
Abstract
Background:
Usutu virus (USUV) is an emerging arbovirus transmitted predominantly by mosquitoes of the genus Culex, although several other genera, including Aedes, Culiseta, Mansonia, Anopheles, and Coquillettidia, have also been implicated as potential vectors. The virus was originally found in sub-Saharan Africa but is increasingly detected in Europe. While human infections are often asymptomatic, USUV has been linked to neuroinvasive disease. In temperate regions, where mosquito activity is seasonal, mechanisms enabling virus persistence are poorly understood. Vertical transmission has been proposed as a route for interepidemic maintenance, but experimental evidence remains limited.
Objective:
To determine whether USUV can be vertically transmitted between mosquito generations and assess the effect of temperature on the vertical transmission rate in Cx. pipiens bioform molestus.
Methods:
Laboratory-reared Cx. pipiens bioform molestus were provided with a bloodmeal infected with USUV and maintained at either 27°C or 23.5°C. Infected females (F0) were allowed to oviposit, and two successive generations (F1 and F2) were reared. Individuals from all life stages were screened for USUV RNA using RT-qPCR. Infection prevalence and relative viral RNA quantification were compared between temperature conditions and generations.
Results:
At 27°C, the vertical transmission rate was high, with 94% (n = 30) of F1 and 100% (n = 11) of F2 adult mosquitoes testing positive, although USUV RNA was also detected in immature stages (eggs, larvae, and pupae). At 23.5°C, only one F1 adult mosquito (1.25%) tested positive, with no infection detected in larvae or other immature stages and no F2 infection was detected. Temperature had a significant effect on vertical transmission rate (p < 0.001), while differences between generations were not statistically significant.
Conclusion:
These findings demonstrate that vertical transmission of USUV in Cx. pipiens bioform molestus occurs at significantly higher rates at 27°C (94–100%) compared to 23.5°C (1.25%), suggesting temperature-dependent mechanisms specifically affecting vertical inheritance. While temperature effects on viral acquisition and dissemination are well-established, our experimental design specifically tests vertical transmission by examininginfection in F1 and F2 offspring that had no direct exposure to the virus, allowing us to distinguish vertical inheritance from horizontal transmissionprocesses.
Graphical abstract
Keywords
Introduction
Usutu virus (USUV) is an emerging mosquito-borne flavivirus and an increasing public and animal health concern in Europe and sub-Saharan Africa (Ashraf et al., 2015; Clé et al., 2019). First isolated in South Africa in 1959, USUV circulates primarily in a bird–mosquito enzootic cycle involving mosquitoes of the genus Culex, particularly Culex pipiens, and avian reservoir hosts; other genera such as Aedes, Culiseta, Mansonia, Anopheles, and Coquillettidia have also been implicated as potential vectors (Mancini et al., 2017; Williams et al., 1964). Mosquitoes undergo a holometabolous life cycle consisting of four distinct stages: egg, larva, pupa, and adult. The aquatic immature stages (egg, larva, and pupa) precede the emergence of the terrestrial adult, which is responsible for host-seeking, blood-feeding (in most cases), and pathogen transmission (John Ravindran and Eapen, 2024). While human infections are often asymptomatic, cases of neuroinvasive disease have been reported, highlighting the virus’s zoonotic potential and underscoring the need to understand its mechanisms of persistence and transmission (Graninger et al., 2022; Vilibić-Čavlek et al., 2025). Recent reports further confirm sporadic human neuroinvasive cases in Austria, Italy, and Croatia, reinforcing its emerging public health relevance (Angeloni et al., 2023; Vilibić-Čavlek et al., 2025). Following its detection in Italy in 1996, retrospective and prospective studies have demonstrated widespread circulation across Europe, with subsequent increases in detection likely reflecting both geographic spread and improved surveillance. The virus has since expanded northward, causing high mortality in birds, particularly the Eurasian blackbird (Turdus merula) (Ashraf et al., 2015; Holicki et al., 2020; Rijks et al., 2016; Van Irsel et al., 2025; Weissenböck et al., 2010).
In temperate regions, where seasonal conditions restrict year-round mosquito activity and reduce vector–host contact, a central and unresolved question is how USUV persists during winter or interepidemic periods. The mechanisms underlying USUV overwintering and persistence remain poorly understood. Vertical transmission in mosquitoes can occur via two primary mechanisms: transovarial transmission, in which viruses infect developing oocytes prior to oviposition, and transovum transmission, where eggs become contaminated with virus during or after oviposition (Darby et al., 2023; Schilling et al., 2025). Through vertical transmission, infected female mosquitoes can pass the virus directly to their offspring. This route has been described for other flaviviruses, including West Nile virus (WNV) and Zika virus (ZIKV) (Anderson et al., 2020,2012; Ciota et al., 2017; Nag et al., 2021), and may similarly enable USUV to persist during environmentally unfavorable periods and bridge seasonal gaps in transmission. Evidence for vertical transmission of USUV remains limited. A recent study by Schilling et al. (2025) detected USUV RNA in wild-caught Culex mosquitoes, suggesting, but not confirming, vertical transmission in natural populations. However, other mechanisms may also contribute to virus maintenance, including the reintroduction of the virus via migratory birds and the possibility of chronic infection in avian hosts, leading to repeated episodes of viremia that sustain enzootic circulation (Kuchinsky, 2022). To date, no experimental studies have demonstrated whether USUV can be vertically transmitted across generations, nor identified the environmental conditions under which this occurs.
Temperature is a key environmental determinant of arbovirus transmission, influencing viral replication, dissemination, and vector competence across mosquito life stages. For vertically transmitted viruses, temperature can also affect infection of developing oocytes, viral load in progeny, and persistence across generations (Agarwal et al., 2017; Bellone and Failloux, 2020; Lequime et al., 2016). However, despite its well-established role in horizontal transmission, the influence of temperature on USUV vertical transmission remains unexplored.
Culex pipiens sensu lato is a principal vector of USUV in Europe and comprises two ecologically distinct bioforms: pipiens and molestus. The pipiens bioform is primarily ornithophilic and requires a blood meal for reproduction, whereas the molestus bioform is autogenous, anthropophilic, and capable of reproducing without a blood meal. Additionally, Cx. pipiens bioform molestus is well adapted to urban and subterranean environments, where it may remain active year-round under favorable conditions. These ecological and physiological differences suggest that the molestus bioform may play a distinct role in virus maintenance, particularly in urban settings.
To address this gap, we experimentally tested two interlinked hypotheses: (1) that USUV can be vertically transmitted across generations in Culex pipiens bioform molestus, and (2) that the vertical transmission rate increases with temperature, being higher under peak summer-like conditions (27°C; late June–mid July) compared to late summer/early autumn conditions (23.5°C; late August–September). These ecological and physiological traits make this bioform particularly relevant for investigating vertical transmission and potential mechanisms of viral persistence in urban environments.
Understanding whether and how USUV can persist through vertical transmission is critical for predicting seasonal re-emergence, informing vector control strategies, and assessing the long-term establishment of the virus in temperate regions. As climate change alters temperature profiles and extends mosquito breeding seasons, identifying the ecological mechanisms that enable arbovirus persistence is increasingly important for public health preparedness across Europe and beyond.
Materials and Methods
Mosquito rearing
Eggs of Cx. pipiens bioform molestus were originally collected from field-collected mosquitoes in the Netherlands (Vogels et al., 2016). Eggs were placed in larval trays containing 2 L of distilled water and provided daily with finely ground fish food (SUPERVIT 8MIX, Tropical, Chorzów, Poland). Larval development was monitored daily, progressing through four instars (L1–L4). Upon pupation, pupae were transferred to 100 mL beakers with distilled water and placed inside rearing cages equipped with 10% sucrose solution and oviposition cups to support adult emergence and reproduction.
Rearing was conducted in a controlled-climate chamber (HPP410eco, Memmert GmbH, Germany) maintained at either 27.0°C ± 0.1°C or 23.5°C ± 0.1°C, with a 12:12 h light:dark photoperiod and 70% (±0.5) relative humidity (Graphical Abstract).
Following oviposition, F0 females were maintained under their respective temperature conditions and monitored throughout the experimental period. A subset of females was collected and processed for RT-qPCR analysis to confirm infection status, while the remaining individuals were discarded according to standard laboratory protocols.
The mosquito colony used in this study was not screened for insect-specific flaviviruses or endosymbionts such as Wolbachia. However, the colony has been maintained under controlled laboratory conditions for multiple generations, minimizing the likelihood of uncontrolled environmental contamination or unintended infections.
USUV stock and titration
USUV stock (USUV, Africa 3 lineage; sublineage Africa 3.1; GenBank: KY128482.1) (Snoeck et al., 2022) was obtained from Erasmus MC (Rotterdam, the Netherlands) through the European Virus Archive Global, isolate S. nebulosa-6004/NL/2016. Passage 4 of the virus was used for all assays. Plaque assay was performed using Vero B4 cells (African green monkey kidney, ATCC® CCL-81™ derivative) to determine viral titer (1.2 × 106 Plaque-forming units per milliliter [PFU/mL]). Vero B4 cells were cultured in Dulbecco’s Modified Eagle Medium (DMEM; Gibco, Thermo Fisher Scientific, USA) supplemented with 10% heat-inactivated fetal bovine serum (FBS; Sigma-Aldrich, USA) and 1% penicillin–streptomycin (Gibco, Thermo Fisher Scientific, USA). Cultures were maintained at 37°C in a 5% CO2 humidified incubator.
Plaque assay protocol
Vero B4 cells (1.5 × 105 cells/well) were seeded into 24-well tissue culture plates (Corning, USA) and incubated overnight to form a confluent monolayer. The following day, cells were washed twice with sterile phosphate-buffered saline (PBS; pH 7.4, Gibco) and 200 µL of 10-fold serial virus dilutions (in serum-free DMEM) were added per well in duplicate. Plates were incubated at room temperature for 1 h with gentle rocking every 15 min to facilitate adsorption. After adsorption, the inoculum was removed and replaced with an overlay composed of a 1:1 mixture of 2× DMEM supplemented with 4% FBS and 2.4% Avicel® RC-581 (FMC BioPolymer, Philadelphia, USA), resulting in a final concentration of 1 × medium with 1.2% Avicel. Plates were incubated at 37°C for approximately 4 days.
Fixation, staining, and titer calculation
Infected cells were fixed with 1 mL of 4% paraformaldehyde (PFA; Sigma-Aldrich) per well and incubated at 4°C overnight. Following fixation, plates were rinsed several times with distilled water and stained with 1% (w/v) crystal violet solution (Sigma-Aldrich) in 20% methanol for 1 h at room temperature. Excess stain was removed, plates were washed thoroughly with distilled water, and air-dried overnight in the dark. Viral titers were determined by counting plaques in wells with fewer than 20 plaques. Titers (PFU/mL were calculated according to the following formula:
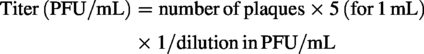
RNA quantification and standard curve
Usutu viral RNA was quantified via spectrophotometry using the DeNovix DS-11 FX + system (DeNovix, Delaware, USA), yielding a concentration of 23.622 ng/µL. A tenfold serial dilution of the virus was prepared, and corresponding Ct values were obtained via USUV-specific RT-qPCR. These values were used to generate a standard curve correlating Ct values to USUV relative viral RNA quantification (Fig. 1), which enabled estimation of USUV viral load in mosquito-derived samples.
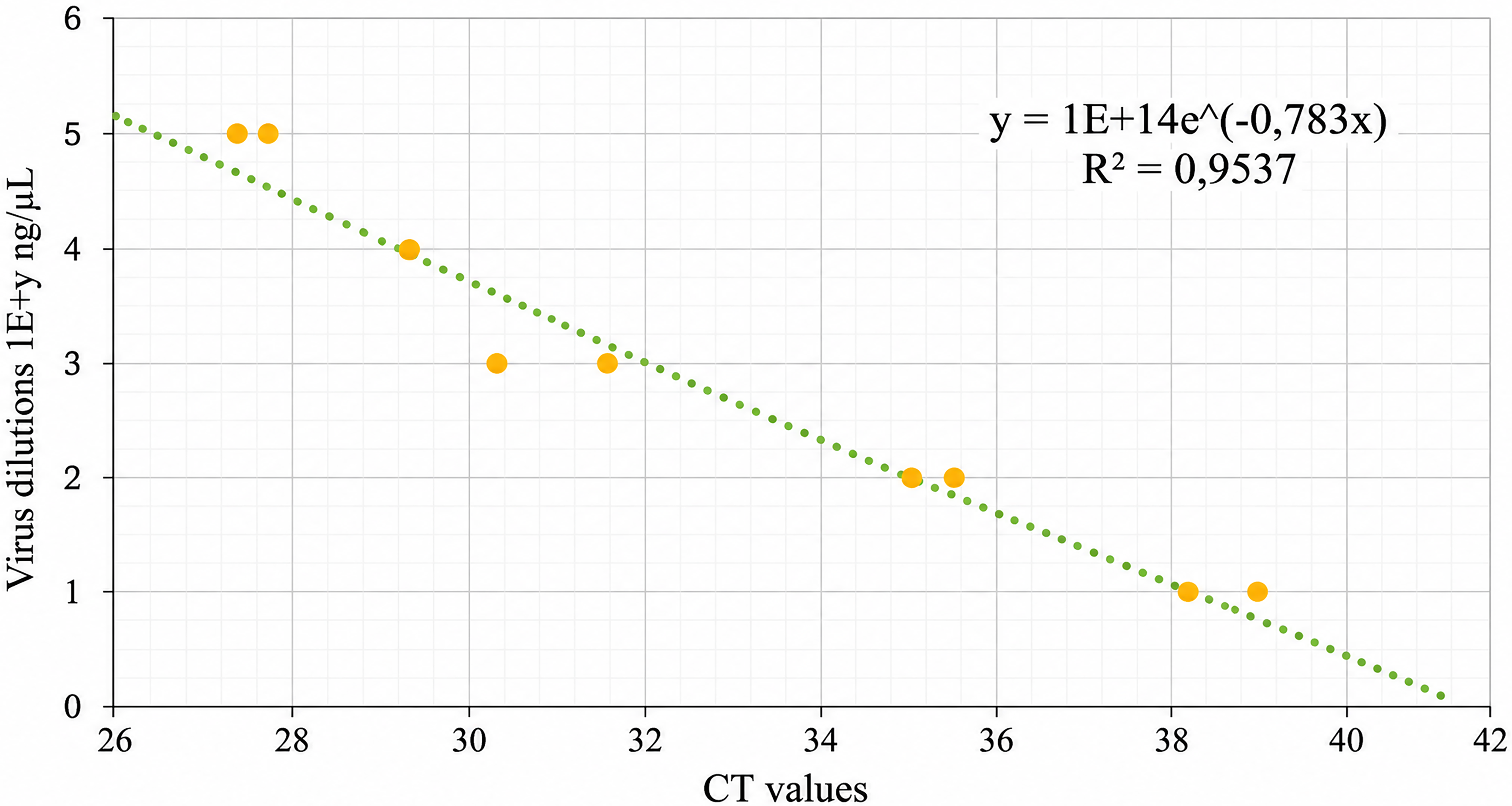
The standard curve of USUV titer values in nanograms per microliter and RT-qPCR Cycle threshold (Ct) values. USUV was serially diluted, and the Ct values were recorded on a QuantStudio 5 System (Applied Biosystems). Standard curve correlating virus concentration with Ct values. The regression yielded an R2 of 0.9537, indicating a strong linear relationship. Ct values were converted to concentrations using the regression equation, with a cutoff Ct of 40 corresponding to 5.9 × 10−4 ng/μL. USUV, Usutu virus.
Vertical transmission experiment
Adult Cx. pipiens bioform molestus females from the parental (F0) cohort were exposed to USUV-infected blood meals under two constant temperature conditions (27°C and 23.5°C) (Graphical Abstract). To monitor initial infection and viral uptake, a small subset of engorged females (n = 5 per temperature condition) was immediately processed as day-1 post-infection controls. These day-1 controls provide baseline viral RNA measurements to confirm successful infection from the blood meal. No additional “day-0” controls were required, as negative controls consisted of unfed females and females exposed to non-infectious blood meals. The remaining engorged females were maintained for oviposition to generate the F1 progeny, which were reared through successive life stages and gave rise to the F2 generation. Counts of individuals at each developmental stage, as well as pools screened for infection, are summarized to illustrate survival and progression under the two temperature regimes (Supplementary Table S1).
Parental (F0) cohort and blood feeding
Adult females were allowed to mature for 7 days post-emergence and starved for 24 h before feeding. They were offered a USUV-infected blood meal consisting of a 1:1 mixture of anonymized human blood (Norrlands University Hospital blood bank, Umeå, Sweden) and USUV stock (final concentration: 1.2 × 106 PFU/mL). A membrane feeding system (Hemotek Ltd., United Kingdom) with Parafilm covering was used to deliver 750 µL per feeder, maintained at 37°C and placed overnight in a dark environment to promote feeding.
Postfeeding handling and rearing
Following feeding, mosquitoes were anesthetized at +4°C, and fully engorged females were transferred to secure rearing cages (BugDorm, MegaView Science Co., Ltd., Taiwan). Nonengorged individuals were euthanized. Two experimental groups were established: Group A (27°C) and Group B (23.5°C). In each cage, 10 crystallizing dishes containing 50 mL distilled water were provided to facilitate oviposition. Blood-fed females were maintained under standard rearing conditions with 10% sucrose solution ad libitum.
F1 generation rearing and sampling
For clarity, mosquitoes sampled under the two temperature treatments are hereafter referred to as Group A (27°C) and Group B (23.5°C) throughout this analysis. Fifty-nine egg rafts including 29 from the 27°C and 30 from 23.5°C of fully engorged female mosquitoes exposed to an USUV-infected blood meal (both Group A and B) were collected. A subset of egg rafts was sampled and the remainder was transferred to larval trays (approx. 30 × 45 × 12 cm) containing 2 L of distilled water and fed progressively increasing amounts of finely ground fish food (SUPERVIT 8MIX, Tropical, Chorzów, Poland) to support larval development. Larvae were sampled at each instar (L1–L4), as well as at the pupal stage (from Groups A and B) systematically. The remaining four egg rafts were stored at –80°C for downstream viral analysis.
Emerging pupae from the reared trays were transferred daily to new BugDorm cages (32.5 × 32.5 × 32.5 cm), where adults were allowed to emerge and provided with 10% sucrose solution ad libitum. Seven days after first emergence, an oviposition dish (50 mL distilled water) was introduced to initiate a second generation (F2) rearing. As with F1, egg rafts, larval stages, pupae, and adults were systematically sampled and frozen at –80°C.
Sample homogenization
For each generation and temperature group, we sampled at least 10 individuals per life stage (L1–L4, pupae, adults), except for life stages in the
Detection and quantification of USUV RNA
Total RNA was extracted from individual mosquito samples using the QIAmp Viral RNA Mini Kit (QIAGEN, Hilden, Germany) following the manufacturer’s protocol. Extracted RNA was either used immediately or stored at –80°C prior to RT-qPCR analysis. Quantification of USUV RNA was performed using the QuantStudio™ 5 Real-Time PCR System and QuantStudio™ Design and Analysis software v1.5.2 (Thermo Fisher Scientific, USA). Reactions were conducted using the qPCRBIO 1-Step Go Lo-ROX kit (PCR Biosystems Ltd., United Kingdom) according to the manufacturer’s instructions, in a final reaction volume of 20 µL. Each reaction contained 1 µL of extracted RNA, 5 µL of 2× Lo-ROX master mix, and 0.8 µM of each primer, along with 0.4 µM of probe (or specify actual concentrations if different). The USUV-specific primers and probe were synthesized by Eurofins Genomics (Germany) and used as follows: Forward:(5′→3′) CAAAGCTGGACAGACATCCCTTA (Eurofins Genomics, Germany), USUV-Reverse: (5′→3′) CGTAGATGTTTTCAGCCCACGTT (Eurofins Genomics), and USUV-Probe: (5′→3′) [FAM]GACATATGGTGTGGAAGCCTGAT[BHQ1] (Eurofins Genomics) as described by (Nikolay et al., 2014).
Thermal cycling conditions were: reverse transcription at 45°C for 10 min, initial polymerase activation at 95°C for 2 min, followed by 40 amplification cycles of denaturation at 95°C for 5 s, and annealing/extension at 60°C for 25 s. All reactions were run in technical duplicates. A standard curve was generated from a 10-fold serial dilution of quantified USUV RNA (see Methods: Standard Curve Generation), Ct values were converted to relative RNA quantities using a standard curve and are therefore semiquantitative estimates of viral load. Each RT-qPCR plate included a no-template control consisting of nuclease-free water, alongside a positive control containing RNA from the original USUV stock, to monitor assay consistency and ensure that detected viral RNA reflected true infection rather than contamination or experimental artefacts. Mosquitoes with Ct values below the detection threshold (typically <40) were considered positive for USUV RNA. RNA concentrations were compared by life stage and temperature group for downstream analysis.
Data analysis
Viral RNA concentrations for each mosquito sample were estimated by interpolating Ct values from the standard curve generated using serial dilutions of quantified USUV RNA (see Methods: Standard Curve). For statistical analyses, vertical transmission rate was defined as the proportion of offspring testing positive for USUV RNA (Ct < 40) at each life stage and generation. Differences in vertical transmission rates (i.e., proportion of positive individuals) between generations (F1 vs. F2) or temperature groups (27°C vs. 23.5°C) were tested using Fisher’s Exact Test, appropriate for categorical binary outcomes. Differences in Ct values across generations and life stages were assessed using the Kruskal–Wallis rank-sum test, due to the nonnormal distribution of Ct values. When significant, post hoc pairwise comparisons were performed using Dunn’s test with Bonferroni adjustments. Statistical significance was set at α ≤ 0.05 for all tests. To evaluate the magnitude of temperature effects on vertical transmission rates, effect sizes were calculated using Cohen’s h for differences in proportions. A post-hoc power analysis was conducted to confirm the statistical robustness of the observed transmission differences given the experimental sample sizes. For nonparametric comparisons of viral load (Ct values) across generations, the epsilon-squared (ε2) effect size was computed to determine the proportion of variance attributed to the generational cohort. All analyses were conducted in R version 4.3.3, using the rstatix package for nonparametric tests and multiple comparison procedures.
Results
F0 cohort sampling and feeding
At 27°C, a total of 400 adult mosquitoes (males and females) were collected at the time of adult emergence for experimental allocation. Of these, 200 females were offered an infectious blood meal, resulting in 95 engorged individuals. Five engorged females were processed at day 1 postinfection as controls, while the remaining 90 were maintained for oviposition. An additional five unfed females, together with females exposed to a non-infectious blood meal, were included as negative controls.
At 23.5°C, 350 adult mosquitoes were similarly collected at emergence, with 200 females exposed to an infectious blood meal. A total of 100 females successfully engorged; five were processed at day 1 post-infection as controls, and the remaining 95 were maintained for oviposition. A subset of five engorged females per temperature condition was processed as day-1 post-infection controls to confirm initial virus uptake. These controls were not included in subsequent oviposition or vertical transmission analyses. Negative controls, including unfed females and those exposed to non-infectious blood meals, remained negative throughout the study, confirming assay specificity.
First filial (F1) generation
At 27°C, oviposition from the F0 cohort resulted in 29 egg rafts, yielding 130 first instar (L1) larvae. Developmental progression led to 67 fourth instar (L4) larvae, of which 5 were sampled for RT-qPCR analysis, leaving 62 individuals that progressed to pupation. From these, 62 pupae were obtained, with 5 sampled, resulting in 57 adults emerging (25 females and 32 males).
At 23.5°C, 30 egg rafts produced 150 L1 larvae. Developmental progression resulted in 70 L4 larvae and 40 pupae, of which 5 were sampled for RT-qPCR, leaving 35 individuals. From these, 35 adults emerged, comprising 28 females and 7 males.
All developmental counts presented here have been verified to ensure consistency with sampling procedures and corresponding tables.
Second filial (F2) generation
At 27°C, F2 production was limited. Three egg rafts were deposited, yielding 20 L1 larvae, with survival decreasing sharply through development to only 6 pupae. This cohort produced 2 adult females and 3 adult males.
At 23.5°C, F2 production was more robust. A total of 12 egg rafts were deposited, producing 60 L1 larvae and progressing to 30 L4 larvae and 28 pupae. Adult emergence yielded 22 females and 6 males.
Screening for USUV RNA was conducted across both temperature regimes in all collected samples, including male and female mosquitoes.
Comparative trends across temperatures
Although both temperatures supported multigenerational development, mosquito productivity was higher at 23.5°C relative to 27°C, which was particularly evident in the F2 generation, where survival and adult emergence were markedly greater at the lower temperature.
Vertical transmission rate by temperature and generation
At 27°C, vertical transmission of USUV RNA was detected in both F1 and F2 generations. Across all developmental stages, the mean infection rate was 94% (95% CI: 79.19–99.23; n = 30) in the F1 generation and 100% (95% CI: 71.51–100.00; n = 11) in the F2 generation (Table 1). In contrast, at 23.5°C, vertical transmission was minimal. Only 1.25% of F1 individuals tested positive (95% CI: 0.22–6.75; n = 1), and no viral RNA was detected in F2 progeny (0.00%; n = 0) (Table 2). The raw data underlying these confidence intervals are provided in Supplementary Table S1. All infection rates were recalculated based on the exact number of individuals tested at each developmental stage. For samples where n = 5, proportions were adjusted to reflect the true values (e.g., 4/5 = 80%), and previously reported percentages have been corrected accordingly. These revisions ensure consistency between the reported percentages, raw counts, and values presented in Supplementary Table S1. The difference in vertical transmission rates between the 27°C and 23.5°C F1 cohorts yielded a remarkably large effect size (Cohen’s h = 2.43). A post hoc power analysis confirmed that the study achieved approximately 100% statistical power (1−β ≈ 1.0) to detect this temperature-driven difference, demonstrating that the sample sizes were robust despite developmental attrition. Despite these differences in proportions, Fisher’s Exact Test showed no statistically significant difference in detection rates between generations (p = 1.00; odds ratio = 0). The full outputs of the Fisher’s Exact Test are provided in Supplementary Table S1.
Percent of Mosquitoes or Pools in Various Developmental Stages and Generations Testing Positive for Infection with USUV in the 27°C Group, and the Number of Samples Tested (N)
USUV, Usutu virus.
Percent of Mosquitoes or Pools in Various Developmental Stages and Generations Testing Positive for Infection with USUV in the 23.5°C Group
USUV detection threshold
A Ct value of 40 was used as the threshold for detection, corresponding to an estimated RNA concentration of 5.87 × 10−6 ng/μL, based on the standard curve (Fig. 1).
The mean Ct values (±SD) for mosquitoes in the 27°C generations F0, F1, and F2 were 36.43 ± 2.14, 37.39 ± 2.71, and 38.62 ± 0.45, respectively (Table 3), and 26.23 ± N/A (SD not applicable) in the 23.5°C F1 generation. All Ct values are presented as mean ± standard deviation to ensure consistency between text and tables. The predicted viral titer values were 9.6 × 10−4 ng/μL (F0), 4.5 × 10−4 ng/μL (F1), and 1.7 × 10−4 ng/μL (F2), respectively (Table 4) and 2.1 ng/μL in the 23.5°C F1 generation μL.
The Average USUV Ct for Positive Mosquitos Predicted from the USUV Standard Curve. Only Individuals from the 27°C Group Are Listed. Ct Values ≥40 Are Considered Negative
The Average Ct Values and ± SD for Positive Mosquito Life Stages and Generations at 27°C. N/A Means Not Applicable
Viral RNA in the 27°C group was detected in all mosquito life stages for F1 and F2 progeny, with the lowest viral RNA detected in F1 egg rafts (Ct 39.12), and the highest viral RNA in F1 adult female mosquitoes (Ct 35.81) (Table 3). At 23.5°C, viral RNA was only detected in the F1 adult female mosquitoes (Ct 26.23). There was no significant difference between Ct values from different mosquito generations (Kruskal–Wallis X2(2) = 2.17, p = 0.338). The effect size for this generational comparison was minimal (ε2 = 0.048), indicating that the specific filial generation (F0, F1, or F2) accounts for <5% of the variance in viral RNA load. The test outputs are provided in Supplementary Table S1. Multiple comparisons using Dunn’s test with Bonferroni adjustments confirmed no statistically significant differences among pairs; F0 with F1 (p = 0.781), F0 with F2 (p = 0.425) and F1 with F2 (p = 1.00).
No USUV RNA was detected in any negative controls. All positive controls, including blood-fed F0 mosquitoes and virus-spiked blood meals, yielded Ct values < 40 (Table 3).
To account for the relatively small sample sizes, exact binomial confidence intervals were calculated for all corrected transmission estimates. At 27°C, F1 transmission was 80% (95% CI: 37.6–96.4%), and F2 transmission was 100% (95% CI: 71.5%–100%), whereas at 23.5°C, F1 transmission was substantially lower at 1.25% (95% CI: 0.2–6.8%). Bootstrap analyses (10,000 iterations) further supported the robustness of these temperature-dependent effects despite variation in sample size, with full statistical outputs, including exact p-values and effect size estimates, provided in the Supplementary Data.
Discussion
Understanding the mechanisms by which arboviruses persist in temperate regions is essential for predicting future outbreak risks under changing climatic conditions. Our study provides robust experimental evidence that Cx. pipiens bioform molestus, an anthropophilic and urban-adapted mosquito, is capable of vertically transmitting USUV across multiple generations under specific thermal conditions. Essentially, we show that vertical transmission rate is highly temperature-dependent, occurring at 27°C but almost completely absent at 23.5°C. This finding expands current knowledge on USUV ecology and has significant implications for arbovirus maintenance and potential expansion in response to global warming.
Rationale for temperature selection
Temperature conditions were selected to represent ecologically relevant thermal scenarios for Cx. pipiens bioform molestus in temperate urban environments. We hypothesized that the 23.5°C condition represents typical late summer/early autumn temperatures in Northern European cities, including Copenhagen, Malmö, Gothenburg and Stockholm (late August–September), The 27°C condition simulates peak summer temperatures (late June–mid July) in the same regions, corresponding to optimal breeding conditions when mosquito populations reach maximum density (Bellone and Failloux, 2020; Ciota et al., 2014). This 3.5°C difference spans the critical thermal range where many flavivirus biological processes shift from suboptimal to optimal, as documented for WNV and other Culex-transmitted arboviruses (Dohm et al., 2002; Fay et al., 2021; Kilpatrick et al., 2008; Molchanova et al., 2022; Vogels et al., 2017). Furthermore, climate projections indicate that urban heat island effects and global warming will increasingly shift temperate cities from the cooler toward the warmer condition, making this comparison directly relevant to future arbovirus risk scenarios (Caminade et al., 2019; Mousavi et al., 2024).
Vertical transmission as a mechanism for persistence
Horizontal transmission of flaviviruses such as USUV, WNV, and ZIKV typically depends on the interaction between competent vectors and amplifying vertebrate hosts. However, during periods of low mosquito activity or vertebrate host availability, such as temperate winters, horizontal transmission alone may not sustain endemicity. Vertical transmission provides a complementary mechanism, allowing viruses to persist within vector populations across intergenerational gaps (Adams and Boots, 2010; Anderson et al., 2020; Anderson and Main, 2006; Dahl et al., 2022; Lequime et al., 2016; Manore and Beechler, 2015). Our findings align with this hypothesis by demonstrating successful passage of USUV from F0 to F1 and F2 generations under controlled laboratory conditions at 27°C. This result builds upon previous observations of USUV RNA in immature mosquito stages in the field (Schilling et al., 2025), and adds experimental validation of maintenance across two successive generations via vertical transmission.
Thermal gating of vertical transmission
Our most significant finding is how a static temperature difference appears to regulate vertical transmission rates. While adult females at both 23.5°C and 27°C ingested infectious blood, vertical transmission occurred almost exclusively at the higher temperature. At 23.5°C, only a single F1 mosquito tested positive (1.25%), and vertical transmission was detected in the F2 generation. This suggests that temperature likely influences vertical transmission.
The high proportion of vertically infected offspring at 27°C-94% in F1 and 100% in F2-is among the highest reported for any arbovirus in Cx. pipiens (Dahl et al., 2022; Miller et al., 2000) indicating that vertical transmission of USUV is not only feasible but its rates may be affected by temperature. Intriguingly, these transmissions were associated with high Ct values, suggesting limited viral replication in individual mosquitoes. In contrast, the single infected F1 mosquito at 23.5°C exhibited a substantially lower Ct value and a viral load nearly four orders of magnitude higher, pointing to enhanced replication at 23.5°C, despite reduced detection of viral RNA in progeny compared to 27°C. These findings suggest that temperature may differentially influence the rate of maternal transmission and the intensity of infection in progeny. These dynamics have important implications for USUV ecology in temperate urban settings. Cx. pipiens bioform molestus, which occupies thermally buffered subterranean habitats such as storm drains and basements, may sustain vertical transmission at relatively stable temperatures near 27°C.
Our findings parallel those reported for Aedes aegypti, where vertical transmission rate is modulated by environmental factors and the timing of oviposition, reflecting complex interactions between virus replication, vector physiology, and reproductive timing (Ciota et al., 2017; Comeau et al., 2020; Thangamani et al., 2016). Consistent with this, our results support the presence of a thermally regulated transovarial barrier that limits USUV inheritance at cooler temperatures, potentially restricting vertical transmission when ambient conditions fall below a critical threshold. Studies on vertical transmission in Aedes aegypti provide a well-characterized framework for understanding how flaviviruses can persist across mosquito generations. By comparing these findings to our results in Culex pipiens bioform molestus, we can contextualize species-specific differences in transmission efficiency, ecological adaptation, and potential for virus maintenance in temperate urban environments. Such comparisons highlight both shared mechanisms and vector-specific factors that influence arbovirus persistence and seasonal dynamics
Implications for vector biology and arbovirus ecology
Our study used Cx. pipiens bioform molestus, a mosquito bioform that is largely autogenous, anthropophilic, and adapted to urban underground environments, such as subway tunnels and sewer systems (Anonymous, n.d.; Fritz et al., 2015). Unlike the Cx. pipiens bioform pipiens, which is more ornithophilic and subject to seasonal diapause, Cx. pipiens bioform molestus can remain active year-round in warm microhabitats (Fonseca et al., 2004; Fritz et al., 2015). This ecological adaptation may make Cx. pipiens bioform molestus an ideal vector for sustaining arboviruses like USUV during interepidemic or overwintering periods, especially in urbanized or artificially heated settings.
Importantly, this study offers one of the few experimental demonstrations of multigenerational vertical transmission of a flavivirus under defined environmental conditions. Most existing literature either stops at F1 detection or relies on field-collected larvae with uncertain infection routes (Ciota et al., 2017; Comeau et al., 2020; Schilling et al., 2025). By demonstrating successful F2 infection, we strengthen the case that USUV can be stably inherited, rather than transiently passed to a single generation.
Climate change and emerging risk landscapes
Temperature plays a central role in shaping mosquito vector competence, virus replication, and transmission dynamics (Mordecai et al., 2017). Our data adds to growing evidence that even a modest increase in temperatures can significantly alter arbovirus persistence mechanisms (Bellone and Failloux, 2020; Lequime et al., 2016). A rise from 23.5°C to 27°C, well within the range of seasonal fluctuation in many temperate cities, was sufficient to unlock intergenerational transmission of USUV. This suggests that climate warming could expand the windows and geographic range in which vertical transmission becomes viable, thereby enhancing long-term maintenance of USUV in regions previously considered climatically marginal (Ashraf et al., 2015).
In this context, Cx. pipiens bioform molestus could play an increasingly important role in USUV ecology and epidemiology. However, we acknowledge that our findings do not demonstrate arbovirus persistence fully independent of bird–mosquito transmission cycles. The study was conducted under two controlled, static temperatures, whereas environmental temperatures in natural settings fluctuate. Therefore, while our results indicate that vertical transmission of USUV is temperature-dependent under laboratory conditions and may contribute to intergenerational maintenance, they do not provide direct evidence that USUV can be maintained without vertebrate hosts in the field.
The sharp thermal threshold observed between 23.5°C and 27°C likely reflects temperature-dependent processes in mosquito reproductive physiology and viral replication. In Culex pipiens sensu lato, vitellogenin synthesis and ovarian development are temperature-sensitive processes that directly influence the vertical transmission rate (Ferreira et al., 2020; Provost-Javier et al., 2010). Additionally, flavivirus replication rates increase exponentially within this temperature range, potentially reaching the minimum viral load threshold required for successful gonadal infection and vertical passage (Bisht and te Velthuis, 2022; Zhang et al., 2022).
Limitations and future directions
This study has several limitations that should be acknowledged. First, RT-qPCR detection of viral RNA does not confirm the presence of infectious virus; future studies should assess viral infectivity through culture methods or transmission assays to vertebrate hosts. Second, our experiments used constant temperature regimes, whereas natural environments experience fluctuating temperatures that may differently influence vertical transmission dynamics. Third, we assessed only the first oviposition cycle; evaluating successive gonotrophic cycles could provide insights into delayed viral dissemination, particularly at lower temperatures where extrinsic incubation periods are extended.
Additionally, we did not screen oviposition water for potential viral RNA contamination from infected female frass. While unlikely given rapid RNA degradation and high dilution factors in our system, future studies should incorporate environmental controls to definitively distinguish contamination from genuine vertical transmission. Finally, the viral loads detected in offspring were relatively low (high Ct values), and we did not assess dissemination to tissues relevant for horizontal transmission. Determining whether vertically infected offspring can subsequently transmit the virus to vertebrate hosts remains an important research priority.
While laboratory conditions ensure experimental rigor, field validation across diverse Culex populations and geographic regions will be essential for translating these findings into operational surveillance and risk assessment frameworks.
Conclusion
This study supports our hypotheses and demonstrates that vertical transmission of USUV in Cx. pipiens bioform molestus occurs at a high rate but may be constrained by temperature. Our findings suggest that rising environmental temperatures, whether seasonal, urban, or climate-induced, may increase the likelihood of intergenerational virus maintenance in mosquito populations, potentially facilitating the persistence and re-emergence of USUV in temperate zones. These insights underscore the need to incorporate nonhorizontal transmission pathways into arbovirus surveillance and modeling frameworks, particularly in a changing climate.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article and its Supplementary Data.
Authors’ Contributions
O.W.L., A.L., and T.L. Designed the experiments, A.L., T.L., V.N.L., A.S., V.O., and O.W.L. Coordinated with the initial optimization experiments with wild-caught mosquitoes from Uppsala prior to the main experiment. O.W.L., L.W., V.N.L., A.S., A.D., A.G.L., M.B., and D.A.M. Conducted the experiments and laboratory work. L.W., A.S., S.M., P.I.C., and J.B.O. Analyzed the data, and O.W.L., V.O., L.W., V.N.L., and A.S. Wrote the article. O.W.L., A.L., and T.L. Supervised and provided funding acquisition. All authors read and approved the final article before publication.
Footnotes
Acknowledgments
The authors would like to acknowledge Prof. Rickard Ignell of the Swedish University of Agricultural Sciences for providing us with the Culex pipiens bioform molestus colony that they used in the study. He also gave them the necessary protocols for mosquito colony maintenance.
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This work was supported by funding from the
Supplemental Material
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.