
Abstract
From 2006 through 2008, 543 synanthropic Rattus norvegicus were sampled from 20 alleys clustered in five high-density human neighborhoods throughout Baltimore, Maryland, USA. Results indicated that rat coronaviruses (RCV) circulated endemically and infected most rats in the region. Collected animals were tested by serology and RT-PCR for evidence of exposure to RCV and the results were used to characterize the epizootiology of natural infection. Serology showed that 71.8% of rats had IgG antibodies to RCV, indicating previous exposure, while only 3.0% showed evidence of recent infection based on positive PCR results. Incidence was estimated at 16% per month. Antibody and PCR results were uniformly distributed throughout the city. Consistent with historical laboratory studies, RCV appeared to be horizontally transmitted and generated an acute infection with a short period of infectiousness. Evidence of past infection increased with rat age but did not differ significantly between males and females. The low incidence (6/1000) of infectious rats (PCR positive/IgG negative) limits the risk of spillover to nearby species, including humans. These results encourage surveys for other wildlife agents to collect sufficient samples to estimate the power to detect the agents and use serological surveys as a screening tool if active infection is of short duration. Otherwise, substantial efforts may be expended with a low likelihood of detection.
Introduction
Synanthropic rodents in urban centers represent a public health risk because their populations often reach large numbers in close proximity to high densities of humans. These people often lack access to health care, so rodents can transmit various pathogens that cause little or no overt disease in the rats, to the people (Easterbrook et al, 2007, 2008). The diseases present as undifferentiated illnesses in humans, making their identification challenging (e.g., Vinetz, et al, 1996). Norway rats (Rattus norvegicus) are a global threat because of their wide geographic distribution, their exceptional population densities, and the demonstrated diversity of viral and bacterial agents they carry (Easterbrook et al, 2007; Camp et al, 2022; LeDuc and Childs, 2023).
The emergence of SARS-CoV1, MERS-CoV and more recently SARS-CoV2, in humans, are archetypes of the spillover and adaptation to new hosts (humans) of zoonotic agents (Childs, 2004; Flanagan et al, 2012). Rat coronaviruses (RCV) are betacoronaviruses, identified in the 1960s and 1970s (Bhatt, et al, 1972; Innes and Stanton, 1961; Parker et al, 1970) and are related to SARS-CoV-2 (e.g., Woo et al, 2010; Bartak et al, 2021; Islam et al, 2021). Experimental RCV infections in laboratory R. norvegicus were described as acute, self-limited infections that produced little or no clinical disease, except in newborn rats (Parker et al, 1970).
Little is known about RCV epizootiology and early in the COVID-19 pandemic, Bartak and colleagues (2021) reviewed RCVs, commenting on the “… notable lack of interest in rat coronavirus in the context of a potential zoonosis… the high capacity of this family for transient mutations in the hypervariable region of the … S-spike protein … may lead to another crossing of the interspecies barrier…”. When coupled with Li et al’s (2021) report that they “…uncovered a polybasic cleavage site, RARR, in the spike (S) protein of RCoV-GCCDC4, which is dominant in RCoV” in rats, there was renewed interest in characterizing RCVs in wild host populations.
New studies described the molecular heterogeneity of RCVs from a limited number of samples but little new information was generated to characterize the epizootiology of the virus in its hosts (Zhu et al, 2021; Niendorf, et al, 2021; Camp et al, 2022; Fisher et al, 2023). Even the earliest descriptions of RCV provided little detail beyond its existence in wild animals (Parker et al, 1970; Gilmutdinov et al, 2020). The limited evidence of RCV based on genetic data is in striking contrast to earlier serological surveys, such as Parker et al (1970) and Easterbrook et al (2007, 2008) that reported seroprevalence of 40% (N = 108 rats) and 90%, (N = 150 rats) respectively.
While local environmental differences may account for the apparent discrepancy in reported infection levels among the studies, methodologies also need to be considered. Parker (1970) used Complement Fixation assays, and Easterbrook et al (2007, 2008) relied on IgG serology while recent surveys used RT-PCR to detect nucleic acids. RCV IgG persists long after viral clearance (Bhatt and Jacoby, 1977; Jacoby, 1986) while infection is short-lived (Jacoby, 1986). More recent surveys with their smaller sample sizes likely lacked sufficient statistical power to find RCV as they were seeking incident infection in a small pool of samples (Camp et al, 2022 for further discussion). Evaluating the risk of RCV spillover from rats and adaptation to new hosts requires both information on the virus’s genetic structure (Niendorf et al, 2021; Zhu et al, 2021; Fisher et al, 2023) as well as a better understanding how likely actively infectious rats will contact a human. If active infection is uncommon in rats there is a lower likelihood that rat-human contact cross-species transmission events occur, reducing opportunities for adaptation and species jumping. It is not possible to assess the risk without better understanding how RCV is maintained and transmitted within feral Norway rat populations.
To better describe RCV epizootiology we used an archived, large collection of wild rat populations in Baltimore, Maryland, USA, that were obtained to study the role of rodent population structure on pathogens (Glass et al., 2016). Here, we compare serological characteristics and RT-PCR results from the same individuals for RCV and provide descriptions of rat populations that reside near humans. The study characterizes the prevalence and incidence of RCVs in these populations extending the previous century’s studies on Norway rat populations, their pathogens and their impacts on human populations (Emlen, et al, 1949; Davis, 1953; Childs et al., 1987a, 1991).
Materials and Methods
Sampling and processing
Synanthropic rats were collected in central alleys of high-density residential neighborhoods throughout Baltimore. These neighborhoods, historically, (Davis, 1953, Childs et al, 1987a; Glass, et al, 1989; Easterbrook et al, 2005, 2008; LeDuc and Childs, 2023) had large infestations of R. norvegicus that foraged within the alleyways (Glass et al, 1989, 2009). Movements of rats generally covered short distances (Gardner-Santana, et al, 2009; Glass et al, 2016), except mate-seeking forays that could extend several blocks (Glass, et al, 2016).
The survey used a spatially clustered design with groups of four alleys sampled simultaneously for several weeks a year, during two years. Five areas throughout the residential areas of the city were sampled, resulting in 20 alleys surveyed (see Glass et al, 2016). This design was intended to evaluate if there was spatial heterogeneity in infection at different spatial scales. Trapping was performed on consecutive nights in each area until 50 rats were collected from each area during autumn, spring, or winter. Twenty Tomahawk live traps, baited with peanut butter, were placed in the alleys of each block, set near local sunset, and collected the next morning. Trapping was repeated the second year on the same schedule. Rats were transported to the laboratory where they were euthanized according to approved Animal Care and Use Protocols. Upon euthanasia, blood samples were collected by cardiac puncture and organs were sterilely collected, individually, and immediately frozen at −80 C. Standardized data forms, described elsewhere in detail, recorded a unique animal number, date of capture, alley, area collected, sex, and body mass (Glass, et al, 2016).
Laboratory diagnostics
After all wild rat materials were collected they were tested for both IgG antibody, by Multiplexed Fluorometric ImmunoAssayTM (MFIA), and RCV nucleic acids by RT-PCR through the generous assistance of Charles River Laboratories in 2009. The archived samples were collected separately from Easterbrook et al. (Easterbrook et al., 2005, 2007, 2008) and were assayed for anti-RCV IgG antibodies. Samples tested by MFIA followed protocols developed by Charles River Laboratories (Charles River Laboratories, 2018). Each plate included two positive (high, low) and one negative control sample. A plate was considered ‘interpretable’ when at least both the low positive and negative control yielded the correct results. A subsample of specimens was also tested by ELISA using a bivalent A59- and JHM-strain viral lysate antigen. Concordance analysis between the ELISA and MFIA was performed by calculating a Kappa statistic (Landis and Koch, 1977) with standard cutoffs for quality of agreement between assay methods.
RT-PCR was used to identify rats with RCV nucleic acid. They were identified using a protocol developed by Charles River Laboratories. PCR used an ABI 7300 with SuperMix-UDG w/ROX Invitrogen kit #11743500, with SYBR Green. The protocol used a single cycle at 50.0 C for two minutes, followed by a single cycle at 95 C for 10 min, and 45 cycles of 95 C for 15 s and 60.0 C for one minute. Primers amplified all available RCVs except RCV-NJ. The product was 82 bp with the forward primer SDAV1216F (AGAGAGGGACGAAACAAACGG) and the reverse primer SDAV1297R (ATTTCGCTGCACGGCACT), and was identified with a fluorescent probe, SDAV1237P (CTCAGAAAGAAGAATTAG). Standard curves were generated using 5 ul 101 - 107 positive controls in 25 ul reactions. Lung and salivary gland tissues from each wild rat were extracted and run in duplicate.
Statistical analyses
Following laboratory tests for RCV, the field survey data were updated by linking the unique animal numbers in the two files. Basic summary statistics of rats surveyed; numbers by alley and area, sex ratio, and body mass were calculated using EpiInfo version 7.2.5.0 (Dean et al, 2011).
EpiInfo, which relies on frequentist statistical methods, was used for exploratory data analyses of the proportions of IgG antibody-positive and PCR-positive rats stratified by body size class, sex, and area. Maximum likelihood estimates with 95% CI were used to identify effects that were considered in subsequent modeling. Previously identified size classes were used to subdivide rats into juvenile and recently sexually matured (<301 g) (Childs et al; 1987a; Glass et al., 1989), adults (301–400 g), and older adults (>400 g). Growth rates for urban rats (Glass et al, 1988a, 1988b) indicated that these sizes corresponded to rats with average ages of <4 months, 4–7 months, and >7 months of age, respectively.
Based on laboratory studies (see above) PCR positive rats were identified as potentially infectious. Demographic factors influencing local infection rates were modeled with Bayesian inference methods (Walsh et al, 2007). This was undertaken to control for potential confounding effects in observational survey studies caused by variation in rat population structure among areas, primarily size (=age) structure that might differ among areas and alleys, temporal (yearly) heterogeneity, as well as the spatial autocorrelation driven by the adjacency of the neighboring alleys and areas potentially affecting local virus transmission. Size and sex were treated as categorical variables, and nested random effects were included for each alley and area. Fitted probabilities of positive PCR tests for each rat were averaged over each alley and area; the mean and 95% Credible Intervals were calculated from the distribution of these estimates of site PCR positivity rate. Analyses were performed with Markov-Chain Monte Carlo methods, in R, using the rstanarm library, an interface to the Stan software for the current Bayesian inference program (Goodrich et al, 2020).
Results
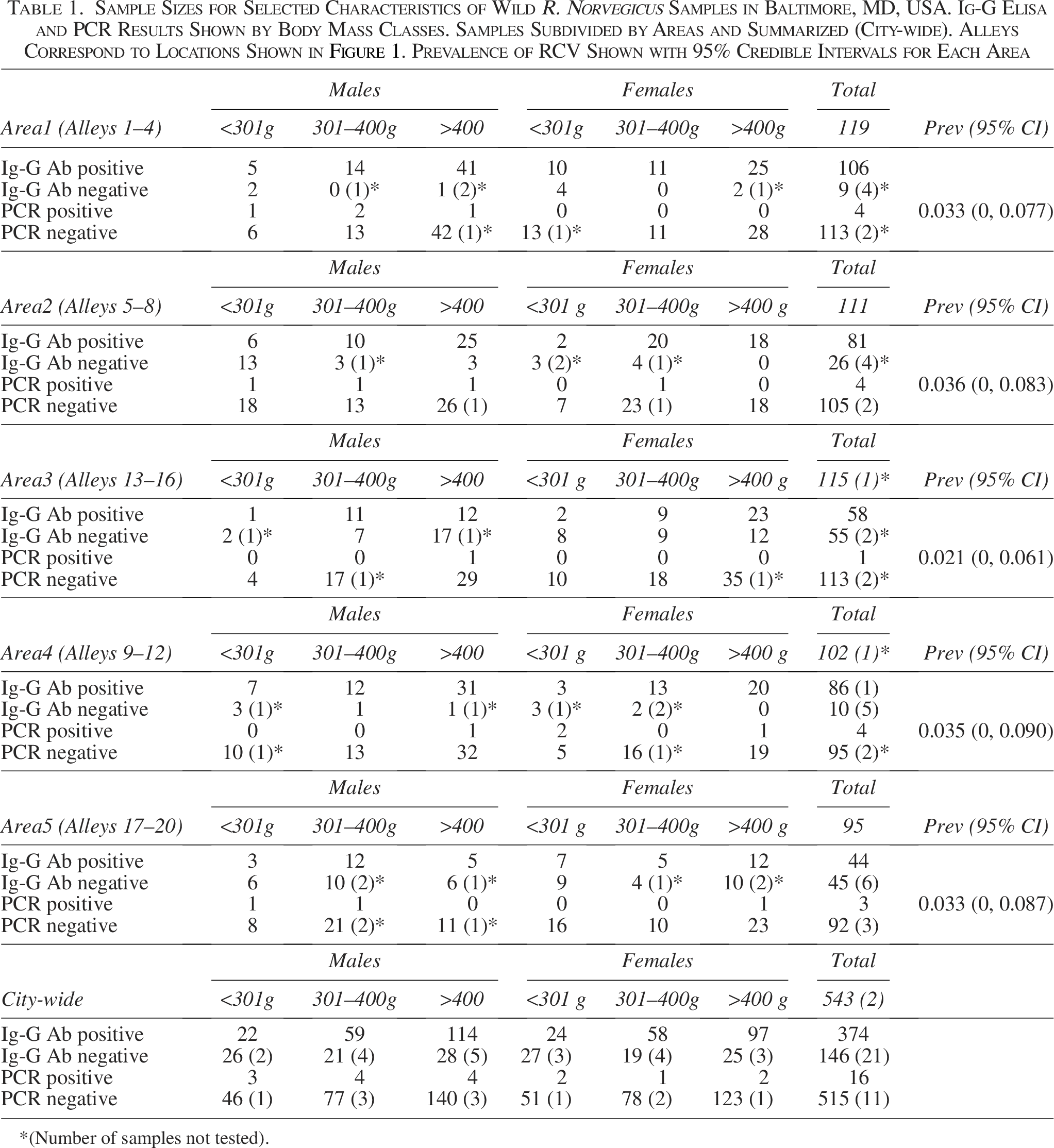
A total of 543 R. norvegicus were collected, euthanized, and tested for RCVs from the 20 alleys clustered in the five areas between November 2006 and May 2008. Traps were set during 27 weeks of the survey and checked nightly. The locations of the areas with the alley ID numbers are reproduced here (Fig. 1). The five areas averaged 17.8 km apart (range 7.04 km-37.6 km). The number of rats collected from the areas ranged from 95 (Alleys 17–20) to 119 (Alleys 1–4) (Table 1). Rats varied in body mass from 40->611 g.

Locations of sampling locations for Rattus norvegicus within Baltimore City. Five areas for sampling are indicated by the clusters of numbered alleys sampled (1–20). Background is the U.S. census tracts, including the surrounding region of Baltimore County and the Chesapeake Bay. Major interstate highways are shown in heavy continuous lines. (Used with permission from Glass et al 2009; PLoS One 2009; 4 (6):e5794. https://doi.org/10.1371/journal.pone.0005794).
S
*(Number of samples not tested).
Based on the frequency of positive IgG ELISA tests, 71.8% (N = 522 tested; 21 samples not tested due to limited sample volumes) were previously infected with RCVs. Validation of the MFIA assays (Charles River Laboratories, 2018) showed near-perfect concordance between the MFIA and the confirmatory test (Kappa = 0.96). Using body mass as a surrogate for the duration of environmental exposure (=age), younger rats (≤300 g) were less likely to have been previously infected (IgG positive), than either of the older (301–400 g, or >400 g) categories of adult rats with IgG antibody rates increasing from 46.4% (95% CI = 36.5, 56.5) to 74.5% (95% CI = 67.6, 81.4) and 79.9% (95% CI = 75.1, 84.8). There was no statistical difference in antibody prevalence between males and females when stratified by size class (OR = 1.025; 95% CI = [0.678, 1.550]).
Sixteen rats were RT-PCR positive (3.0%; N = 532; 11 samples not tested due to limited material). Positive samples tended to be more common among younger adults (301–400 g), although the difference was not statistically significant (OR = 2.793; 95% CI = [0.813–12.200]). Further stratifying within the youngest age group (<301 g) suggested that infection first appeared when rats were two to three months old. There were no PCR positive rats among the limited sample of the youngest group (<100 g; n = 17), with the first positive samples found in rats 101–200 g (2.8%; n = 35) and the proportion peaked in the recently matured animals (201–300 g; 7.8%; n = 51). The proportion of PCR-positive rats declined among the oldest rats (2.5%; n = 437). Males tended to have RCV nucleic acid more frequently than did females (4.0% vs 1.5%; N’s = 274 and 258, respectively), but the difference was not statistically significant (OR = 2.867; 95% CI = [0.854, 8.545]). Among 16 PCR-positive animals, most (86.7%) also had measurable IgG antibodies. The three PCR-positive/IgG negative animals were presumably infected within the week prior to capture and were infectious (Jacoby, 1986). We used these three individuals with the 146 IgG-negative (=susceptible) rats to estimate the incidence of new RCV infections within the rat populations as 16.2%/month.
Modeling RCV infection dynamics, using the PCR positives, within the study area, with Bayesian estimation methods supported the interpretation of the exploratory, frequentist, results. The model parameter posteriors for positive PCR prevalence for area or year effects overlapped zero, showing little evidence for geographic or temporal heterogeneity within the study area (Walsh et al., 2007). There was no evidence for annual differences in PCR positive animals, by area, based on broadly overlapping 95% credible intervals, when aggregated by year of capture. Models including conditional autoregression for the random effects did not fully converge and yielded a posterior distribution for the autoregression parameter similar to the prior, suggesting minimal spatial autocorrelation. Rat sex also did not contribute to the final model. The estimated prevalence of PCR-positive rats varied little among areas (0.021–0.035), and the differences were not statistically different from one another, as characterized by broadly overlapping 95% credible intervals (Table 1).
Two final models were considered; both included age, which either used the original three ages (in this model mass classes contributed most substantially to variation in observed prevalence among areas). The alternative model further subdivided <301 g mass classes. This model collapsed to rats ≥101 g and those <101 g, since no differences were seen in the parameter estimates among the larger mass classes. The random effects of alleys and areas did not contribute to variability in the prevalence of infection. This is consistent with the conclusion that any differences in prevalence between sites were explained by differences in the age distributions of the sampled rats.
Discussion
RCVs, despite being identified more than 60 years ago (Bhatt, et al, 1972; Innes and Stanton, 1961; Parker et al, 1970), received little attention except among veterinary scientists (Bhatt and Jacoby, 1977; Jacoby, 1986) and neurovirologists (e.g., Schwender et al, 1991; Cowley and Weiss, 2010). In laboratory studies, RCVs typically caused acute infection with little or relatively minor disease in host species, except for neonates (Parker et al, 1970; Jacoby, 1986). The adaptive immune response, included cellular (notably CD-8 T cells) and humoral immunity, marked by circulating IgG, that developed within seven days, peaked about day 14, and remained elevated until at least day 30 (Jacoby, 1986). In studies, 2–2.5 month old rats were experimentally infected and sacrificed daily. Virus was recoverable from nasopharyngeal washes and lymph nodes by day 2 and from lungs by day 3. Virus could not be recovered once IgG was measurable on days 7–8 (Bhatt and Jacoby, 1977). Despite this, IgG antibody remained detectable until experiments ended at 30 days (Jacoby, 1986). T-cell immunity, alone, was sufficient to minimize host disease impacts while IgG served as a marker of recovery from infection but was not required for clearance (Jacoby, 1986; Cowley and Weiss, 2010). Animals were resistant to reinfection with the same strain of RCV (Bhatt and Jacoby, 1977; Jacoby, 1986). Vertical transmission from infected dams to their offspring was not observed, with virus maintained by horizontal transmission among colony members presumably by contaminated bedding (Jacoby, 1986; Bartak, et al, 2021).
Few studies of RCVs in wild rats (Rattus spp) were reported until the emergence of SARS-CoV1, and MERS-CoV (Woo et al, 2009). As the dynamics of host jumping (Childs, 2004; Flanagan et al, 2011) became better appreciated, interest in RCVs reawakened. This was driven both by the genetic relationship of RCVs and SARS-CoV1 (Woo et al, 2009; Bartak et al, 2021; Islam et al, 2021) and by the widespread physical proximity of R. norvegicus and human populations (LeDuc and Childs, 2023). This raised concern that RCVs posed an unassessed risk to people (Bartak et al, 2021).
Subsequent field surveys were short-term yielding small sample sizes (Zhu et al, 2021; Niendorf, et al, 2021; Camp et al, 2022; Fisher et al, 2023) that produced limited documentation of RCV in the rats (see Li et al, 2021 for a counter-example). However, reports of infected rats with variants of RCVs (Zhu et al, 2021; Li et al, 2021) reinforced speculation of the risk for RCVs to jump to humans (Bartak et al, 2021). There was little evidence to document rat-to-human spillover of RCV, and despite genetic sequences, few details outlining the epizootiology of RCVs in wild reservoir populations as an antecedent condition for species jumping (Childs, 2004). Conversely, other studies reported possible spillover of SARS-CoV1 from humans into urban Norway rat populations, raising concerns that new environmental reservoirs of SARS might develop (Fisher et al, 2023).
Our study was intended, in the absence of documented RCV spillover, to better describe the epizootiology of these viruses, and consequently, why identifying the natural host(s) of SARS-like viruses has been challenging (Niendorf et al, 2021; Camp et al, 2022; Fisher et al, 2023). It also points to the need to more explicitly define field sampling strategies and recognize the within-host responses to infection when designing surveys. The recent surveys usually obtained sample sizes <50 animals/site and used PCR to detect virus. If only 1–3% of rats are PCR positive (Table 1), 50 samples lack sufficient statistical power to detect infection in even one rat, with a likelihood of 80%. At best, RCV infection would appear sporadic. If laboratory studies are correct and RCV is horizontally transmitted, then targeting older animals will be more successful for sequence recovery. We recommend, as a general rule, that future surveys for wildlife pathogens report power calculations for the chance of detection rather than reporting virus as ‘absent’ when sample sizes are insufficient to detect a threshold of infection. A properly curated, sufficiently sampled collection better ensures that interpretable results are generated, as well as reducing detection time (Cook, et al, 2020). Similar conclusions were discussed in more detail by Camp et al (2022). This study confirms that RCV’s have circulated undetected in urban U.S. areas for at least 20 years. The distances among areas (Fig. 1) far exceeded the observed dispersal rates of rats in this area (Glass et al, 1989, 2016) suggesting the absence of an ‘area effect’ in the Bayesian models is consistent with a longer-term, endemic persistence.
Our results support early laboratory characterizations that RCVs cause acute infections in rats and RCV are horizontally transmitted, likely from a common environmental source. Both exploratory analyses and the Bayesian models indicate that the youngest rats are less likely to show evidence of current or past infection. The Bayesian model with two size classes (<101 g or greater) produced a parsimonious result, with no PCR positive animals in the youngest age class. Overall, the PCR data indicated that few rats were recently infected (3.0% of more than 500 individuals; Table 1) and even fewer were presumptively actively infectious (0.6%; N = 3 IgG negative/PCR positive animals). Despite these low levels, there was widespread evidence of endemic transmission (71.8% of the sample were IgG positive). The estimated monthly incidence was comparable to reports by Childs et al. (1987b) for hantavirus in rats from Baltimore (12.1%/month).
Our survey did not find PCR-positive samples among the smallest animals (≤100 g), however, those sample sizes lacked statistical power (Table 1). Examination of the data showed recent infection seemed to peak among recently mature rats (201–300 g) (Glass et al, 1989), the explicit modeling did not require further distinction of rat age. Antibody prevalence continued to increase as rats aged (Table 1), implicating an ongoing hazard of environmental transmission among susceptible rats. Taken together, these results support laboratory studies that RCVs were horizontally transmitted (Jacoby, 1986; Bartak et al, 2021), presumably because of either protective maternal antibody from previously infected dams or lethal effects on neonates born to uninfected dams (Parker et al, 1970).
One aspect of infection dynamics may bear future examination. There was not a statistically significant difference in IgG antibody prevalence between sexes (IgG antibody prevalence 72.2%: 71.6%, male: female; Table 1), but there was a trend for males to be more frequently PCR positive than females (4.0% vs 1.5%; Table 1). Why infection prevalence (IgG antibody) should be so similar while incidence (PCR positives) should appear elevated among males may be a sampling artifact. Alternatively, there may be sex differences in the effectiveness of viral clearance that impact the sex-specific rates of infectiousness (Klein and Flanagan, 2016). The limited number of PCR-positive animals precludes a more definitive interpretation. In the field, even larger sample sizes would be needed to resolve the question. Instead, laboratory studies may be the more appropriate approach to resolve the alternatives.
Results demonstrate the value of including serological tests, as screening tools, in surveys for previously unrecognized infectious agents. It suggests the discrepancy between previous serosurveys and more recent surveys for virus is more methodological and apparent than demonstrating geographic variation in transmission dynamics. Despite the short-term nature of infection, RCV seems to have persisted endemically within these wild rat populations. Evidence was found in all neighborhoods tested from 2006 through 2008, with no year-to-year variation, and PCR-positive rats were found in all areas of residential neighborhoods within the city at comparable levels (Table 1).
The low rate of active infection in the host rat populations suggests that monitoring nearby human populations to detect spillover is unlikely to demonstrate species jumping. Seoul (SEOV) virus PCR positive tests in these rats (data not shown) was 50% (N = 542; 1 missing), while surveys to detect spillover of SEOV into people from these areas showed very low rates of cross-species transmission (Childs et al, 1991). Confirmed SEOV IgG-positive humans, with no travel history outside the city, were less than 2/1000 (Glass, et al, 1990; Childs et al, 1991). The even lower level of active RCV infection within rats suggests substantially fewer rats are available to transmit RCV to people, even though the modes of transmission of RCV and SEOV within the host species seem similar.
Conclusion
RCV transmission and opportunities to adapt to humans seem unlikely in these modern urban settings. Adaptation requires many steps, but even before genetic variants are selected, the repeated contacts that bring the viruses from the original host into proximity with the new (human) host seem to occur rarely, if at all (Childs, 2004). As such, the short duration of RCV infection in individual wild rats serves as an initial, within-host, barrier that makes these events unlikely.
Authors’ Contributions
Field sample collection (G.G.), Laboratory sample collection (G.G. and S.K.), Laboratory testing and data management (S.K.), Data analyses (G.G. and A.W.). Article preparation (G.G., A.W. and S.K.).
Footnotes
Acknowledgments
The authors thank the many staff and students who, over the years, helped in the field and laboratory collecting and testing materials. Special thanks to the Charles River Laboratories for diagnostic assistance with this study. We also wish to thank Diane E. Griffin, the chair of the W. Harry Feinstone Department of Molecular Microbiology & Immunology (1994–2014) for supporting the inquiries outlined here and for her support over the decades to better understand how viruses cause diseases and impact the health of people. We also acknowledge the two reviewers for insightful comments to improve earlier drafts of the article.
Author Disclosure Statement
The authors do not report a conflict of interest.
Funding Information
This project was supported by NSF Grant # EF0525751. The Johns Hopkins Animal Care and Use Committee (protocol no. RA05H6) approved all procedures described in this study.