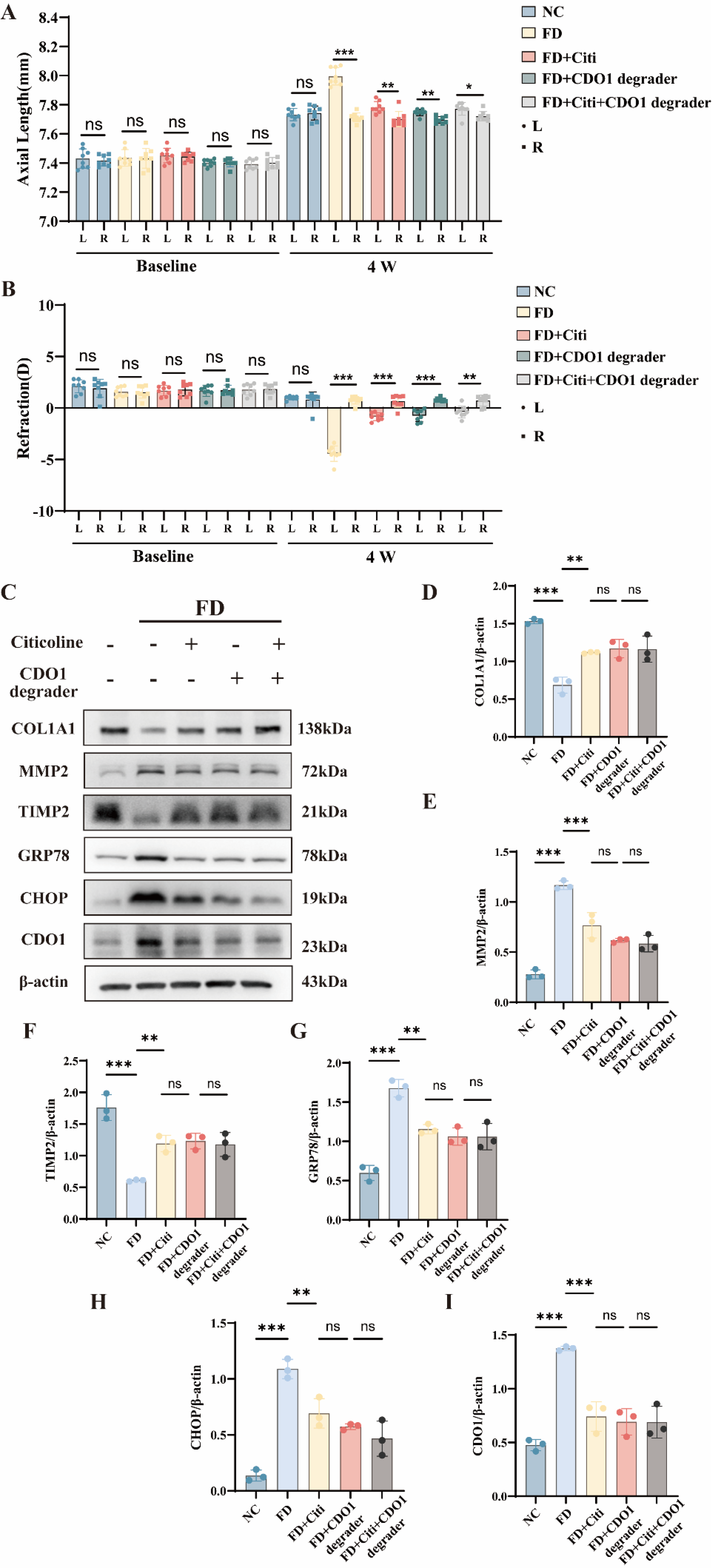
To determine whether CDO1 contributes to myopia progression in vivo, we evaluated the effects of pharmacological CDO1 inhibition in the FD guinea pig model. Based on prior studies, a concentration of 20 μM was selected for in vivo intervention (Tutter et al., 2025). Morphological and biometric assessments showed that pharmacological knockdown of CDO1 significantly mitigated form-deprivation-induced myopic changes. Compared with contralateral control eyes, eyes treated with the CDO1 degrader showed reduced axial elongation and myopic refractive shift (Fig. 8A and B). These changes were similar to those observed in the citicoline-treated group. Western blot analysis of scleral tissues showed that CDO1 knockdown reduced MMP2 expression and restored COL1A1 and TIMP2 levels (Fig. 8C,D–F). CDO1 inhibition also reduced the ER stress markers GRP78 and CHOP (Fig. 8C,G–I). Combined treatment with citicoline and the CDO1 degrader did not further enhance these molecular changes. These findings demonstrate that pharmacological inhibition of CDO1 modulates scleral remodeling and ER stress during myopia progression in vivo.
Discussion
Citicoline is an endogenous neuroprotective agent known to modulate mitochondrial function, oxidative stress, and apoptosis (Aminzadeh and Salarinejad, 2019; Martynov and Gusev, 2015). In this study, we examined its effects on oxidative stress and endoplasmic reticulum (ER) stress in a myopia animal model and hypoxia-induced scleral fibroblasts. Citicoline reduced intracellular ROS levels, increased GSH content, and improved mitochondrial membrane potential in hypoxia-exposed scleral fibroblasts. It also decreased the expression of the ER stress in both the FDM model and hypoxia-induced HSFs. These findings indicate that citicoline alleviates oxidative and ER stress in myopic scleral tissue, helping maintain cellular homeostasis and delay myopia progression. The results support a mechanistic basis for considering citicoline as a potential therapeutic strategy for myopia control.
Citicoline’s effects on oxidative and ER stress
Previous studies suggest that hypoxia plays a critical role in scleral ECM remodeling during myopia development (Wu et al., 2018; Xiao et al., 2025). Under hypoxic conditions, excessive ROS accumulation disrupts redox balance, impairs mitochondrial function, and alters ECM metabolism, contributing to axial elongation (Liu and Wang, 2017). In our models, hypoxia was associated with increased ROS levels and reduced mitochondrial membrane potential in scleral fibroblasts. Citicoline markedly attenuated these alterations and helped maintain mitochondrial function and redox homeostasis. These findings are consistent with previous reports describing the antioxidant and mitochondrial protective effects of citicoline in neurodegenerative disease models (Di Simone et al., 2024; Matyja et al., 2008; Qian et al., 2014). Meanwhile, citicoline increased intracellular GSH levels, indicating enhanced cellular redox buffering capacity. Oxidative stress and ER stress are closely linked. These findings suggest that citicoline may also modulate ER stress by restoring redox balance
The role of CDO1 in scleral remodeling
To examine the molecular mechanisms underlying the protective effects of citicoline, we performed transcriptomic analysis in hypoxia-exposed scleral fibroblasts with or without citicoline treatment. Among the differentially expressed genes, CDO1 was selected for further study based on its consistent expression across RNA-seq, RT-qPCR, and protein analyses. CDO1 regulates cysteine metabolism and intracellular redox balance. It influences cellular responses to oxidative stress (Chen et al., 2022; Conrad and Friedmann Angeli, 2015; Yang et al., 2014). Redox regulation has been closely linked to ECM remodeling in various pathological conditions (Eble and de Rezende, 2014; Grosche et al., 2018). In our study, changes in CDO1 expression were associated with altered levels of ECM-related genes, including COL1A1, MMP2, and TIMP2. In vitro, CDO1 knockdown and overexpression altered the expression of these matrix-related markers. Similarly, pharmacological inhibition of CDO1 in vivo was associated with changes in ECM components in scleral tissue. These findings suggest that CDO1 may contribute to scleral matrix remodeling during myopia progression. Similar roles have been reported in other disease contexts, where CDO1 is involved in metabolic regulation and cellular homeostasis.
Interplay between redox balance, ER stress, and ECM remodeling
We next examined how oxidative stress and ER stress relate to ECM remodeling in scleral fibroblasts. Oxidative stress and ER stress are closely interconnected, particularly under hypoxic conditions. Redox imbalance disrupts protein folding and induces ER stress (Díaz-Bulnes et al., 2020; He et al., 2024). In myopia, these stress responses have also been implicated in abnormal scleral remodeling and ECM dysregulation (Ikeda et al., 2022). In our study, hypoxia was associated with increased ROS levels and ER stress activation in scleral fibroblasts. Notably, CDO1 may represent a key node within this process. CDO1 regulates cysteine metabolism and influences intracellular redox balance. This may affect downstream stress responses and cellular homeostasis. In our models, modulation of CDO1 expression was associated with coordinated changes in GSH levels, ROS accumulation, and stress-related markers. These findings suggest that CDO1 may act as a mediator linking redox imbalance and ER stress to ECM remodeling (Chen et al., 2022; He et al., 2025).
Translational considerations and limitations
These findings should be interpreted with several limitations in mind. The concentration of citicoline was selected for mechanistic studies under controlled conditions and may not reflect clinically relevant dosing. Most conclusions are based on in vitro experiments using scleral fibroblasts. These models may not fully capture the complexity of the in vivo microenvironment. Although pharmacological modulation of CDO1 was performed in vivo, the absence of genetic models limits the strength of the mechanistic evidence. The pharmacokinetics of citicoline in ocular tissues also remain poorly understood. It remains unclear whether comparable drug concentrations can be achieved in scleral tissue after topical administration. This uncertainty may affect the interpretation of the translational relevance of the current findings.
Future directions
Citicoline alleviates oxidative and ER stress in scleral fibroblasts and in a myopia model. Our findings suggest that CDO1 is involved in linking redox imbalance to cellular stress responses and may contribute to ECM remodeling during myopia progression. These results point to a potential mechanism how metabolic and stress-related pathways interact in scleral remodeling. Further work is needed to clarify the upstream regulation of CDO1 and its downstream effects on ECM homeostasis.
Translational Perspective
Citicoline demonstrates preclinical potential as a therapeutic candidate for myopia by modulating oxidative stress and endoplasmic reticulum homeostasis, thereby influencing scleral ECM remodeling. However, several barriers remain, including limited understanding of ocular pharmacokinetics, uncertainty regarding achievable scleral drug concentrations, and the lack of long-term safety data. Further studies are required to define optimal dosing strategies, characterize tissue distribution, and validate efficacy in clinically relevant models before translation to human applications.
Innovation
This study identifies CDO1 as a novel downstream target of citicoline in the sclera and demonstrates that its downregulation mitigates oxidative and ER stress, thereby preserving ECM homeostasis. These findings broaden the therapeutic potential of citicoline in myopia prevention and reveal a new molecular target for intervention.
Materials and Methods
Electronic laboratory notebook was not used in this study.
Animals
All animal experiments in this study were approved by the Ethical Review Committee for Laboratory Animals of Chongqing Medical University (Approval No. [IACUC-CQMU-2025-06003]) and were conducted in accordance with the ARVO Statement for the Use of Animals in Ophthalmic and Vision Research and the ARRIVE guidelines. A total of 45 guinea pigs aged 2–3 weeks were obtained from Guizhou Kangnuo Experimental Animal Breeding Company Limited (License No.: SCXK (Gui) 2023-0001). All animals were housed in standard cages within the Laboratory Animal Center of Chongqing Medical University (Chongqing, China) at a controlled temperature of 22 ± 2°C. Animals were housed in standard cages at 22 ± 2°C under a 12-h light/dark cycle (lights on 9:00 a.m. to 9:00 p.m.), with light intensity of 100–200 lux during the light phase.
Establishment of Form-Deprivation myopia animal model
27 guinea pigs were randomly assigned to three groups: Normal control (NC) group, FD group, and FD with citicoline treatment (FD + Citi) group using a random number generator to minimize selection bias. The NC group received no intervention. In the FD and FD + Citi groups, the left eye was covered with a white latex headgear for 4 weeks to induce myopia. Based on CCK-8 assay results indicating enhanced cell viability at 200 μM, citicoline eye drops were administered to the left eye of animals in the FD + Citi group twice daily at 9:00 a.m. and 5:00 p.m. to mimic a circadian dosing schedule. The NC group was administered citicoline-free vehicle drops in the same volume and schedule to control for handling effects. Refractive error, axial length, and choroidal thickness were assessed at 2 and 4 weeks using a photorefractor (Striatech GmbH, Tubingen, Germany), A-scan ultrasound (SW-1000, Tianjin, China), and OCT (VG200D, SVision Imaging Ltd., China), respectively, with all measurements performed by investigators blinded to group allocation.
Drug preparation
Citicoline (Med Chem Express, HY-B0739) was dissolved in phosphate-buffered saline (PBS; Servicebio, G4202). For in vivo experiments, 1 mg of citicoline powder was dissolved in 2 mL PBS to prepare a high-concentration stock solution. Before use, the stock was diluted with 8 mL PBS to yield 10 mL of a 200 μm working solution. Using a pipette, 20 μL of this solution was administered topically to the left eye of guinea pigs in the FD + Citi group, while the other two groups received an equal volume of PBS as a control. For in vitro experiments, 10 mg citicoline powder was dissolved in 4 mL PBS to prepare a 5 mM stock solution, which was diluted to a 1 mM working solution with PBS before further dilution in culture medium to the desired concentrations.
Primary human scleral fibroblast culture and cell model establishment
HSFs were isolated from donor scleral tissues following the method described by Liu et al. (2022). Cells between passages 3 and 6 were used for experiments. Fibroblasts were cultured in DMEM/F12 (Gibco, C11330500BT) supplemented with 20% fetal bovine serum (Sorfa, SX1101) and 1% penicillin-streptomycin (Beyotime, C0222) at 37°C in a humidified incubator with 5% CO2. The culture medium was refreshed every 3 days. For hypoxia modeling, cells were incubated in a tri-gas incubator at 1% O2 for 24 h. Four groups were established: (1) normoxic control (O); (2) normoxia with citicoline treatment (OC); (3) hypoxia (H); and (4) hypoxia with citicoline treatment (HC). These abbreviations were used for group identification throughout the study.
Cell function assay
To systematically evaluate the effect of citicoline on hypoxia-induced changes in human scleral fibroblast function, multiple assays were performed, including assessments of cell viability, apoptosis, oxidative stress, and mitochondrial function.
CCK-8 assay
Cell viability was assessed using the CCK-8 assay to evaluate the effects of citicoline on HSFs. Cells were seeded in 96-well plates at a density of 8 × 10³ cells per well and incubated at 37°C with 5% CO2 for 24 h to allow adherence. Subsequently, cells were treated with varying concentrations of citicoline (0–400 μM) for 6, 12, 24, and 48 h. At each time point, 10 μL of CCK-8 reagent (MedChemExpress, HY-K0301) was added to each well, followed by incubation at 37°C for 2 h. Absorbance at 450 nm was measured using a microplate reader (Arioskan LUX Multimode Microplate Reader, Thermo Scientific) to determine cell metabolic activity and proliferation. Each group was performed in triplicate wells, and experiments were independently repeated three times.
Apoptosis assay
Apoptosis was assessed using an Annexin V-FITC/PI double staining kit (Beyotime, C1383S). Treated cells were trypsinized, washed 1–2 times with PBS, and then stained with Annexin V-FITC and propidium iodide (PI) according to the manufacturer’s instructions. The cells were incubated in the dark at room temperature for 15 min before analysis. Apoptotic rates, including early and late apoptosis, were quantified by flow cytometry.
Mitochondrial membrane potential assay
The effect of citicoline on mitochondrial membrane potential (ΔΨm) was evaluated using the JC-1 probe (Beyotime, C2006). HSFs were seeded in 6-well plates at a density of 5 × 104 cells per well. Following treatment, cells were incubated with JC-1 staining solution prepared according to the manufacturer’s instructions at 37°C for 25 min in the dark. After incubation, cells were washed twice with JC-1 buffer and then imaged using a fluorescence microscope (Leica DMi8, Wetzlar, Germany). The ratio of red to green fluorescence intensity was calculated to assess changes in mitochondrial membrane potential, where increased green fluorescence indicates mitochondrial depolarization (decreased membrane potential), and predominant red fluorescence corresponds to polarized mitochondria with intact membrane potential.
ROS assay
Intracellular ROS levels were measured using the DCFH-DA fluorescent probe (Beyotime, S0034S) to evaluate the modulatory effect of citicoline. HSFs were seeded in 6-well plates at a density of 5 × 104 cells per well. At the end of treatment, cells were incubated with 1 mL of 10 μM DCFH-DA working solution at 37°C for 30 min in the dark. Following incubation, cells were washed twice with PBS to remove excess probe and then stained with Hoechst 33342 (Solarbio, C0031) for 5 min to visualize nuclei. After a final wash, fluorescence images were acquired using a Leica fluorescence microscope. The intensity of green fluorescence served as an indicator of intracellular ROS accumulation.
Measurement of cytoplasmic Ca2+ concentration
Intracellular cytoplasmic Ca2+ levels were measured using the fluorescent calcium indicator Fluo-4 AM (Beyotime, S1061S) following the manufacturer’s instructions. Briefly, cells were washed three times with PBS and incubated with 500 μL of Fluo-4 AM working solution (final concentration: 1 μM) in serum-free basal medium at 37°C for 15 min. After incubation, cells were washed again with PBS. Subsequently, cells were digested with 0.25% trypsin without EDTA, washed with PBS, and analyzed by flow cytometry to determine fluorescence intensity, which reflects intracellular Ca2+ concentration.
SOD, MDA, and GSH assays
Oxidative stress levels in HSFs and guinea pig scleral tissues were assessed using commercial assay kits for SOD (Beyotime, S0101M), MDA (Solarbio, BC0025), and GSH (Solarbio, BC1175). Briefly, HSFs and scleral tissues were lysed or homogenized using pre-chilled extraction buffers provided in the respective kits. The homogenates were filtered and centrifuged at 10,000 ×g for 10 min at 4°C. The resulting supernatants were collected for biochemical analysis. SOD activity, MDA levels, and GSH content were determined according to the manufacturer’s protocols. Results were normalized to total protein concentration and expressed as follows: SOD activity in units per milligram of protein (U/mg protein), MDA concentration in nanomoles per milligram of protein (nmol/mg protein), and GSH content in microliters per milligram of protein (μL/mg protein).
Western blot
Total protein was extracted from scleral tissues and cultured cells using freshly prepared lysis buffer (RIPA: PMSF = 100:1). Protein samples were separated by SDS-PAGE using 10% or 12.5% polyacrylamide gels (Epizyme, PG112; PG113) and then transferred onto polyvinylidene difluoride membranes with pore sizes of 0.45 or 0.22 μm (Millipore, IPVH00010; ISEQ00010). Membranes were blocked with 5% skimmed milk (Beyotime, P0216) for 1 h at room temperature, followed by incubation with primary antibodies overnight at 4°C. After three washes with TBST, membranes were incubated with appropriate horseradish peroxidase-conjugated secondary antibodies for 1 h at room temperature. Protein bands were visualized using an enhanced chemiluminescence detection kit (MedChemExpress, HY-K2005) and quantified using ImageJ software. Protein expression levels were normalized to β-actin. Detailed information on primary and secondary antibodies is provided in Table 1.
List of Primary and Secondary Antibodies Used for Western Blot and Immunofluorescence
CDO1, cysteine dioxygenase 1; COL1A1, type I collagen; HRP, horseradish peroxidase; MMP2, matrix metalloproteinase-2; TIMP2, tissue inhibitor of metalloproteinases-2.
Immunofluorescence
Eyeball pre-embedding and sectioning were performed by Hubei Bios Biotechnology Co. Paraffin-embedded sections were sequentially dewaxed in xylene and rehydrated through graded ethanol. Antigen retrieval was carried out using EDTA buffer in a microwave oven. After cooling, sections were blocked with goat serum for 30 min at room temperature and incubated overnight at 4°C with anti-COL1A1 primary antibody (Proteintech, 67288-1-Ig). The next day, sections were washed three times with PBS and incubated with Alexa Fluor 488-conjugated goat anti-mouse IgG (Proteintech, SA00013-3) for 1 h at room temperature. Nuclei were counterstained using DAPI-containing anti-fade mounting medium (Beyotime, P0131), and fluorescence images were acquired using a fluorescence microscope. For cell immunofluorescence, HSFs were fixed with 4% paraformaldehyde for 15 min, permeabilized with 0.1% Triton X-100 (Solarbio, T8200) for 10 min at room temperature, and washed three times with PBS. Cells were then blocked with goat serum for 30 min and incubated overnight at 4°C with the same COL1A1 primary antibody. Subsequent staining steps were consistent with the tissue immunofluorescence protocol.
Real-time quantitative PCR
Total RNA was extracted using TRIzol reagent (Tripure Isolation Reagent,11667165001) according to the manufacturer’s instructions. Reverse transcription was performed using a commercial cDNA synthesis kit (Accurate Biology, AG11728). Quantitative PCR was carried out using SYBR Green qPCR Master Mix (Servicebio, G3326), with a reaction mixture containing nuclease-free water, gene-specific primers, and SYBR Green premix. Reactions were set up in 96-well PCR plates (Bio-Rad, MLL9651) and run on an ABI 7500 Real-Time PCR System (Applied Biosystems, USA). β-actin was used as the internal reference gene. Relative gene expression levels were calculated using the 2^–ΔΔCt method. Primer sequences used in this study are listed in Table 2.
List of Primer Sequences Used in This Study

Hematoxylin and eosin staining
Pre-embedding and paraffin embedding of eyeballs were performed by Hubei Bios Biotechnology Co. Serial sagittal sections (5 μm thick) were prepared from paraffin-embedded eyeballs. Sections were baked at 65°C for 1 h, dewaxed in xylene, and rehydrated through a graded ethanol series. After rehydration, sections were stained with hematoxylin for 5 min, followed by 0.5% eosin for 1–2 min. Slides were then washed with distilled water, dehydrated through graded ethanol, cleared in xylene, and mounted with neutral resin. Images were captured using a light microscope (Leica DMi8, Germany). Scleral thickness was quantified using ImageJ software by measuring three sections per group at the same distance from the optic nerve.
Transmission electron microscopy (TEM)
Following euthanasia, guinea pig eyeballs were promptly enucleated, and scleral tissues were dissected under a stereomicroscope. The tissues were cut into ∼1 mm³ pieces and immediately fixed in 2.5% glutaraldehyde prepared in 0.1 M phosphate buffer (pH 7.4) at 4°C for at least 24 h. After fixation, samples were rinsed three times with 0.1 M phosphate buffer (15 min each) and post-fixed in 1% osmium tetroxide for 1–2 h. Tissues were then dehydrated through a graded ethanol series, transitioned with acetone, and embedded in Epon 812 epoxy resin. After polymerization, ultrathin sections (70–90 nm) were prepared using an ultramicrotome, mounted on copper grids, and stained with uranyl acetate followed by lead citrate. Sections were examined and imaged using a transmission electron microscope (Hitachi TEM System, Japan). For cellular ultrastructure analysis, treated human scleral fibroblasts were harvested by trypsinization, centrifuged at 1500 rpm for 10 min, and fixed in 2.5% glutaraldehyde overnight at 4°C. Subsequent processing steps were identical to those described for tissue samples.
RNA interference
To investigate the role of CDO1 in Citicoline-mediated regulation of the stress response in HSFs, gene silencing was performed using small interfering RNA (siRNA) targeting CDO1. The siRNA sequence was designed and synthesized by Tsingke Biotechnology Co., Ltd. (Beijing, China), and the specific sequence is listed in Table 3. A non-targeting siRNA (si-NC) was used as a negative control. When HSFs reached 50%–60% confluence, transfection was carried out using Lipofectamine RNAiMAX (Thermo Fisher, 1377850) following the manufacturer’s protocol, with a final siRNA concentration of 50 nM per well. After 24 h, cells were harvested for RNA to evaluate knockdown efficiency via RT-qPCR. Successfully transfected cells were subsequently exposed to hypoxic conditions to assess the effects of CDO1 silencing on scleral ECM remodeling and endoplasmic reticulum stress-related molecules, in comparison with citicoline treatment. All transfections were conducted under serum-free conditions to ensure specificity and transfection stability.
Table of siRNA Sequence for Knocking Down CDO1

Construction of CDO1-overexpression vectors
To investigate the role of CDO1 overexpression in scleral fibroblasts, a CDO1 overexpression plasmid was designed and synthesized based on the CDO1 gene sequence by Tsingke Biotechnology Co., Ltd. (Beijing, China). When HSFs reached 50%–60% confluence, plasmid transfection was performed using Lipofectamine RNAiMAX (Thermo Fisher, 1377850) according to the manufacturer’s instructions, with a final amount of 1.25 μg plasmid DNA per well. After 24 h of transfection, cells were harvested for protein extraction, and overexpression efficiency was evaluated by Western blot analysis. Successfully transfected cells were subsequently exposed to hypoxic conditions to assess the effects of CDO1 overexpression on scleral ECM remodeling and the expression of endoplasmic reticulum stress–related molecules, in comparison with the citicoline-treated group. All transfection procedures were conducted under serum-free conditions to ensure specificity and transfection stability.
Pharmacological degradation of CDO1 in vivo
To pharmacologically modulate CDO1 expression in vivo, a previously reported small-molecule VHL molecular glue degrader targeting CDO1 NVS-VHL720 (Med Chem Express, HY-160525) was employed, as described in A small-molecule VHL molecular glue degrader for CDO1 (Tutter et al., 2025). Forty guinea pigs were randomly assigned to five groups: Normal control (NC) group, form-deprivation myopia (FD) group, FD with citicoline treatment (FD + Citi) group, FD + Citi with CDO1 degrader treatment (FD + Citi+CDO1)group, and FD with CDO1 degrader treatment (FD + CDO1) group using a random number generator to minimize selection bias. The CDO1 degrader was dissolved in DMSO and administered via subconjunctival injection at a concentration of 20 μM in a total volume of 5 μL per eye, once daily for four weeks.
Data acquisition
The RNA-seq datasets were obtained from the Gene Expression Omnibus (GEO) database: GSE317152.
Study design and rigor
Animals were randomly assigned to experimental groups using a random number generator to minimize selection bias. Investigators performing ocular biometric and histological assessments were blinded to group allocation. Sample sizes were determined based on prior studies in our laboratory and published literature using form-deprivation guinea pig models, as well as established experimental practices. These studies provide a reference for detecting differences in axial length, refractive error, and scleral remodeling (Kang et al., 2023). Formal power analysis was not performed.
Statistical analyses
All statistical analyses were conducted using GraphPad Prism version 9.5.0 (GraphPad Inc., USA). Data normality were evaluated with the Shapiro–Wilk test. For comparisons between two independent groups, unpaired two-tailed Student’s t-tests were employed when the data followed a normal distribution, whereas the Mann–Whitney U test was applied for non-normally distributed data. For comparisons among multiple groups, one-way or two-way analysis of variance was performed, followed by Tukey’s multiple comparisons test. For nonparametric data, the Kruskal–Wallis test with Dunn’s multiple comparisons test was used. All experiments were independently repeated at least three times. Results are expressed as the mean ± standard deviation, unless specified otherwise. Statistical significance was defined as a p value less than 0.05. Significance levels are represented as follows: *p < 0.05, **p < 0.01, ***p < 0.001; ns indicates no significant difference. Exact p values for all statistical comparisons are provided in Supplementary Tables S1, S2, S3, S4, S5, S6, S7, and S8.
Footnotes
Author Disclosure Statement
Peijuan Wu and Shan He participated in the writing of the manuscript. Li Zhang made substantial contributions to the conception or design of the study, as well as to the acquisition, analysis, or interpretation of data. Houli Li and Jiaxue Wu drafted the manuscript or critically revised it for important intellectual content. Qiuyun Sun and Kun Yi agreed to be accountable for all aspects of the work, ensuring that any questions relating to the accuracy or integrity of any part of the study were appropriately investigated and resolved. Wenjuan Wan and Ke Hu provided financial support for the research. The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article.
Funding Information
This work was supported by the
Supplemental Material
Supplemental Material
Abbreviations Used
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.