
Abstract
Background:
Mayaro virus (MAYV), an emerging and endemic arthritogenic alphavirus in tropical regions of South America, represents a growing public health threat and is primarily transmitted by Haemagogus genus mosquitoes. Although MAYV infection typically results in a self-limiting febrile illness, a significant proportion of patients develop chronic musculoskeletal symptoms, notably persistent arthralgia and arthritis.
Rationale:
This comprehensive review examines the immunopathogenesis of MAYV infection, focusing on the complex dynamics of innate and adaptive immune responses and the mechanisms contributing to postviral joint inflammation. Particular emphasis is placed on macrophage activation, inflammasome signaling (notably NLRP3), and the intrinsic susceptibility of joint-resident cells, including synoviocytes, chondrocytes, and osteoblasts. Furthermore, this review discusses recent evidence of viral persistence within tissue reservoirs as a key driver of chronic pathology and proposes a conceptual model in which MAYV persistence within joint-resident cells sustains chronic NLRP3 inflammasome activation, thereby driving long-term arthralgia. Additionally, the viral strategies for immune evasion—chiefly mediated by the nonstructural protein nsP2—are discussed in the context of interferon suppression and transcriptional shut-off.
Conclusion:
The article provides a critical evaluation of current and developing therapeutic strategies, including antiviral agents such as favipiravir and immunomodulatory agents targeting IL-1β, tumor necrosis factor, and IL-6 signaling. Understanding the interplay between viral determinants and host immune responses is critical for the rational design of effective interventions against MAYV-induced chronic inflammation and arthritis.
Introduction
Mayaro virus (MAYV) is an emerging arthritogenic alphavirus with increasing epidemic potential in the Americas. It is a positive-sense, single-stranded RNA alphavirus belonging to the Togaviridae family and is primarily found in tropical forested regions (Weaver et al., 2008). Based on genome sequencing, MAYV has been classified into three genotypes: D (predominant in South America), L (mainly found in Brazil), and N (reported in Peru) (Endy, 2020). Additionally, MAYV is part of the Semliki Forest antigenic complex, which includes Bebaru, Chikungunya virus (CHIKV), Getah, Semliki Forest, Ross River, O’nyong-nyong, and Una viruses, all of which share common antigenic sites and produce similar clinical manifestations (Acosta-Ampudia et al., 2018).
Vector arthropods, particularly mosquitoes, exhibit a high degree of adaptability to diverse ecosystems, contributing to their global distribution and facilitating the widespread transmission of arboviruses. Hematophagous insects require blood meals for oviposition, and their feeding frequency influences reproductive output. This ecological behavior increases contact rates between arboviruses and vertebrate hosts, thereby promoting sustained transmission cycles (Osorio and Arbeláez, n.d.; Lozada-Martínez et al., 2021).
Since its initial isolation in Trinidad in 1954, sporadic human cases have been reported across South American forest regions. The virus has been primarily isolated from mosquitoes of the genus Haemagogus, particularly Haemagogus janthinomys, which are considered the main sylvatic vectors (Acosta-Ampudia et al., 2018). More recently, MAYV has been detected in urban settings, likely due to increased urbanization. Evidence suggests that the introduction of arboviruses into novel environments can enable new mosquito species to participate in the transmission cycle (Weaver and Reisen, 2010; Mackay and Arden, 2016).
Although the current incidence remains low relative to that of other arboviruses, Mayaro fever poses a growing public health concern, especially in tropical and subtropical regions (Diagne et al., 2020). The geographic expansion of vector species and the clinical resemblance of MAYV infection to other arboviral diseases, such as dengue or CHIKV, complicate diagnosis, leading to underreporting and underestimation of its epidemiological burden. MAYV infection causes Mayaro fever, a dengue-like illness that is typically nonfatal but is characterized by fever, headache, rash, myalgia, and arthralgia. Notably, MAYV is associated with a persistent inflammatory response in the joints during the postacute phase, which may result in viral arthritis clinically similar to that caused by CHIKV. This includes chronic joint inflammation, pain, and stiffness. It is estimated that between 50% and 89% of patients develop arthralgia, while ∼75% report myalgia. Musculoskeletal symptoms may persist for several months. Chronic arthralgia has been reported in up to 54% of patients 1 year after the acute phase. The most commonly affected joints include the hands (63%), knees (38%), wrists (25%), ankles and feet (25%), elbows (13%), and shoulders (7%) (Wei et al., 2024).
As MAYV infection has the potential to induce prolonged inflammatory responses that may persist beyond the acute phase (Mackay and Arden, 2016; Acosta-Ampudia et al., 2018), this review highlights its possible role in the pathogenesis of postviral inflammatory joint disorders. Based on the available evidence, we propose a conceptual model in which MAYV establishes persistence within joint-resident cells, such as synovial fibroblasts and macrophages. This localized viral presence promotes the sustained production of proinflammatory cytokines, thereby driving persistent arthralgia in infected patients.
Immune Response to MAYV
MAYV is transmitted to humans through the bite of infected female mosquitoes, primarily of the Haemagogus genus. Upon inoculation, the virus enters the bloodstream either as free virions or within infected monocytes. Viral replication occurs in leukocytes and lymphoid organs such as the spleen and liver, which serve as primary sites of viral amplification (Chen et al., 2015; Mackay and Arden, 2016; Wei et al., 2024).
Subsequently, the virus spreads to muscles, bones, and joints, where it triggers potent innate immune activation via receptors such as RIG-I, MDA-5, TLR3, and TLR7 (Chen et al., 2015; Tasso de Oliveira et al., 2015; Fros and Pijlman, 2016; Wei et al., 2024). This activation leads to the production of type I interferons, proinflammatory cytokines (interleukin-6 [IL-6], tumor necrosis factor alpha [TNF-α], and IL-1β), and chemokines (CCL2 and CXCL10), all of which play key roles in regulating both local and systemic inflammation (Chen et al., 2015; Tasso de Oliveira et al., 2015; Wei et al., 2024). Recent studies have elucidated some of the immunological mechanisms underlying persistent inflammatory complications, revealing an imbalance in cytokine and chemokine levels that may account for prolonged joint inflammation (Mostafavi et al., 2019; Ferreira et al., 2020). During the early phase of infection, the immune system mounts a strong inflammatory response. Research conducted in patients from Peru and Europe have detected significant elevations in cytokines such as IL-12p70, IL-10, and Vascular Endothelial Growth Factor (VEGF), which are markers associated with a Th1-type response and vascular repair processes. Increased levels of IL-6, IL-8, and MCP-1 have also been observed, all of which are key molecules involved in recruiting immune cells to the joints. Interestingly, while some studies have reported an early rise in TNF-α in in vitro models, others have reported reduced levels in human patients, suggesting that TNF-α regulation may vary depending on the infection context, possibly reflecting species-specific immune responses or differences in infection stages (Santiago et al., 2015; Hernández-Sarmiento et al., 2024).
Multiple studies have demonstrated that the persistence of articular symptoms, such as chronic arthralgia, is associated with sustained cytokine expression, including IL-13, IL-17, IL-7, and VEGF, even several months after acute infection (Suhrbier and Mahalingam, 2009; Hean Tang et al., 2022). Notably, patients who develop postviral arthritis exhibit persistently elevated levels of inflammatory mediators, whereas IL-10 (an anti-inflammatory cytokine) is markedly decreased, suggesting an immunological imbalance that favors joint damage (Ferreira et al., 2020).
Macrophages play essential roles in several inflammatory joint diseases, including rheumatoid arthritis, spondyloarthropathies, and gout arthritis. Importantly, they have also been implicated in the pathogenesis of infectious arthritis, such as septic arthritis and Lyme arthritis. In alphavirus infections, macrophages are a major component of the cellular infiltrate in affected tissues, contributing significantly to the inflammatory milieu (de Castro-Jorge et al., 2019).
It has been demonstrated that MAYV can infect both murine and human macrophages, inducing their apoptosis and promoting the release of TNF-α and reactive oxygen species (ROS), which are key elements of inflammation and tissue damage (Mackay and Arden, 2016; Diagne et al., 2020; Ferreira et al., 2020). Activation of the NLRP3 inflammasome, particularly in bone marrow-derived macrophages, has also been documented as a central mechanism driving IL-1β production and amplifying the inflammatory response. This process contributes significantly to the inflammatory milieu dominated by the aforementioned proinflammatory cytokines (de Castro-Jorge et al., 2019; Mostafavi et al., 2019).
Furthermore, key resident cells of the joint tissue, such as osteoblasts, chondrocytes, and synoviocytes, are susceptible to MAYV infection. These cells not only support viral replication but also actively secrete the key inflammatory mediators previously described, further promoting immune filtration (Diagne et al., 2020). Additionally, infection stimulates monocyte osteoclastogenic activity by recruiting circulating monocytes to bone tissue and exposing them to macrophage colony-stimulating factor and receptor activator of nuclear factor kB ligand (RANKL), which are produced by osteoblasts, stromal cells, and activated T-cells. RANKL binds to its receptor, RANK, expressed on monocytes, triggering intracellular signaling pathways such as NF-kB and NFATc1 (the master transcription factor for osteoclast differentiation), ultimately leading to the fusion of monocytes into multinucleated osteoclasts. Moreover, the inflammatory cytokines mentioned above further increase RANKL expression and directly enhance osteoclastogenesis, thereby contributing to bone damage and cartilage erosion (Chen et al., 2015; Diagne et al., 2020; Engdahl et al., 2025). These findings suggest that MAYV not only induces self-limited viral arthritis but may also lead to a debilitating long-term condition.
Although neutralizing antibodies (IgM and IgG) are produced in most patients, their presence does not always correlate with the resolution of articular symptoms. Some individuals with adequate IgG titers still develop persistent arthritis, reinforcing the notion that MAYV-induced immunopathology may be independent of viral neutralization and more closely linked to inflammatory profiles and host cellular susceptibility (Fros and Pijlman, 2016). Furthermore, emerging evidence suggests that these cytokine profiles may exhibit sexual dimorphism. Drawing on related arthritogenic alphaviruses, biological sex appears to modulate the magnitude of the innate response, with females often exhibiting higher levels of proinflammatory mediators, potentially due to the influence of estrogens on Toll-like receptor (TLR) and type 1 interferon pathways (Berghöfer et al., 2006). The presence of estrogen response elements in the promoters of several cytokine genes suggests that the female hormonal environment could exacerbate the NLRP3-driven inflammatory loop, thereby leading to the chronic joint symptoms observed in MAYV-infected women (Rio et al., 2025).
Collectively, these findings indicate that post-MAYV arthritis results from an exacerbated and sustained inflammatory response in joints and related tissues, modulated by macrophage activation, inflammasome signaling, ROS production, and the involvement of resident cells such as synoviocytes and osteoblasts (Fig. 1). The identification of biomarkers, such as IL-13, IL-17, or VEGF, may be key for predicting severe disease and developing targeted immunomodulatory therapies (Fros and Pijlman, 2016; Diagne et al., 2020; Ferreira et al., 2020).
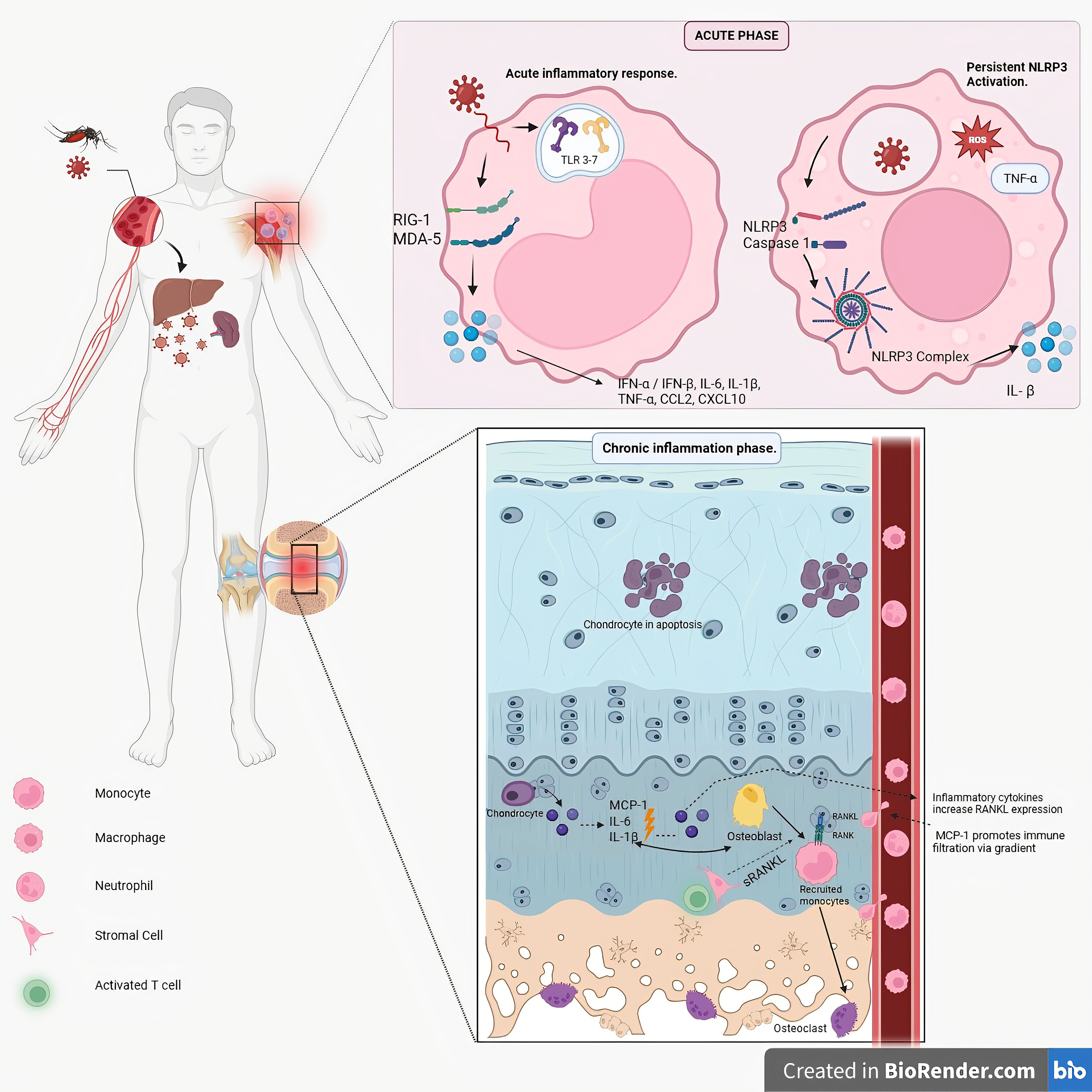
Chronic joint inflammation induced by the immune response to Mayaro virus (MAYV). Following the bite of an infected mosquito, MAYV spreads through the bloodstream, initially targeting immune organs such as the liver and spleen, and later peripheral tissues, including muscle, joints, and bone. Viral recognition by Pattern recognition receptors (PRRs) triggers the production of type I interferons, IL-6, TNF-α, and IL-1β, promoting immune cell infiltration. Macrophages activate the NLRP3 inflammasome, leading to IL-1β secretion and intensified joint inflammation. MAYV also infects osteoblasts, chondrocytes, and synoviocytes, enhancing viral replication, inflammatory mediator release, osteoclastogenesis and osteoclast activation, resulting in bone erosion, cartilage damage, and chronic postviral arthritis.
Viral Evasion Mechanisms
One of the most relevant features of MAYV is its ability to evade the host immune system. Recent studies have shown that this virus suppresses the production of type I and III interferons from the early stages of infection, disrupting the host’s early innate immune signaling cascades before an adequate immune response can be mounted. In particular, IFN-β production is strongly suppressed in infected cells, depriving them of a crucial factor for controlling viral infections (Akhrymuk et al., 2012; Ishida et al., 2021; Liu et al., 2022).
The primary viral element involved in evading antiviral responses is the nonstructural protein nsP2, whose significance has also been established in other alphaviruses, such as CHIKV and Sindbis virus (SINV) (Santiago et al., 2015; Scholte et al., 2015; de Castro-Jorge et al., 2019). In the context of MAYV, nsP2 effectively suppresses interferon induction by acting downstream of IRF3 phosphorylation and nuclear translocation, suggesting that it directly interferes with the transcription and/or translation of IFN-encoding mRNAs. Therefore, an nsP2 mutant with reduced transcriptional silencing capacity (nsP2 P722S) was significantly less effective at inhibiting this pathway, underscoring the importance of this silencing mechanism (Akhrymuk et al., 2012; Ishida et al., 2021).
Far from targeted interference, nsP2 promotes global transcriptional silencing by binding to and degrading essential proteins in the RNA polymerase II machinery, such as Rpb1 and TFIIE2, thereby inducing rapid and global host shutoff. This molecular hijacking not only prevents the synthesis of antiviral mRNAs but also facilitates the preferential translation of viral proteins by depleting the pool of competing cellular transcripts, a critical determinant of the high viral loads observed during the acute phase (Akhrymuk et al., 2012; Fros and Pijlman, 2016; Ishida et al., 2021). Although Rpb1 degradation is proteasome-dependent, the mechanism underlying the loss of TFIIE2 remains unclear. Interestingly, other alphaviruses, such as Semliki Forest virus, also reduce TFIIE2 levels, whereas SINV does not, indicating divergent strategies within the genus (Akhrymuk et al., 2012; Ishida et al., 2021).
In addition to its role in suppressing antiviral responses, nsP2 from CHIKV and MAYV has been shown to interact with host proteins involved in stress granule formation, such as G3BP1 and G3BP2, thereby preventing the activation of cellular mechanisms that would typically restrict viral replication. This confers a replicative advantage to the virus during the early stages of infection (Lum and Ng, 2015; Scholte et al., 2015). These multifaceted immune evasion strategies contribute to viral persistence, thereby shaping the subsequent chronic inflammatory milieu.
Proposed Mechanism Model: Tissue Sequestration and Chronic Inflammation
Expanding upon previous observations of arthritogenic alphaviruses, we propose an expanded, integrative model of MAYV pathogenesis (Fig. 1).
Beyond traditional immune evasion, emerging evidence suggests that “tissue sequestration” is a key driver of MAYV chronicity. This mechanism suggests that the virus does not merely circumvent the initial host defense but actively establishes long-term reservoirs within specific cellular niches, such as synovial fibroblasts and tissue-resident macrophages, as well as recently identified reservoirs in reproductive tissues (Guzman et al., 2025; Arévalo et al., 2026).
These “sanctuary sites” effectively protect viral RNA and antigens from neutralizing antibodies and systemic clearance, facilitating a state of low-grade persistent inflammatory stimulus. Consequently, the localized presence of viral remnants acts as a chronic trigger for NLRP3 inflammasome activation, providing a robust molecular explanation for why joint debilitating symptoms persist long after the resolution of systemic viremia. Chronic activation of this pathway may drive sustained production of IL-1β and IL-18, thereby maintaining a low-grade inflammatory state that underlines prolonged arthralgia (de Castro-Jorge et al., 2019; Guzman et al., 2025). This hypothesis is further supported by recent evidence showing viral antigen persistence in maternal and placental compartments, even in the absence of detectable infectious particles (in murine models) (Arévalo et al., 2026).
We further propose that early immune evasion mechanisms, especially nsP2-mediated host transcriptional shutdown and interferon suppression, create a permissive environment for the establishment of these viral reservoirs (Akhrymuk et al., 2012). This temporal coupling between acute immune suppression and subsequent persistence represents a critical, yet unexplored transition in MAYV infection. These viral sanctuary niches may function as immunologically protected microenvironments, limiting the access of neutralizing antibodies and cytotoxic lymphocytes. The persistence of viral RNA or antigens within these compartments provides a continuous source of innate immune stimulation (Guzman et al., 2025).
Finally, we propose that host-related factors, including biological sex, may modulate both viral persistence and inflammatory outcomes. Sex-dependent differences in immune responses, potentially mediated by hormonal regulation and differential cytokine profiles, could influence the stability of viral reservoirs and the magnitude of chronic inflammation, representing an important direction for future investigation, as recent studies have highlighted transient sex-dependent differences in viral distribution during the acute phase (Rio et al., 2025; Arévalo et al., 2026). Collectively, this integrative model links early immune evasion, viral persistence within tissue niches, and sustained innate immune activation into a framework that may explain the chronic inflammatory phenotype associated with MAYV infection.
Potential Immunomodulatory Therapies and Control of Persistent Inflammation in MAYV Infection
Numerous studies have been conducted to identify effective treatments against MAYV. However, the diagnosis of this particular virus remains challenging due to its clinical similarities with other arboviruses, such as CHIKV and dengue virus. Consequently, MAYV infections are often overlooked or misdiagnosed until more severe symptoms, such as persistent joint inflammation, emerge. In light of this, a review of potential therapeutic approaches was carried out, focusing on treatments that may be beneficial both during acute infection and in the management of postinfectious joint complications (Table 1).
Antiviral and Immunomodulatory Therapies for MAYV Infection

CAPS, cryopyrin-associated periodic syndromes; CHIKV, Chikungunya virus; MAYV, Mayaro virus; RA, rheumatoid arthritis.
One of the most extensively studied therapeutic candidates is favipiravir, a broad-spectrum antiviral agent. In experimental studies using murine models, favipiravir was administered before, during, and after MAYV infection. Treatment resulted in a significant reduction in viral titers in both blood and tissues when administered either prophylactically or early in infection. However, no reduction in viral replication was observed when the drug was given after infection had been established, indicating that its antiviral efficacy is time-dependent and largely limited to early intervention stages (Scholte et al., 2015).
Other studies have proposed the use of plant-derived virucidal compounds. For instance, a flavonoid isolated from the roots of Maytenus imbricata exhibited direct antiviral activity against MAYV. Notably, this compound acted directly on viral particles without affecting host cells. The 50% inhibitory concentration (IC50) was reported to be 37.9 ± 2.4 μM, with a selectivity index greater than 43, suggesting high antiviral potency with low cytotoxicity (Andreolla et al., 2022).
Additional therapeutic strategies are focused on suppressing the exaggerated inflammatory response triggered by the virus. While the immunopathology of MAYV remains under investigation, several mechanisms contributing to chronic inflammation have already been identified. One such mechanism is activation of the NLRP3 inflammasome, which has emerged as a key component in the immunopathogenesis of MAYV, particularly in the context of persistent joint inflammation. A pivotal study in murine models by de Castro-Jorge et al. (2019) demonstrated that MAYV induces robust activation of the NLRP3 inflammasome in macrophages, leading to the maturation and secretion of inflammatory cytokines, including IL-1β and IL-18. Remarkably, in NLRP3-deficient mice, significant reductions in local inflammation, tissue swelling, and immune cell recruitment were observed following infection, positioning NLRP3 as a promising therapeutic target for mitigating virus-induced joint damage (Coll et al., 2015; de Castro-Jorge et al., 2019).
As a result, various immunomodulatory agents have been proposed to block this inflammatory axis. These include anakinra (an IL-1 receptor antagonist), canakinumab (a monoclonal antibody against IL-1β), and MCC950 (a selective NLRP3 inhibitor), which is currently approved for other inflammatory conditions such as rheumatoid arthritis and cryopyrin-associated periodic syndromes. Although these agents have not yet been tested clinically in the context of MAYV infection, their proven efficacy in other inflammatory and autoinflammatory diseases highlights their potential as relevant therapeutic options for future clinical research in MAYV-associated arthritis (Coll et al., 2015; de Castro-Jorge et al., 2019).
Another promising approach involves the use of immunomodulatory therapies previously evaluated in cases of arthritis induced by CHIKV. Among these, the application of TNF inhibitors and IL-6 receptor antagonists, such as tocilizumab, have been proposed as a potential strategy to mitigate chronic inflammation and joint damage (Ng, 2017; Bilsborrow et al., 2021). Nevertheless, clinical caution is imperative when timing immunomodulatory interventions. Administration of TNF-α or IL-6 inhibitors during the acute phase of infection may inadvertently compromise the host’s primary antiviral defenses, potentially facilitating unchecked viral dissemination. Given that type I interferon responses and early inflammatory signaling are critical for restricting alphavirus replication, such targeted immunosuppression should be reserved for the postviral chronic phase, where persistent joint inflammation is driven by dysregulated immune pathways rather than active viral replication (Ng, 2017; Bilsborrow et al., 2021; Guzman et al., 2025).
Current Gaps and Future Directions
Despite significant advances in understanding the immunopathogenesis of MAYV infection, several critical gaps remain that hinder the development of targeted clinical interventions. One of the most pressing unresolved questions concerns the precise molecular mechanisms underlying long-term viral persistence within joint tissues. While emerging studies support the concept of “tissue sequestration,” the specific cellular reservoirs, particularly the roles of synovial fibroblasts and resident macrophages as sanctuary sites, remain poorly defined in human subjects. Future research must determine the stability of residual viral RNA or antigens and their capacity to act as chronic pathogen-associated patterns that drive sustained inflammation (Guzman et al., 2025).
Another major limitation is the incomplete characterization of the adaptive immune response and its transition to a chronic state. Although persistent cytokine signatures (IL-13, IL-17, and VEGF) have been associated with chronic arthralgia, the specific contributions of T-cell subsets, B-cell responses, and the potential for “molecular mimicry” in disease progression remain incompletely understood. In particular, the involvement of Th17-driven responses and their synergistic relationship with the NLRP3 inflammasome in joint destruction warrants deeper investigation to identify novel therapeutic axes (Suhrbier and Mahalingam, 2009; Wauquier et al., 2011; Hean Tang et al., 2022).
Additionally, there is a notable absence of longitudinal human cohort studies that integrate clinical, virological, and immunological data. Much of the current knowledge is extrapolated from cross-sectional human data or limited preclinical models, which may not fully capture the complexity of human immunopathology. Furthermore, recent findings highlight a significant translational gap regarding sex-dependent differences in MAYV chronicity; investigating how the host hormonal environment modulates viral evasion and inflammatory resolution is now a priority for personalized medicine. The identification of MAYV in reproductive tissues and sperm introduces a critical new dimension to its pathogenesis, suggesting that these tissues may not only act as immunologically protected reservoirs contributing to chronicity but also raise the biological plausibility of nonvector transmission routes, such as sexual or vertical transmission (Rio et al., 2025; Arévalo et al., 2026).
From a therapeutic perspective, the lack of validated antivirals or immunomodulators specifically approved for MAYV remains a critical hurdle. While strategies such as NLRP3 inhibition or natural compounds like Silymarin have shown recent promise in murine models, their safety profiles and optimal administration windows in humans require rigorous evaluation. Future investigations should also prioritize characterizing the persistence of viral RNA in these specific niches and evaluating its implications for public health, as alternative transmission cycles could significantly alter the epidemiological dynamics and clinical management of MAYV-infected patients during and after the acute phase.
Conclusion
MAYV represents a growing virological and immunological challenge in the Americas. Although often overshadowed by other arboviruses, its ability to induce long-lasting joint inflammation highlights the need for increased surveillance, clinical recognition, and biomedical research. The evidence presented in this review underscores that persistent arthritic symptoms are not solely the result of ongoing viral replication but rather a consequence of a dysregulated and sustained inflammatory response involving cytokines such as IL-6, IL-17, and TNF-α, as well as the activation of the NLRP3 inflammasome. Moreover, MAYV’s immune evasion capabilities, particularly through nsP2-mediated interferon suppression, enhance viral persistence and pathogenicity. While no specific antiviral or immunomodulatory therapy is currently approved for MAYV, emerging data support the repurposing drugs used in other viral or autoimmune conditions. Future research should focus on identifying predictive biomarkers of chronic inflammation and conducting clinical trials to assess targeted immunotherapies. Addressing these gaps is essential for mitigating the long-term burden of MAYV infection and improving outcomes for affected individuals.
Finally, the potential for MAYV adaptation to urban vectors and the risk of large-scale outbreaks underscore the need for enhanced epidemiological surveillance and vector competence studies. Integrating ecological, virological, and immunological approaches will be essential to bridge the current translational gap and mitigate the long-term public health impact of MAYV in the Americas.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Disclaimers
The opinions expressed in this article are our own and do not constitute the institution’s official position.
Ethics Approval
This is a literature-based review. The Research Ethics Committee from Universidad Cooperativa de Colombia has confirmed that no ethical approval is required.
Authors’ Contributions
J.C.H. and N.A.T.: Conceptualization. T.M.G.: Writing—original draft. J.M.A.-C., J.C.H., and N.A.T.: Writing—review and editing. T.M.G. and J.C.H.: Investigation. T.M.G.: Visualization. J.C.H. and N.A.T.: Project administration.
Footnotes
Acknowledgments
The authors thank the Universidad Cooperativa de Colombia, the Universidad Autónoma de Nayarit, and the Corporación Universitaria Remington for supporting this research.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Universidad Cooperativa de Colombia, the Universidad Autónoma de Nayarit, and the Corporación Universitaria Remington. The funders had no role in the study design, data collection and analysis, publication decision, or article preparation.