
Abstract
The goal of assessing psychosocial stress as a process and outcome in naturalistic (i.e., field) settings is applicable across the social, biological, and health sciences. Meaningful measurement of biology-in-context is, however, far from simple or straightforward. In this brief methods review, we introduce theoretical framings, methodological conventions, and ethical concerns around field-collection of markers of psychosocial stress that have emerged from 50 years of research at the intersection of anthropology and human biology. Highlighting measures of psychosocial stress outcomes most often used in biocultural studies, we identify the circumstances under which varied measures are most appropriately applied and provide examples of the types of cutting-edge research questions these measures can address. We explain that field-based psychosocial stress measures embedded in different body systems are neither equivalent nor interchangeable, but this recognition strengthens the study of stress as always simultaneously cultural and biological, situated in local ecologies, social–political structures, and time.
Introduction
Stress is an encompassing term that refers to the body’s physiological response to environmental challenges—or stressors—that strain an individual’s ability to maintain homeostasis and undermine adaptive capacity (Ice and James 2007; Pearlin et al. 1981). Stressors can be physical (e.g., altitude), biological (e.g., disease and nutrition), or psychosocial in origin (e.g., discrimination and inequality). Here, we present an integrated approach to field-based measurement and interpretation of psychosocial stress outcomes using biomarkers as biologies-in-context. Our brief review is grounded in decades of biocultural research within anthropology and human biology that considers stress as a primary point to understand the dynamics between sociocultural and biological dimensions of the human experience (Dufour 2006; Hicks and Leonard 2015; Leatherman and Goodman 2020; Stinson et al. 2000). The history and relevance of this biocultural approach to the wider social, biological, and health-related sciences are explained elsewhere (Glass and McAtee 2006; Hertzman and Boyce 2010; Krieger 2001; McEwen 1998, 2012; Meloni 2014; Roberts and Rollins 2020; Sapolsky 1998; Taylor et al. 1997; Worthman and Kohrt 2005). Our goal, rather, is to identify key practices in field-based stress biomarker collections that have emerged from decades of biocultural research, including available options, agreed-on conventions, and ethical considerations.
Basic Principles: Biocultural Measurement of Psychosocial Stress Outcomes
Biocultural frameworks to understand stress processes overlap greatly with biosocial ones (e.g., McDade and Harris 2018, Roberts and Rollins 2020); both are based in a recognition of human biology as dynamically connected to social contexts across the life span and that neither can be fully explained without some consideration of the other. Perhaps the clearest distinction of a biocultural approach is the emphasis on integrating the proximate (highly local) context into research design, and thus the primacy of both fieldwork and theories related to local cultural and biological variation. Almost all biocultural research is field based, anchored in the fundamental understanding that physiological expressions of stress are dynamic and based in proximate context. Physical bodies are situated in time, space, and patterns of practice in the “situated biologies” of real people in the real world (Niewöhner and Lock 2018). Biocultural studies of stress, however, now place greater emphasis on how data collection and interpretation happen, so typically integrate methods such as extended participant observation and interviewing, which elucidate more detailed salient dimensions of the social stress process (Dufour 2006). This work is situated in varied theories of culture as highly localized phenomena that can be both a source of psychosocial stress and a means to alleviate or buffer it. One example of a specific theory is cultural consonance; operationally, it evaluates metrically the extent to which an individual aligns with locally shared norms and practices (Dressler 2017, 2020). For example, Dressler and colleagues (2016) found that lower cultural consonance in social support in Brazil was associated with higher stress measures (based on blood C-reactive protein). Another example is social theories of gender as structural inequality. For example, Nepali women are primarily responsible for household water, so low water access elevates their stress as measured by blood pressure—but not that of their husbands (Brewis et al. 2019). Without such proximal theorization of within-cultural variation, it is difficult to interpret when and why individual measures of stress vary, such as between those in the same household.
Drawing on political–economic theory, biocultural assessments of stress also recognize that the stress experience is often situated within historically inequitable social structures (Leatherman and Goodman 2020). Factors like wealth, power, prestige, social connection, and historical trauma are all important mediators or moderators of the stress process (Link and Phelan 1995; McEwen 1998, 2012; Sapolsky 1998), including what people perceive as stressful (Dressler 1991; Singer et al. 2016). Again, recognition that these unequal structures matter then demands integration into fieldwork of theory and methods for characterizing relevant aspects of the political–economic context and the place of sampled individuals within it, so that measures of psychosocial stress can be interpreted correctly (e.g., Dressler 2005; Flinn and England 1997; Hicks and Leonard 2015). Piperata et al. (2016, 2020) use this approach to understand how long-standing land distribution and economic policies led to widespread food insecurity and, relatedly, psychosocial stress among women in León, Nicaragua. In this context, asking others for food was so stigmatized that when women drew on their social networks to cope with food insecurity, it promoted more (rather than less) psychosocial stress.
Applying anthropological theories of human genetic and developmental adaptation, biocultural assessments of psychosocial stress also recognize that meaningful interpretation of stress outcome biomarkers must consider possible underlying variation in relevant physiological processes (e.g., Martin 2019). Individuals vary in physiological stress responses with a host of individual factors like genetic predispositions, prior environmental exposures, gender, body size or composition, and sleep patterns, to name a few. Relatedly, life history theory suggests that the way bodies identify and respond to stress varies by life stage and may include trade-offs across organ systems and over time (e.g., skeletal growth versus immune function) (see Shattuck-Heidorn et al. 2017). Due to this fully expected variation, individuals may experience—and physically manifest—the same event or environmental condition very differently not just from person to person, but also across time and with changing personal circumstances.
Relatedly, in a biocultural framework, population-level variation in underlying stress physiology is always assumed, a point especially relevant when comparing stress outcome measures beyond a well-defined local context. Many, often irreversible, phenotypic traits in humans reflect highly localized interactions between genotypes and the environment through the process of developmental plasticity. One of the best examples is the extreme variation in the measurable ranges of ovarian hormones in women entering puberty in ecologies with differing energetic demands (Ellison 1996). Thus, comparing stress outcomes across groups requires explicit theories of exactly how and why stress markers might vary, a point carefully developed in the earliest biocultural studies using adaptability frameworks (e.g., Baker et al. 1986) and still adhered to today.
Some Commonly Applied Biomarkers for Field Measurement of Stress Outcomes.
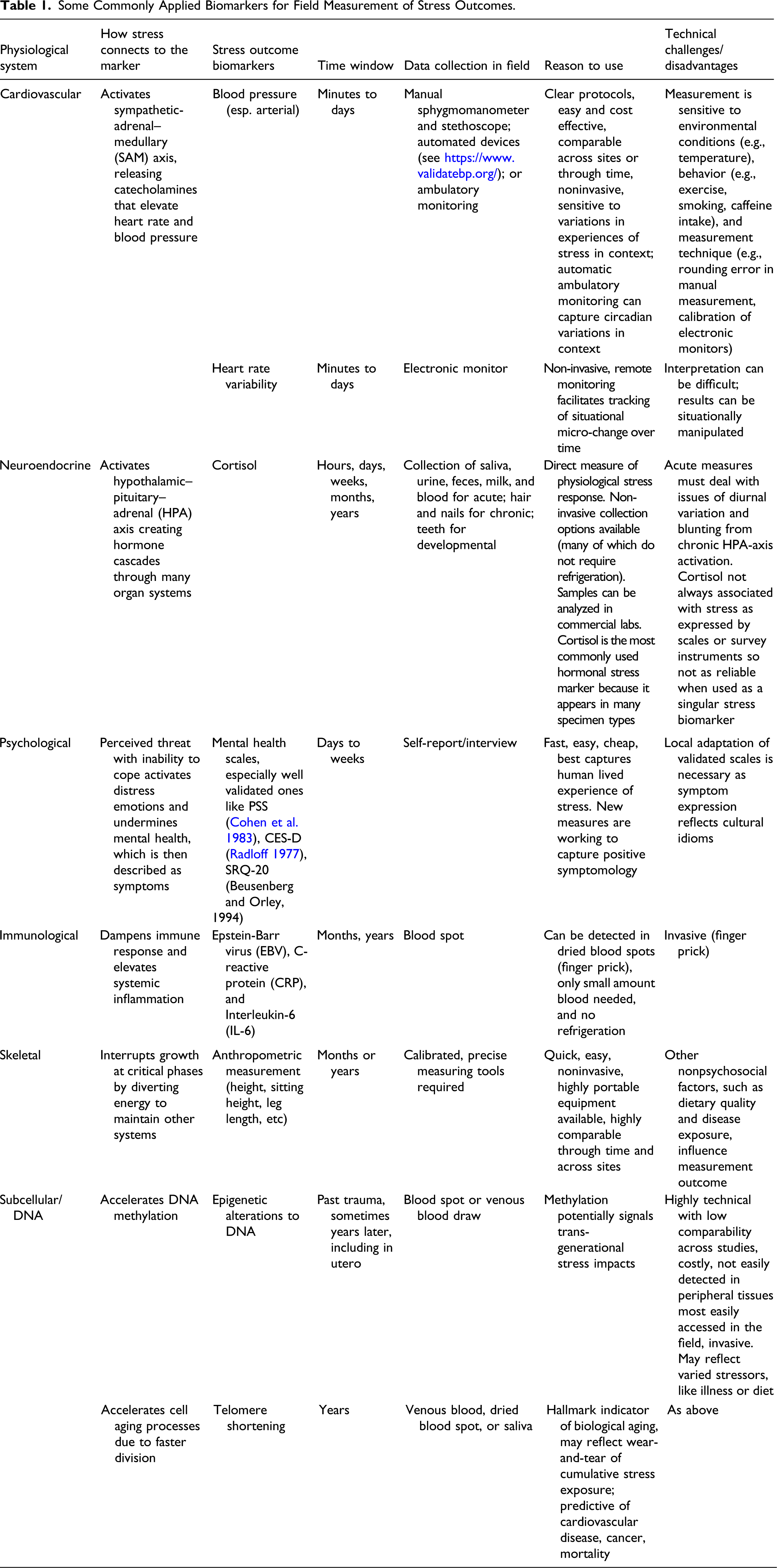
Measuring Psychosocial Stress in Context: Opportunities, Limitations, and Examples
Here, we outline (Table 1) and provide examples of the primary suite of readily available, acceptable biomarker-based methods for assessing psychosocial stress outcomes as biology-in-context. Biomarker here refers to variable, quantifiable expressions of physiological systems. Acceptable means considered by biocultural practitioners as adequately theorized to give meaningful results, sufficiently robust for field-based data collection, and ethically defensible. This list is not exhaustive, but rather highlights the methods widely applied and for which opportunities and limitations are reasonably recognized.
One of the first biomarkers applied in field-based psychosocial stress assessment, heightened blood pressure (e.g., McGarvey and Baker 1979; Scotch 1963), remains widely used because it is noninvasive and easy to measure. However, a lack of understanding as to why it varies individually, temporally, and across populations can lead to misinterpretation (James and Gerber 2018). Heart rate variability is a more recent and closely related measure (e.g., Bell et al. 2019). Both blood pressure and heart rate variability are captured relatively easily in the periphery of the body and have well-established connections to diseases like hypertension, obesity, and type 2-diabetes (Juster et al. 2010; Sapolsky 1998; Steptoe and Kivimäki 2013). This means they can illuminate biological pathways through which psychosocial stress influences human health (Crosswell and Lockwood 2020; Dressler 2004; Worthman and Costello 2009). For example, assessment of social contexts has clarified that greater exposure to market-based lifestyles and the internalization of new but unachievable social and economic expectations of success explain higher blood pressure levels and risk of chronic disease (Bindon et al. 1997; Dressler 1999; Dressler et al. 2005; Pollard et al. 2000; Silva et al. 2016; Steffen et al. 2006; Valeggia and Snodgrass 2015; Waldron et al. 1982). Psychosocial stressors such as racism have also been shown to explain blood pressure variability in the African diaspora better than skin tone or genetic ancestry, pointing to the primacy of sociocultural processes (Gravlee et al. 2005, 2009; Non et al. 2012). Another common and variable source of psychosocial stress relates to gendered expectations, responsibilities, and opportunities. Among Musuo in China, matrilineal social arrangements, which elevate women’s status, are associated with women’s lowered blood pressure while patrilineal arrangements are not (Reynolds et al. 2020).
Neuroendocrine–hormonal biomarkers reflect acute psychosocial stress activation of the hypothalamic–pituitary–adrenal (HPA) axis (Ice and James 2007). For example, in New Zealand, evening salivary cortisol levels were associated with both living in poverty and racial/ethnic discrimination among pregnant women (Thayer and Kuzawa 2014, 2015). Interestingly, their infants exhibited greater cortisol responses to vaccination, suggesting that maternal HPA activation during pregnancy had lasting effects on infants’ HPA axes. Cortisol is the most frequently used neuroendocrine biomarker of psychosocial stress, in part because it can be measured in a range of specimen types (e.g., saliva, blood spots, serum, hair, teeth, finger/toenails) that capture different time scales. The range of options is important as some specimen types are more field-friendly and culturally acceptable than others. In addition, in studies of acute stress, using minimally invasive specimen types like saliva to measure cortisol concentrations is preferred over serum/plasma, partly because saliva collection induces less stress (Vagnoli et al. 2015).
The immune system contains a host of receptors for stress hormones, including cortisol, and psychosocial stress can dampen the immune response, worsen levels of systemic inflammation, and increase susceptibility to disease (Cohen et al. 2019). Development of dried blood spot protocols allows indirect measurement of immune function (e.g., C-reactive protein [CRP], Epstein-Barr virus [EBV]) outside of traditional clinical settings (Cepon-Robins 2021; McDade et al. 2007). Individual experiences with changes in social, economic, or political hierarchies are then linked to variation in these measures (McDade 2002; McDade et al., 2000). In Peru, for example, Tallman (2018), using levels of EBV antibodies, illustrated how adoption of new ideas related to cash-wealth as a marker of success explained reduced immune function among men with lower socioeconomic status. Immune biomarkers can also illustrate the protective value of social institutions. In Bolivia, women with higher levels of emotional and instrumental support had less stress, as measured by lower EBV values (Hicks 2014).
Self-reports of mental health symptoms, including expressions of distress/emotion, on validated scales are also accepted by biocultural practitioners as a measure of stress. For example, Oths (1999) demonstrated how reported symptoms of debilidad (a local idiom related to chronic exhaustion) were associated with a gender imbalance in the household within the context of a stressful agricultural life at high altitude. In a water-insecure Bolivian informal settlement, gender roles, household conflicts, and perceptions of injustice around water insecurity better predicted expressions of anxiety and depression than lack of water alone (Wutich 2020). This approach recognizes that perceptual/cognitive processes around symptom expression are always filtered through both cultural and individual sieves. Accordingly, local adaptation and pretesting are considered standard practices even on otherwise widely validated scales. There are many ways this is achieved, including via ethnographically informed cognitive interviewing or cultural consensus/consonance analysis (e.g., Kaiser et al. 2013; Mendenhall et al. 2016; Snodgrass et al. 2017).
The biomarkers discussed above capture relatively recent (i.e., minutes to months) stress effects. However, innovation in measuring cortisol in hair, nails, and teeth provides information on stress exposure over longer periods. For example, Swales et al. (2018) documented higher hair cortisol associated with both recent and childhood traumatic events among a U.S. sample of pregnant women. Anthropometric measures reflecting delays or stalling of skeletal growth are also often applied as a signal of chronic psychosocial stress over months or years. Central here is the recognition that when psychosocial stressors accumulate or persist, toxic stress can compromise an individual’s ability to rebound from duress, and thus disrupt processes of growth and development (Frongillo et al. 1997; Nelson 2018). For example, among children in Mandeville, Jamaica, the quality of interactions with caregivers predicted individual growth trajectories (height-for-age) above other situational factors like place of residence (natal homes vs. institution) or diet (Nelson 2016).
Epigenetic modification represents a newer measure of stress (Thayer and Non 2015). For example, Congolese mothers’ traumatic experiences while pregnant were associated with DNA methylation in their newborns (Mulligan et al. 2012), and children conscripted in the 1996–2006 war in Nepal exhibited changes in regulatory genes relevant to their phenotypic resistance to viral infections (Kohrt et al. 2016). Another relatively novel stress measurement is epigenetic age (Ryan 2020), based on the recognition that psychosocial stress accelerates cell aging. While currently challenging to interpret, measures of cellular aging, such as telomere length, have the potential to be used to assess the longer-term effects of psychosocial stress on the body not visible through other means (Epel et al. 2004; Marioni et al. 2016; Rentscher et al. 2020; Zahran et al. 2015). For example, racial discrimination, but not other forms of unfair treatment, was associated with shortened telomeres among African Americans in Tallahassee, Florida, suggesting that lifetime exposure to racism may be uniquely stressful (Rej et al. 2020).
Of course, these varied stress markers are not discrete, because the systems they relate to are interconnected. For example, neuroendocrine–hormonal biomarkers reflect acute stress activation of the HPA axis (Ice and James 2007) and can be measured directly—but this activation also increases cardiac output (e.g., blood pressure) (Kaltsas and Chrousos 2007), serotonin, and dopamine, leading to the experience and reporting of depressive symptoms (see Sapolsky 2004). Biocultural studies of stress always assume interrelationships (including feedback loops), unless there is clear evidence to the contrary (though these complex interactions remain incompletely specified).
While interconnected, it is important to recognize that the measures are neither equivalent nor interchangeable. Varied measures can yield disparate findings, such as cortisol concentrations being unassociated with self-perceived psychosocial stress (e.g., Hollenbach et al. 2019; Olstad et al. 2016). Thus, reference categories for comparisons must account for temporal, individual, and population-level variation; universal benchmarks are unlikely to be useful (see, e.g., Hruschka 2021). Instead, each biomarker is considered to reflect just one version of a story about how social context becomes embodied, with its own time scale. For these reasons, studies ideally deploy a range of biomarkers and interpret them relationally as different embodied manifestations of stress. To accomplish this, many scholars have adopted models of allostatic load—defined the cumulative burden of chronic stress across body systems (see Edes and Crews 2017; Guidi et al. 2021).
Some Ethical Considerations
The collection of psychosocial stress outcome biomarkers among living people raises ethical issues. As noted, different specimen types are identified as harmful or not across communities, and stress measurement itself can induce stress. More broadly, the use of political–economic theory, often deployed in designing biocultural research, demands attention to equity, beneficence, and justice in researcher–community relationships, and careful consideration of how findings are communicated and applied (e.g., Leatherman and Goodman 2011; Wutich 2020). Deploying biomarkers in the contexts of situated knowledge elevates those responsibilities because it rests on established trusting and long-term relationships with cultural experts and study communities. Best practices involve transparent data-sharing practices, as determined through consultation with community members. Over the past 15–20 years, the data sovereignty movement has clarified and asserted Indigenous people’s rights to biomarker data collected in their communities. Access and benefit sharing frameworks (Hudson et al. 2020; Robinson 2015) establish terms for storage, accessing, and use that are mutually beneficial to researchers and study communities, and advance community goals in ways that adhere fully to local values. Biocultural researchers are extending the impact of their work through community-engaged, participatory research practices, offering communities benefits beyond near-term solutions to persistent risks. Examples include community education, youth leadership development, or policy advocacy (e.g., Boston et al. 2015; Schell and Tarbell 1998). That said, such ethically necessary practices invariably lengthen the time and complexity of studies, as well as cost. Accordingly, these ethical dimensions need to be planned at the outset of any field-based research.
Footnotes
Acknowledgments
We acknowledge the US National Science Foundation Cultural Anthropology Program grant (Award SBE-2017491) to the NSF Cultural Anthropology Methods Program and Drs. Amber Wutich, Alissa Ruth, and Melissa Beresford for their role in organizing the workshop that generated this collaboration. Any opinions, findings, and conclusions or recommendations expressed in this material are those of the authors and do not necessarily reflect the views of the National Science Foundation.
Declaration of Conflicting Interests
The authors declare no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the National Science Foundation grant number Award SBE-2017491.