
Abstract
Background:
Triatomines are a group of nearly 158 species of hematophagous insects known as kissing bugs. Several are recognized as important vectors of Trypanosoma cruzi, the causative agent of Chagas disease. Triatoma huehuetenanguensis, a triatomine described in Guatemala in 2019, is of particular interest, as it belongs to the dimidiata subcomplex, an epidemiologically important group in southern Mexico and Central America.
Objective:
This review integrates advances regarding its discovery, distinctive morphological characteristics, life cycle, feeding and defecation patterns, geographic distribution records, and recent contributions to its genetics.
Results:
This review identifies critical knowledge gaps concerning its taxonomy, ecology, genetic structure, ecological plasticity, and potential interactions with other members of the dimidiata subcomplex. These gaps all currently limit an accurate assessment of its role in the transmission of T. cruzi. For example, there is not yet sufficient knowledge of its genetics to establish its relationship with other species in the complex.
Conclusions:
Addressing these gaps through integrative and interdisciplinary research is essential to improve vector surveillance, risk assessment, and control strategies in endemic regions.
Introduction
The subfamily Triatominae (Hemiptera: Reduviidae) includes 158 species of hematophagous insects distributed mainly in the Americas, although with some representatives in Asia and Africa (de Paiva et al., 2025; Monteiro et al., 2018). Commonly known as kissing bugs, triatomines play a relevant role in global public health because they are natural vectors of the parasite Trypanosoma cruzi, the causal agent of Chagas disease. Chagas disease is classified as a neglected disease and is a complex problem from a biological, environmental, economic, and even political perspective (for a review, see De Fuentes-Vicente et al., 2023; Hernández-Flores et al., 2025). Given the current lack of effective and accessible treatments for Chagas disease, the best tool to combat it continues to be the effective control of insect vectors.
In recent decades, the study of kissing bugs has allowed us to identify patterns in their distribution, behavior, and biology that are essential for understanding the transmission dynamics of T. cruzi. (Ceccarelli et al., 2025; De Fuentes-Vicente et al., 2018). Some species have already been identified as highly epidemiologically important, such as Rhodnius prolixus, Triatoma infestans, and Triatoma dimidiata (Carbajal-de-la-Fuente et al., 2022; Ouali and Bousbata, 2025). However, there are still significant gaps in knowledge of other species, especially recently described species such as T. huehuetenanguensis (Lima-Cordón et al., 2019).
The new description of T. huehuetenanguensis raises fundamental questions about its biology, ecology, vector capacity, geographic distribution, and interaction with humans. Integrating the available information and clarifying what is still unknown will help to guide research efforts to understand its epidemiological importance in Chagas disease.
Recent advances in integrative taxonomy and molecular systematics have reshaped the understanding of triatomine diversity in the Americas, highlighting that accurate species delimitation is essential not only for systematics but also for effective vector surveillance, risk assessment, and Chagas disease control.
Literature Search Strategy
This review was conducted as an integrative synthesis of the available literature on T. huehuetenanguensis and related species within the dimidiata subcomplex. We performed a comprehensive literature search of major scientific databases, including Web of Science, Scopus, PubMed, and Google Scholar. Searches were conducted using combinations of keywords such as “Triatoma huehuetenanguensis,” “Triatoma dimidiata subcomplex,” “Triatominae taxonomy,” “vector ecology,” “Chagas disease,” and “Trypanosoma cruzi.”
The literature search for T. huehuetenanguensis covered publications from its description in 2019 to 2025. We considered peer-reviewed articles, taxonomic descriptions, ecological studies, and epidemiological reports. Studies were included based on their relevance to the delimitation, distribution, ecology, genetics, and epidemiological significance of the species. We excluded nonpeer-reviewed sources and studies lacking primary data or a clear taxonomic context.
History of Its Discovery and Distinctive Characteristics
The specimens that contributed to the description of T. huehuetenanguensis were collected in the Department of Huehuetenango in northwestern Guatemala (Lima-Cordón et al., 2019). Previously, the specimens had been taxonomically identified as T. dimidiata based on Lent and Wygodzinsky (1979), given the considerable morphological similarities. In parallel, a genomic analysis using SNPs (single nucleotide polymorphisms) grouped the specimens into a well-supported monophyletic clade named Triatoma sp. aff. dimidiata (Justi et al., 2018).
To describe these specimens as a new species, Lima-Cordón et al. (2019) provided molecular and morphological evidence that supported its classification as a species distinct from T. dimidiata within the T. dimidiata subcomplex, which also includes T. dimidiata, Triatoma hegneri, Triatoma brailovskyi, Triatoma gomeznunezi, and Triatoma mopan (Dorn et al., 2016; Monteiro et al., 2013). These species differ in their geographic distribution and morphology. The distribution of T. hegneri is restricted to northern Central America and is primarily associated with wild environments, and T. mopan has been reported in forested areas of southern Mexico and Belize, with little evidence of domestication. However, T. gomeznunezi is distinguished by a more localized distribution and morphological features that are more distinctive compared to T. dimidiata sensu lato (Monteiro et al., 2018).
T. huehuetenanguensis shares extensive morphological overlap with T. dimidiata, but there are distinguishing morphological characters, including size (smaller than T. dimidiata) and color (lighter than T. dimidiata and without noticeable small dark spots near the spiracles). This was also observed by Cruz-Alegría et al. (2024) in populations from southern Mexico. Molecular analysis with the nuclear gene ITS-2 and the mitochondrial gene cyt b recovered specimens identified as T. huehuetanguensis as a single monophyletic clade with high support and low genetic divergence (Fig. 1). However, the taxonomic delimitation of T. huehuetenanguensis must be analyzed within the broader context of the T. dimidiata sensu lato complex, one of the most taxonomically problematic groups within the Triatominae subfamily. Several studies have documented that T. dimidiata constitutes a complex of cryptic species with marked morphological, genetic, and ecological variability across its geographic distribution, which has historically hindered the definition of clear specific boundaries (e.g., Dorn et al., 2016; Justi et al., 2018; Monteiro et al., 2013).
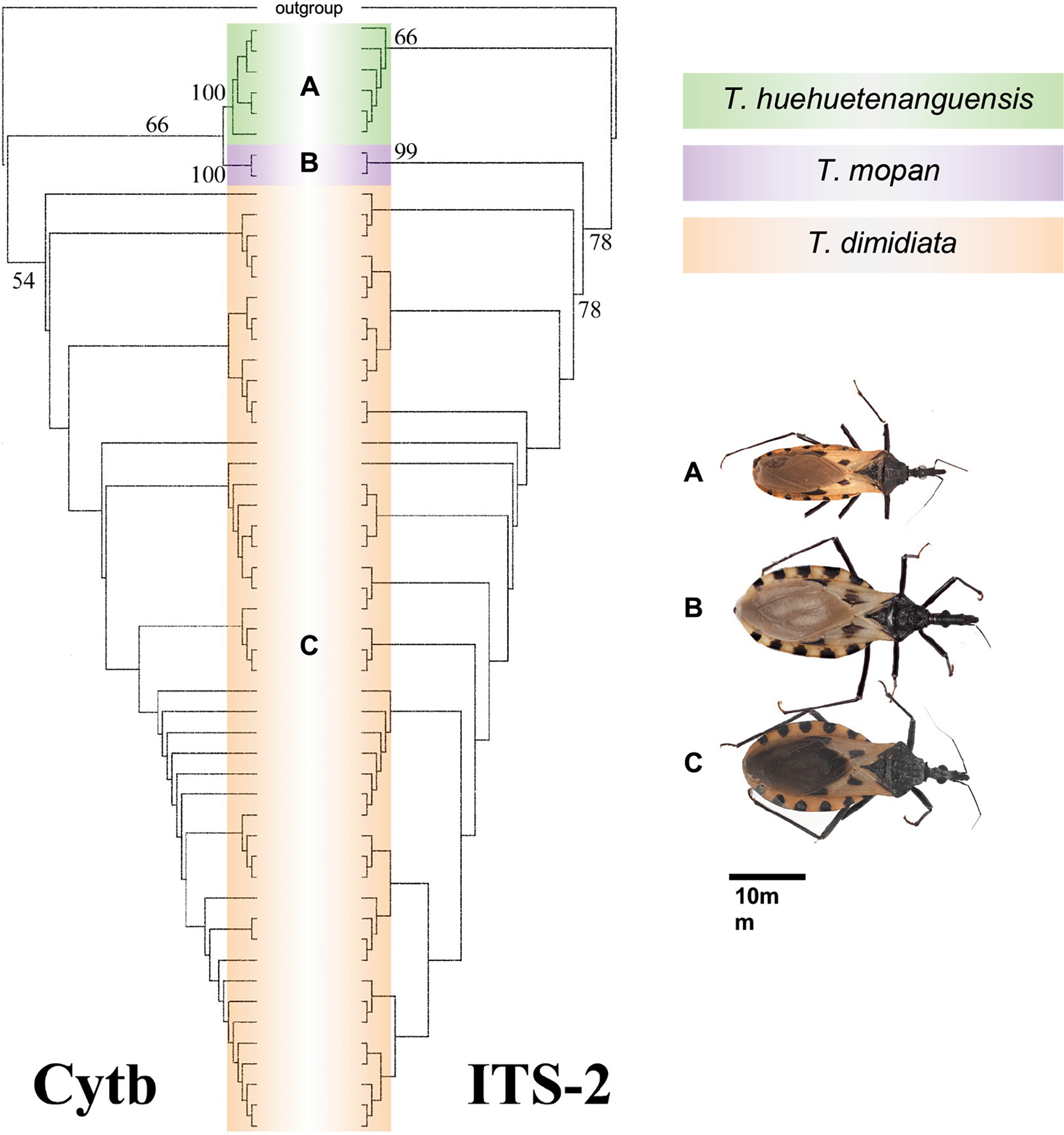
Maximum likelihood phylogenies of CytB and ITS-2, which recovered a single monophyletic clade with high support (from Lima-Cordón et al., 2019).
Thus, the differentiation of T. huehuetenanguensis relies on a set of subtle morphological characters and molecular evidence based on a limited number of markers. While this supports its recognition as a distinct species, it has also generated debate about the degree of actual evolutionary isolation from other entities within the subcomplex. The morphological overlap with T. dimidiata and the intraspecific variability observed in populations from different regions underscore the need to interpret its delimitation with caution and in an integrative manner.
Life Cycle of T. huehuetenanguensis
Like the rest of the triatomines, T. huehuetenanguensis presents hemimetabolous development that includes the egg, five nymphal stages, and the adult. To date, only one study using specimens from southeastern Mexico has evaluated its development time, the number of blood meals required for molting, cumulative mortality, time to start feeding, feeding and defecation behaviors, and fertility and fecundity (see Michel-Parra et al., 2021) (Table 1). Here, we highlight that the development time from egg to adult is 253 days for T. huehuetenanguensis, similar to the 269 days reported for T. dimidiata (Reyes and Angulo, 2009). Other species of high epidemiological relevance, such as R. prolixus and T. infestans, have shorter times of 117 days and 130 days, respectively (Arévalo et al., 2007). Furthermore, T. huehuetenanguensis requires up to 11 blood meals to complete its development, a greater number than other important species.
Some Biological Parameters of T. huehuetenanguensis Reported by Michel-Parra et al. (2021)

Defecation time is a key parameter in assessing the vectorial capacity of Triatominae species, as species that defecate while feeding have a greater probability of transmitting T. cruzi. Michel-Parra et al. (2021) determined that T. huehuetenanguensis nymphs defecate between 1 and 10 min after feeding (similar to R. prolixus and T. dimidiata, Arévalo et al., 2007; De Fuentes-Vicente et al., 2023). Adult males took between 11 and 30 min, but adult females defecated while feeding (p < 0.01). To date, a clear relationship between the sex of the insect and the time of defecation has not been established, but it appears to depend on the quality and quantity of the ingested blood and other physiological factors (De Fuentes-Vicente et al., 2018; Lunardi et al., 2015). Continuing to generate information on the defecation patterns of T. huehuetenanguensis will allow us to better understand its relevance in the transmission dynamics of Chagas disease. In this sense, it is necessary to explore how biotic and abiotic factors can influence this pattern, especially since infection with T. cruzi can affect defecation delay (Chacón et al., 2022; De Fuentes-Vicente et al., 2018). Importantly, the presence of ecological or behavioral traits associated with vectorial capacity does not necessarily imply effective transmission under natural conditions. This highlights the need to distinguish potential risk from confirmed epidemiological relevance.
Where Has T. huehuetenanguensis Been Observed?
In the Americas, Triatominae are distributed from the southern United States to southern Argentina, with higher abundance in tropical areas. Climate change scenarios predict range expansion to previously unaffected areas, underscoring the urgent need for proactive vector surveillance (Brasil et al., 2025). A recent study suggests that the distribution of kissing bugs should be grouped into seven bioregions, each of which is represented by an indicator species (Gomez et al., 2024). Occurrence data for T. huehuetenanguensis are still quite scarce, and its potential distribution has not been explored. Since its description in Guatemala, there have been additional records in southeastern Mexico, Belize, and Honduras (Fig. 2). This region, called the “Panamanian Region” by Gomez et al. (2024), includes Colombia, Ecuador, Panama, Guatemala, El Salvador, Belize, and southern Mexico, where T. dimidiata is the most important vector of Chagas disease.
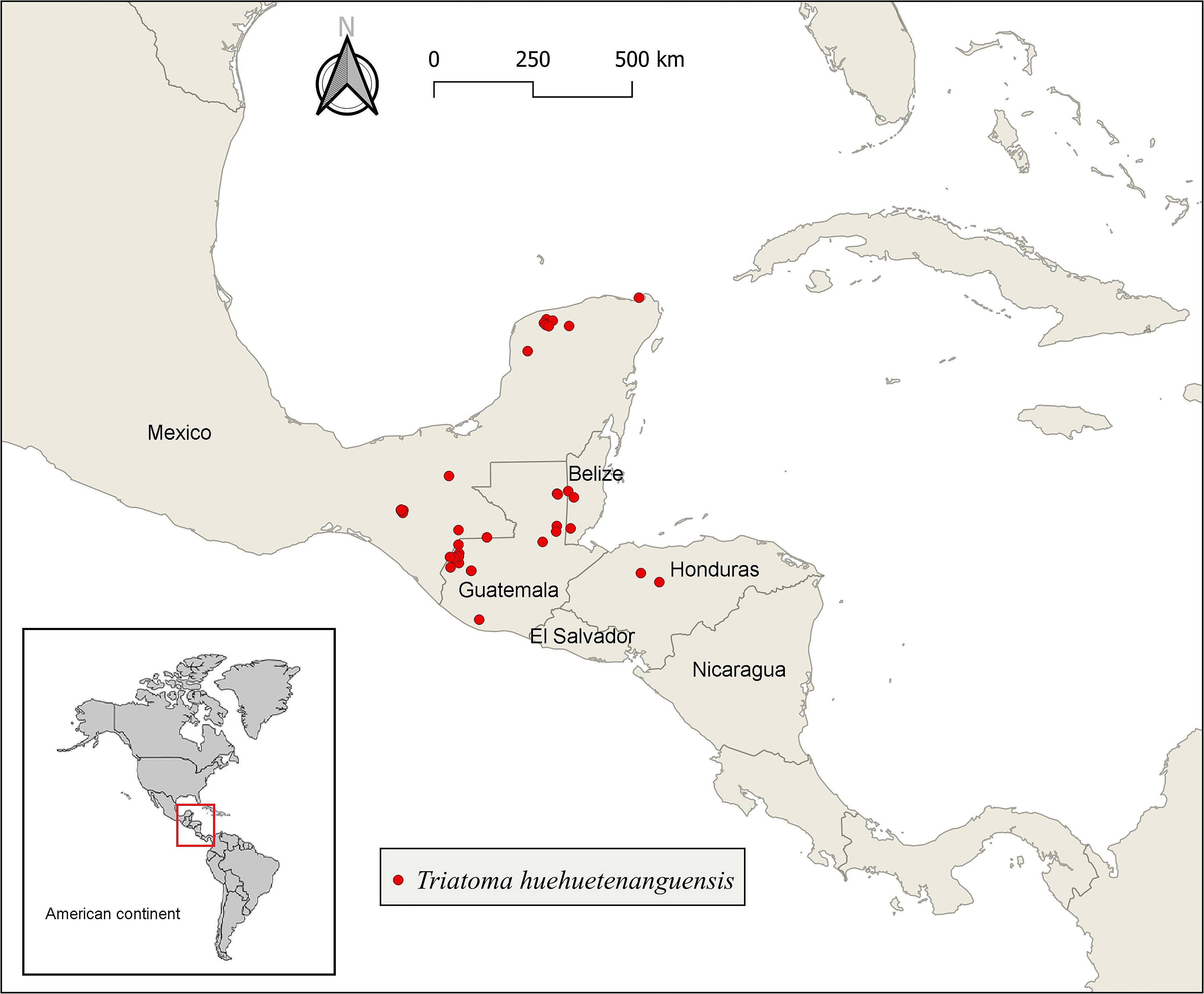
Reported records of T. huehuetenanguensis in southern Mexico and Central America.
The distribution of T. huehuetenanguensis seems to largely overlap with that of T. dimidiate. In fact, Cruz-Alegría et al. (2024) reported both species in a nature reserve in the metropolitan area of Chiapas, Mexico. This raises interesting questions, as the co-occurrence of two morphologically similar triatomine species at the same site (Fig. 3) may have several scientific and epidemiological implications (e.g., Ravazi et al., 2023; Vicente et al., 2022. To name just a few, (1) Will there be intraspecific competition or segregation between the two species due to ecological overlap?, (2) To what extent might taxonomic confusion bias distribution maps or niche models?, (3) Given their phylogenetic closeness, could coexistence favor natural hybridization events, which could further confuse identification and alter their vectorial behavior?, and (4) What would be the impact of both species on the circulation of T. cruzi? Strengthening knowledge of the geographic range of T. huehuetenanguensis should be an urgent line of research, as it would not only help delimit its distribution but also facilitate understanding of the environmental and ecological conditions that play a relevant role in its presence. This, in turn, would enable the development of predictive models for the dispersal of this species, as has been done with other species in central Mexico (Chico-Avelino et al., 2025). These models, in addition to providing information on the distribution of individual species, can help determine the coexistence of several species in a given area (Altamiranda-Saavedra et al., 2024).

Dorsal view of adult specimens of T. huehuetenanguensis and T. dimidiata.
A Large Gap in the Genetics of T. huehuetenanguensis
Genetic studies in Triatominae are essential for understanding several aspects of their biology, epidemiology, evolution, and systematics, which are fundamental elements for understanding the vector dynamics of Chagas disease transmission. From a surveillance perspective, molecular data provide valuable tools for accurate species identification in regions where morphologically similar triatomines coexist. Furthermore, greater genetic variability is associated with a greater capacity to invade different ecotopes (including the human dwelling), as is the case with T. dimidiata (Melgar et al., 2024; Monteiro et al., 2013). For now, we are still far from understanding the genetic structure of T. huehuetenanguensis and its variation between and within populations. A major contribution to the genome of this species was made by Cigarroa-Toledo et al. (2024) with populations from Yucatán, Mexico. The mitogenome was found to have an approximate length of 15,910 base pairs, slightly shorter than that of T. dimidiata (17,008 bp). The mitogenome structure consists of 13 protein-coding genes, 22 transfer RNA genes, two ribosomal RNA genes, and a control region. The characterization of the T. huehuetenanguensis mitogenome represents a significant advance in our understanding of this recently described species. Furthermore, this genomic resource opens the possibility of developing specific molecular markers for precise population determination and the detection of intraspecific genetic variation, which is key to taxonomic delimitation and understanding population dynamics. Decoding the mitogenome strengthens the foundation for evolutionary, ecological, and epidemiological research. Available genetic data, although limited, has played a key role in supporting the taxonomic recognition of T. huehuetenanguensis and its distinction from other members of the dimidiata subcomplex. Molecular markers have helped reveal genetic divergence patterns that are not always evident through morphology alone, reinforcing the importance of integrative approaches for species delimitation within Triatominae.
Thus, although available studies suggest that T. huehuetenanguensis is closely related to T. dimidiata and other species within the subcomplex, the true extent of this differentiation has not yet been fully explored. In species such as T. dimidiata, genomic analyses have revealed a complex population structure and high genetic diversity, associated with its wide distribution and adaptive capacity (Justi et al., 2018). In contrast, for T. hegneri, T. mopan, and T. gomeznunezi, the scarcity of comparable genetic studies limits the understanding of evolutionary relationships within the subcomplex and makes it difficult to assess whether the patterns observed in T. huehuetenanguensis are unique or form part of a broader evolutionary continuum.
Expanding the genetic and genomic datasets of T. huehuetenanguensis would contribute to a broader understanding of evolutionary processes such as population structure, gene flow, and possible hybridization within the dimidiata subcomplex, which are central topics in current discussions on triatomine biodiversity and systematics. There are several lines of evidence related to possible hybridization within the dimidiata subcomplex that vary in their strength and interpretation. First, laboratory studies have shown that interspecific crosses between closely related taxa within the subcomplex are possible under controlled conditions (Ravazi et al., 2023). Second, there is only limited and indirect evidence of introgression in the field: Genetic patterns in some populations may be consistent with gene flow or shared ancestry (Justi et al., 2018), but distinguishing between current hybridization and historical divergence processes remains challenging due to a lack of genomic data. Finally, it has been proposed that ecological and distributional overlap between species may facilitate contact and hybridization. While sympatry and similar habitat preferences may increase the likelihood of interaction, these conditions alone do not constitute evidence of hybridization.
Epidemiological and Public Health Implications
The presence of T. huehuetenanguensis in regions where Chagas disease is endemic raises important epidemiological implications that must be considered within the context of entomological surveillance and vector control programs in Mexico and Guatemala. Its membership in the dimidiata subcomplex, which is historically associated with a high capacity for adaptation to domestic and peridomestic environments, suggests that this species may share ecological and behavioral traits relevant to the transmission of T. cruzi (Justi et al., 2018). The detection of natural T. cruzi infection in wild and domestic populations reinforces the importance of accurately assessing (and not underestimating) its potential role as a vector in local settings.
The current lack of information on the ecology, environmental plasticity, and domestic behavior of T. huehuetenanguensis limits the ability to design specific and efficient control strategies. Previous experiences with T. dimidiata have shown that populations with high ecological variability can respond heterogeneously to conventional interventions, such as residual insecticide spraying (Melgar et al., 2024). Therefore, assuming that the two species are ecologically equivalent without empirical evidence could result in suboptimal or ineffective control strategies.
Should we consider T. huhuetenanguensis an established vector? We believe it is more appropriate to consider it a potential or suspected vector, pending further studies that assess natural infection rates in different areas, feeding behavior, and its degree of association with human environments. This distinction is important to avoid overestimating its epidemiological relevance, while acknowledging its potential role in local transmission dynamics.
Conclusions and Other Final Perspectives on T. huehuetenanguensis
The recent description of T. huehuetenanguensis represents a turning point in the study of Chagas disease vectors in the region. Although early studies on this species have allowed us to recognize its morphological, biological, and genetic differences with respect to other species, gaps remain regarding its ecology, behavior, and role in the transmission of T. cruzi. One of the central challenges remains the consolidation of its taxonomic delimitation within the T. dimidiata sensu lato complex. Within this framework, the T. dimidiata subcomplex is currently understood as a group of closely related taxa that exhibit significant morphological similarity but notable genetic structuring. This group includes T. dimidiata sensu lato and several recently described or revalidated species, such as T. mopan, T. hegneri, T. gomeznunezi, and T. huehuetenanguensis. Phylogenetic and population genetic studies suggest that these taxa represent distinct evolutionary lineages with varying degrees of divergence, even though their boundaries remain partially unresolved due to overlapping morphological traits and potential historical gene flow. Resolving this issue is not just an exercise in systematics; it is also a key aspect with direct implications for entomological surveillance, the construction of distribution maps, and the assessment of epidemiological risk. Another major challenge will be to strengthen the monitoring of T. huehuetenanguensis populations and clarify its current distribution, since its coexistence with T. dimidiata could generate scenarios of ecological overlap, hybridization, or changes in transmission dynamics that have not yet been evaluated.
Another relevant aspect will be the analysis of its ecological plasticity, that is, its ability to adapt to different habitats, including domestic and peridomestic environments. Reports in the existing literature have documented captures in wild environments (e.g., Cruz-Alegría et al., 2024) and domestic settings (e.g., Espinosa-Gonzalez et al., 2022), where natural infection with T. cruzi has been observed. Although some observations suggest that T. huehuetenanguensis may exploit peridomestic or domestic environments, current evidence remains limited and largely indirect. Therefore, inferences regarding domiciliation and epidemiological risk should be interpreted cautiously until systematic field studies confirm stable household colonization and sustained human–vector contact. Understanding these traits has been crucial in the expansion of other epidemiologically important triatomines and could determine whether T. huehuetenanguensis is a primary vector or remains restricted to wild niches. Of the 158 triatomine species, only a subset is considered intrusive with a high domiciliation capacity and is, therefore, responsible for most of the human risk of exposure to T. cruzi (Waleckx et al., 2015). Furthermore, exploring the interaction of this species with environmental (climate, vegetation, and anthropogenic disturbance) and social (housing conditions, human mobility, and the presence of domestic reservoirs) factors will be essential for assessing its potential impact on public health. Finally, the use of more robust molecular and genomic tools will open new possibilities to more precisely delimit its genetic variability, population structure, and evolution with other species within and beyond the dimidiata subcomplex. In fact, a new species of Triatominae with a very close genetic relationship to T. huehuetenanguensis was recently identified in Belize (Gunter et al., 2024). These approaches, combined with longitudinal field and laboratory studies, will allow for a clearer understanding of the vectorial role of T. huehuetenanguensis, in which only the TCI genotype of T. cruzi has been reported to date (e.g., Cruz-Alegría et al., 2024). Addressing current uncertainties surrounding T. huehuetenanguensis will require targeted research efforts focused on evolutionary processes and adaptive responses. Priorities include evaluating potential hybridization within the dimidiata subcomplex, characterizing ecological plasticity under changing environmental conditions, and applying comparative genomic approaches to refine species boundaries and improve epidemiological assessments. In summary, this species presents a scientific challenge, and future research should prioritize specific approaches to clarify the epidemiological and evolutionary relevance of T. huehuetenanguensis. Key priorities include (1) genome-wide analyses to define species boundaries, (2) ecological niche modeling to better understand their environmental constraints and potential distribution under changing conditions, (3) field studies to assess the dynamics of natural infection and transmission potential, and (4) comprehensive investigations of their association with human environments to determine their domiciliation capacity. Addressing these priorities will be essential to refining risk assessments and guiding evidence-based vector surveillance strategies.
Footnotes
Acknowledgments
This article was supported by the Secretariat of Science, Humanities, Technology and Innovation (SECIHTI) as part of the project “CBF-2025-G-1150 Contributions to the knowledge of T. huehuetenanguensis, a newly described vector of Chagas disease in southeastern Mexico.” We also thank Sofia Estrada of the Entomology Clan at UNICACH for the design of ![]() and Cinthia Karime Roblero Oroxón for reviewing the grammar.
and Cinthia Karime Roblero Oroxón for reviewing the grammar.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.