
Abstract
Background:
Vector-borne diseases (VBDs) continue to pose major public health challenges, particularly in tropical and subtropical regions. Conventional vector control strategies, including insecticide spraying, insecticide-treated bed nets, and larvicidal interventions, have substantially reduced disease burden; however, the emergence of insecticide resistance has compromised the long-term effectiveness of these approaches. Consequently, environmentally sustainable biological and genetic vector control methods are increasingly being explored as alternative strategies.
Materials and Methods:
This review comprehensively evaluates recent advances in biological and genetic approaches for mosquito vector control. The article synthesizes evidence from published laboratory, semi-field, and field studies on Wolbachia-mediated pathogen blocking, paratransgenesis using Asaia and Serratia, entomopathogenic fungi, and genetic control tools, including the sterile insect technique (SIT), incompatible insect technique (IIT), release of insects carrying a dominant lethal (RIDL), and CRISPR–Cas9-based gene drive systems.
Results:
Multiple biological and genetic interventions demonstrated substantial reductions in mosquito populations and pathogen transmission under experimental and field conditions. Wolbachia-based systems, SIT, IIT, and RIDL approaches showed promising operational feasibility and field validation, whereas gene drive technologies exhibited strong transformative potential but remain largely experimental. However, significant challenges persist, including scalability, infrastructure requirements, ecological uncertainty, biosafety concerns, regulatory complexity, and ethical considerations. Comparative evaluation indicates that these approaches differ considerably in technological maturity, deployment readiness, and sustainability.
Conclusions:
Emerging biological and genetic vector control strategies represent promising alternatives to conventional insecticide-based approaches for sustainable VBD management. Rather than relying on a single universal intervention, context-specific integration of complementary approaches within existing integrated vector management frameworks, supported by community participation and effective governance, is likely to provide more durable and environmentally sustainable solutions for controlling vector-borne diseases.
Introduction
Vector-borne diseases (VBDs) are caused by different pathogens that are transmitted through insect vectors. Historically, VBDs have contributed significantly to the health burden of human beings in tropical and subtropical regions of the globe. VBDs account for about 17% of all infectious diseases every year and contribute to about 700,000 deaths globally (WHO, 2024). Several genera and species of mosquitoes are known as disease vectors responsible for the transmission of various life-threatening infectious diseases, including malaria, dengue fever, chikungunya, Zika, and yellow fever (WHO, 2024). For example, India, being a tropical country, is highly endemic for many VBDs. This is principally due to diverse eco-climatic conditions, forest areas, and the diversity of vectors and pathogens that provide the most conducive environment for the spread of different VBDs (Bordoloi and Saharia, 2021). In this review, we primarily focus on mosquito-borne diseases, including malaria, dengue, chikungunya, and Zika, and highlight recent advances in biological and genetic vector control strategies. This review is based on literature published over the past decade and follows a narrative approach to synthesize current knowledge, identify research gaps, and discuss future directions.
To combat VBDs, the World Health Organization (WHO) has developed a strategic program called the Global Vector Control Response 2017–2030 (WHO, 2017). Under this program, various countries, especially those in the tropics and subtropics, have contributed significant resources to mosquito control programs. Although various programs to control vector populations are ongoing, outbreaks continue to occur, further extending the burden of infectious diseases geographically due to the complex interaction among the pathogen, vector, and human. Additionally, factors such as migration due to global trade, unplanned urbanization, and environmental dynamics (i.e., climate change and global warming) further influence the spread of VBDs. Conventional approaches to controlling vectors and, in turn, VBDs have relied heavily on chemical insecticides used in indoor residual spraying (IRS) and outdoor fogging with Dichlorodiphenyltrichloroethane (DDT), insecticide-treated nets, and larvicides (Kumar et al., 2024). Despite the historical efficacy of these methods in reducing the burden of VBDs, these approaches are facing limitations due to emerging resistance, particularly to DDT, pyrethroids, and organophosphates in vectors (Khan et al., 2020). Such emergence of insecticide resistance among various vectors has reduced the effectiveness of these interventions. Moreover, indiscriminate use of chemical insecticides raises concerns regarding human health and ecology. For example, the integration of these chemicals into the environment and their toxicity in nontarget organisms, flora, and fauna are well documented (Pathak et al., 2022). These concerns have led to a critical evaluation of existing VBD control measures and initiatives toward more sustainable and biologically driven alternatives. In this communication, we provide a comprehensive update on various control measures for VBDs that are used in different regions of the globe, targeting different VBDs (Fig. 1). The collated information will not only be helpful to vector biologists for the development of research-based vector control strategies but also for policymakers choosing appropriate control measures targeting specific disease vectors. This narrative review critically evaluates major biological and genetic mosquito control innovations developed or substantially advanced over the past decade, with emphasis on their mechanisms, field validation, implementation feasibility, and future challenges.
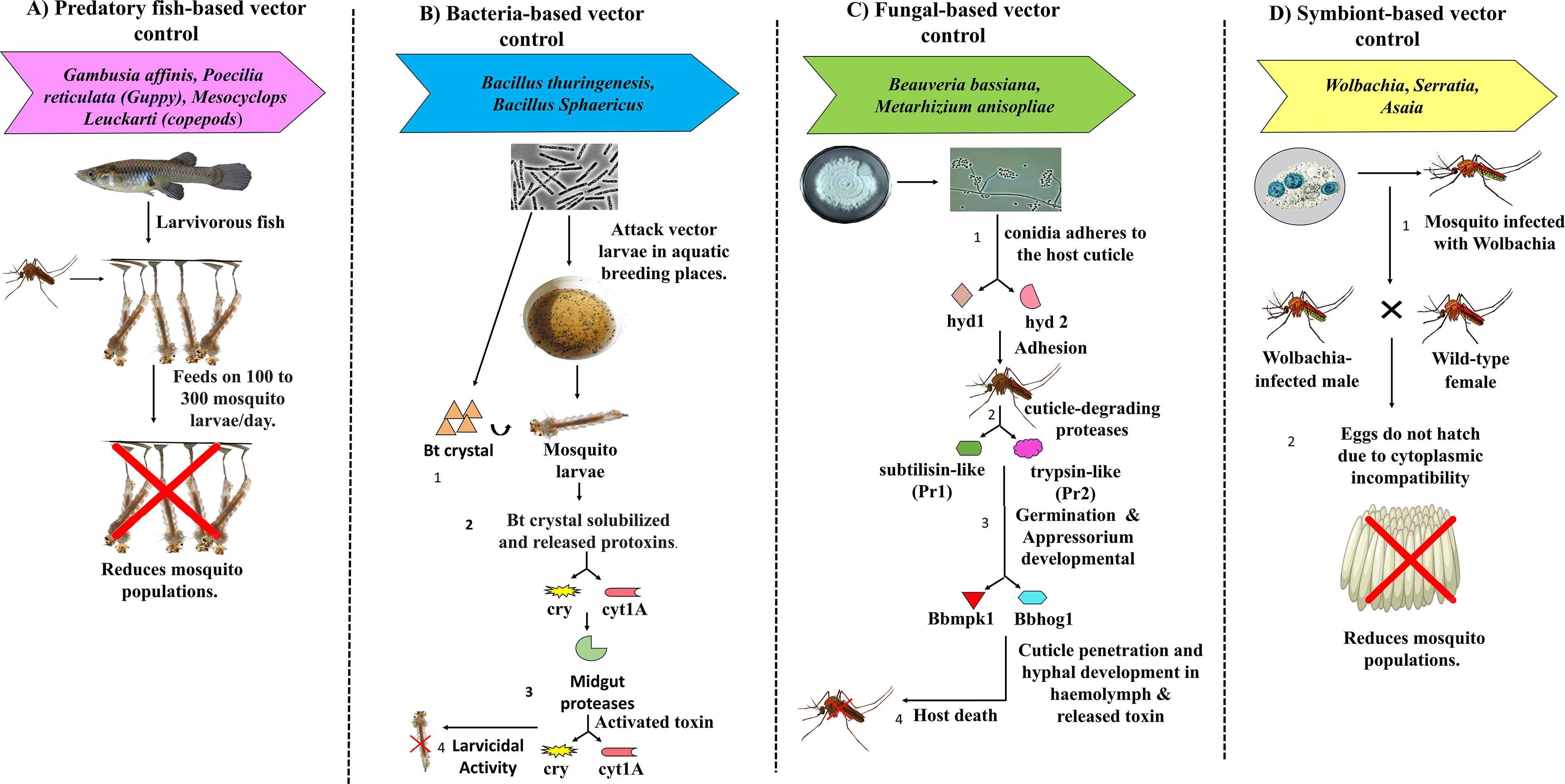
Various biological methods for controlling mosquito vectors. (
Biological Control Measures
Wolbachia-based vector control
Wolbachia is an intracellular Gram-negative endosymbiotic bacterium of the class Alphaproteobacteria, maternally inherited through vertical transmission from females to their offspring. Generally, Wolbachia facilitates its own proliferation and transmission by manipulating host reproduction through feminization, male killing, parthenogenesis, or cytoplasmic incompatibility (CI) (Fig. 1D). Natural infection with this bacterium has been detected in various genera of medically important insect species. However, Wolbachia infection has never been detected in Aedes aegypti, the primary vector for dengue, Zika, and chikungunya viruses. Approximately 66% of all insect species naturally harbor different strains of Wolbachia pipientis. Research over the last two decades on Wolbachia infections has grown rapidly, and many studies have shown successful Wolbachia introduction into A. aegypti. Results from these studies have shown two major biological outcomes: (1) suppression of pathogen replication, known as “pathogen blocking,” and (2) induction of CI in the host system, which reduces mosquito fertility by manipulating reproduction (Walker et al., 2011, Utarini et al., 2021). CI occurs when uninfected females mate with Wolbachia-infected males, resulting in embryo lethality or nonviable offspring, ultimately leading to population reduction (Fig. 1D). These experimental results provide a new strategy for vector control through either population suppression or population replacement by substituting wild populations with high transmission capacity with less competent vector species (O’Neill, 2018). Under the World Mosquito Program, experimental field trials with Wolbachia-infected mosquitoes in multiple countries—including Colombia (Velez et al., 2023), Vietnam (Turner et al., 2023), Brazil (Pinto et al., 2021), Indonesia (Indriani et al., 2020), and Australia (Ogunlade et al., 2023)—have demonstrated significant reductions in arthropod-borne virus (arbovirus) transmission and long-term establishment of Wolbachia-infected mosquito populations. A recent randomized trial conducted in Yogyakarta, Indonesia, showed a 77% reduction in dengue incidence after the release of Wolbachia-infected mosquito vectors (Utarini et al., 2021). Despite these successes, several challenges remain, including temperature sensitivity of some Wolbachia strains (e.g., wMel), incomplete maternal transmission, fitness costs, and potential competition with native microbiota.
Paratransgenesis: Asaia and Serratia
Paratransgenesis is an innovative method that involves genetically modifying symbiotic microorganisms, such as bacteria, fungi, and viruses, that are already present in mosquitoes to deliver anti-pathogen molecules within the vector’s body, preventing them from spreading pathogens. These microorganisms are constantly associated with the host system and can be found in different organs, for example, the midgut, salivary glands, and reproductive organs. Unlike transgenesis, in which modification occurs directly in the mosquito genome, paratransgenesis uses these microorganisms to manipulate the host system where the growth of infectious agents takes place. These symbiotic microorganisms are genetically engineered to block the development and transmission of pathogens by expressing effector molecules (Ratcliffe et al., 2022). Asaia spp. and Serratia spp. are the most promising candidates for paratransgenesis because of several significant features: their ability to naturally colonize mosquito tissues, their ease of cultivation under laboratory conditions, their widespread presence in mosquito populations, their potential for both vertical and horizontal transmission, and ease of genetic manipulation. These qualities make them ideal vehicles for delivering anti-parasitic or antiviral proteins (Gendrin and Christophides, 2013).
Asaia spp
Asaia spp. are naturally occurring Gram-negative, acetic acid bacteria belonging to the class Alphaproteobacteria. They were first isolated from Anopheles mosquitoes, such as Anopheles gambiae and Anopheles stephensi. Unlike Wolbachia, Asaia transmission occurs through copulation between male and female mosquitoes (Egyirifa and Akorli, 2024). Due to their strong colonization ability, Asaia species have been used as a genetically engineered microorganism to express molecules (either anti-malarial or pathogen blocking) that ultimately inhibit Plasmodium development (Cappelli et al., 2019). In a study by Favia and colleagues, stable colonization and transmission of Asaia across generations were reported using green fluorescent protein as a marker. To gain more control over Plasmodium transmission, multiple studies have developed Asaia strains that produce effector peptides targeting specific developmental stages of Plasmodium falciparum (Huang et al., 2022). Experimental studies have demonstrated that genetically engineered Asaia strains expressing anti-Plasmodium effector molecules can significantly reduce parasite development in Anopheles mosquitoes, resulting in substantial reductions in parasite load and oocyst intensity under laboratory conditions. However, quantitative estimates vary across studies, and large-scale field validation of this approach remains limited. Moreover, Asaia is compatible with sugar-bait delivery systems, allowing its deployment in the field through simple feeding bars. Despite its promise, major issues remain in understanding the Anopheles–Asaia relationship. Asaia has one major limitation; its antagonistic interaction with Wolbachia when present in the same host system, complicating co-deployment strategies. Several studies have shown that Wolbachia and Asaia abundances in the reproductive organs of many mosquito species exhibit an antagonistic relationship, particularly in anophelines. Wolbachia-infected mosquitoes had reduced Asaia densities; however, when Asaia was removed from Anopheles, Wolbachia was able to spread vertically.
Serratia spp
Serratia spp. are Gram-negative bacteria used as vector-control agents, especially for mosquito population management, because they secrete bioactive compounds that are either toxic to mosquito larvae or compete with them for nutrients in larval habitats (Gendrin and Christophides, 2013). These species produce various bioactive compounds, such as prodigiosin, a red pigment with antiviral, antifungal, and larvicidal properties, and serralysin, a protease that disrupts the midgut epithelium and may facilitate effector molecule delivery (Esteves and Scharf, 2024). Serratia spp. were first isolated from both Aedes and Anopheles mosquitoes. Studies have reported significant reductions in Plasmodium berghei oocyst formation in mosquito midguts using recombinant Serratia marcescens strains expressing molecules such as scorpine and defensins with anti-malarial activity, resulting in marked reductions in oocyst intensity (Bai et al. 2019). Furthermore, Serratia is used as a paratransgenic tool due to its ability to transmit both vertically and horizontally. It colonizes various mosquito tissues, increasing its potential as a biological tool for controlling arboviral infections. A recent study showed that the Y1 strain of S. marcescens, isolated from wild Anopheles sinensis, significantly suppressed P. berghei development by regulating immunity-related genes such as FBN9 and TEP1 (Bai et al., 2019). Moreover, CRISPR-based editing of Serratia is gaining popularity due to its potential to stably integrate effector genes with inducible promoters. Despite these advantages, several challenges are associated with paratransgenic strategies. These include: (1) the need to develop robust confinement methods, (2) risk assessment of genetically modified microorganisms, (3) the influence of resident microbiota, and (4) the potential for gene transfer to nontarget organisms (Ratcliffe et al., 2022). The regulatory framework for such microbial-mediated genetic interventions is also less clear compared to classical transgenics. Unlike transgenic mosquitoes, which require species specificity and mass rearing, paratransgenesis is more flexible because it is reversible and less invasive, allowing microorganisms to adapt to multiple mosquito species (Mancini et al., 2016).
Entomopathogenic fungi
Entomopathogenic fungi (EPF) naturally infect insects, including mosquito vectors, leading to morbidity and mortality. EPF cause death either by directly penetrating the host cuticle or by being ingested (conidia, blastospores), then colonizing the hemocoel and ultimately killing the host through nutrient depletion and toxin production (Gielen et al., 2024). Infection begins when fungal conidia adhere to the mosquito cuticle, generally on the tarsi or abdominal segments. After adhesion, the conidia germinate, penetrate the cuticle through mechanical pressure and enzymatic degradation (e.g., proteases and chitinases), and develop within the hemocoel. The fungus disrupts physiological activities and secretes secondary metabolites, such as destruxins, which inhibit the host immune response and lead to death within 4–10 days (Ma et al., 2024) (Fig. 1C).
Several studies have demonstrated the high efficacy of entomopathogenic fungi against mosquito vectors. For instance, exposure to Metarhizium anisopliae has been shown to cause up to 80–90% mortality in Anopheles mosquitoes within a few days under laboratory conditions (Scholte et al., 2005). Additionally, fungal infection significantly reduces mosquito longevity and vector competence, thereby lowering transmission potential (Blanford et al., 2005; Thomas and Read, 2007). Unlike bacterial or viral pathogens, EPF do not depend solely on ingestion; they are capable of infecting insecticide-resistant mosquito species, making them an attractive component of Integrated Vector Management (IVM). Several EPF species, including Beauveria, Lagenidium, Isaria, Tolypocladium, Aspergillus, Culicinomyces, and Coelomomyces, are effective biocontrol agents against mosquito vectors (Accoti et al., 2021). Beauveria bassiana and M. anisopliae are the two most frequently studied entomopathogens and are used in mosquito control programs because of their demonstrated efficacies against species such as A. gambiae, A. stephensi, and A. aegypti (Scholte et al., 2005).
Beauveria bassiana
B. bassiana is a soil-borne fungus belonging to the order Hypocreales that infects many insects including beetles, flies, mosquitoes, and various other arthropods. However, B. bassiana poses no threat to animals or plants, making it safe to use with no negative impact on the environment or human health. The infection process follows the same steps described above. Recent studies showed that conidia attach to the insect cuticle through hydrophobic interactions mediated by the hydrophobin genes (hyd1 and hyd2). Mutants lacking these genes exhibit reduced spore hydrophobicity and virulence (Zhang et al., 2023). B. bassiana also expresses genes such as subtilisin-like (Pr1), trypsin-like (Pr2), and cdep1, which produce cuticle-degrading proteases critical for host penetration (Dias et al., 2008). In strain Bb0062, MAP kinase genes (Bbmpk1 and Bbhog1) regulate appressorium development, which allows cuticle penetration. A 2017 study assessed the larvicidal and pupicidal effects of B. bassiana against A. stephensi, Culex quinquefasciatus, and A. Aegypti (Ragavendran et al., 2017). Application of conidia to treated indoor and outdoor surfaces, such as walls and clay pots, significantly infected adult mosquitoes after a single contact (Tawidian et al., 2023). Interestingly, B. bassiana can activate mosquito immune pathways, such as Toll, IMD, and JAK-STAT, thereby reducing the likelihood of resistance development. Field trials using autodissemination devices, in which mosquitoes carry spores to breeding sites, have also shown promise for achieving population-level effects (Tawidian et al., 2023). The demonstrated efficacy of B. bassiana in laboratory and semi-field trials supports its potential as a biological control tool.
Metarhizium anisopliae
M. anisopliae, sometimes referred to as the “green muscardine fungus,” has been widely explored as a vector control agent in the form of larvicides, pupicides, and adulticides for aquatic habitats (Bilal et al., 2012). M. anisopliae exhibits broad host specificity and thermal tolerance, making it suitable for deployment across diverse ecological zones. Unlike other EPF, M. anisopliae conidia often fail to stick to larval cuticles or germinate inside the gut, being released instead in fecal pellets. Some studies have demonstrated infection through adherence to the respiratory siphons of larvae of A. Aegypti, Culex pipiens, and A. stephensi (Scholte et al., 2005). Blastospores of M. anisopliae showed higher virulence than conidia against A. Aegypti pupae, with immediate sporulation observed after pupal exposure. Transgenic strains expressing toxins, such as scorpion-derived neurotoxins (e.g., AaIT), have been developed, increasing mosquito mortality while minimizing risks to nontarget organisms. For example, one study showed that 90% of A. gambiae were killed within 5 days of exposure to genetically modified Metarhizium (Bilgo et al., 2018). Despite their potential, deploying fungal spores in the field remains challenging, as spores must remain viable under extreme conditions such as high temperature and humidity, and persist on treated surfaces long enough to contact target mosquitoes (Qin et al., 2023). Advances in formulation science have developed oil-based and microencapsulated mediums to increase shelf life and environmental resilience. Combining fungal spores with low-dose insecticides or attractants may also improve contact rates. EPF have low environmental impact and are considered safe for humans, animals, and nontarget insects, including pollinators (Meirelles et al., 2023). Nevertheless, several limitations remain. Fungal infections often require several days, allowing mosquitoes to transmit pathogens before death (Paula et al., 2011). Another limitation to large-scale use is the need for standardized field efficacy trials to monitor their impact on disease transmission (Qin et al., 2023). Current research is exploring the integration of EPF with other biological tools, such as Wolbachia and juvenile hormone analogues (Bilgo et al., 2024). If successfully integrated, fungal biopesticides could become a central component of sustainable vector management strategies in both rural and urban environments.
Incompatible insect technique
The incompatible insect technique (IIT) is quite similar to sterile insect technique (SIT) but uses Wolbachia-induced CI in the host system, which produces nonviable offspring. IIT does not require irradiation and therefore avoids the fitness costs associated with irradiation in mosquitoes. In IIT, male mosquitoes infected with a specific Wolbachia strain are released into the wild, where they produce embryos that fail to develop due to CI when mating with uninfected females or females carrying incompatible Wolbachia strains (Pagendam et al., 2020) (Fig. 2B). Due to this convenient procedure, IIT has gained attention in various vector control programs globally, particularly in Asia, Australia, and Europe, targeting Aedes albopictus and A. aegypti (Lim et al., 2024). According to a recent study, weekly releases of Wolbachia-infected male mosquitoes in Guangzhou, China, suppressed 94% of local A. albopictus populations (Zeng et al., 2022). Moreover, combining IIT with low-dose irradiation (IIT–SIT combination) could reduce the risk of accidentally releasing Wolbachia-infected females (Bansal et al., 2024). IIT is closely linked to Wolbachia-based strategies, as both rely on CI to suppress mosquito populations. While Wolbachia-based approaches are often used for population replacement, IIT primarily focuses on population suppression through repeated male releases. In practice, these approaches are increasingly being combined with SIT to improve efficiency and reduce operational risks, particularly the unintended release of females. In addition to these methods, related genetic approaches such as release of insects carrying a dominant lethal (RIDL) have also been explored for mosquito population suppression, further expanding the range of available vector control strategies. Species specificity, nonreliance on chemical insecticides, and reduced operational expenses compared to SIT are some of the key benefits of IIT (Kittayapong et al., 2025). Moreover, some of the ecological issues related to gene drive systems are also addressed by a built-in confinement mechanism offered by the self-limiting nature of IIT (in the absence of released females) (Li et al., 2024). To implement the IIT method successfully, several factors must be considered. These include precise strain selection to ensure bidirectional incompatibility, mass production of high-quality male mosquitoes, and suitable field conditions such as temperature and humidity. Population density at the release site is also a critical factor (Aldridge et al., 2024). All these factors can significantly affect the successful implementation of IIT (CI-based suppression) for vector control. However, current studies are highly focused on developing methods based on IIT–SIT combinations and exploring new strategies for mosquito release methods in the field, considering cost-effectiveness, such as drone-based methods. At the same time, establishing the long-term epidemiological impacts of IIT on disease transmission are being predicted quantitatively by mathematical modeling (Lim et al., 2024).
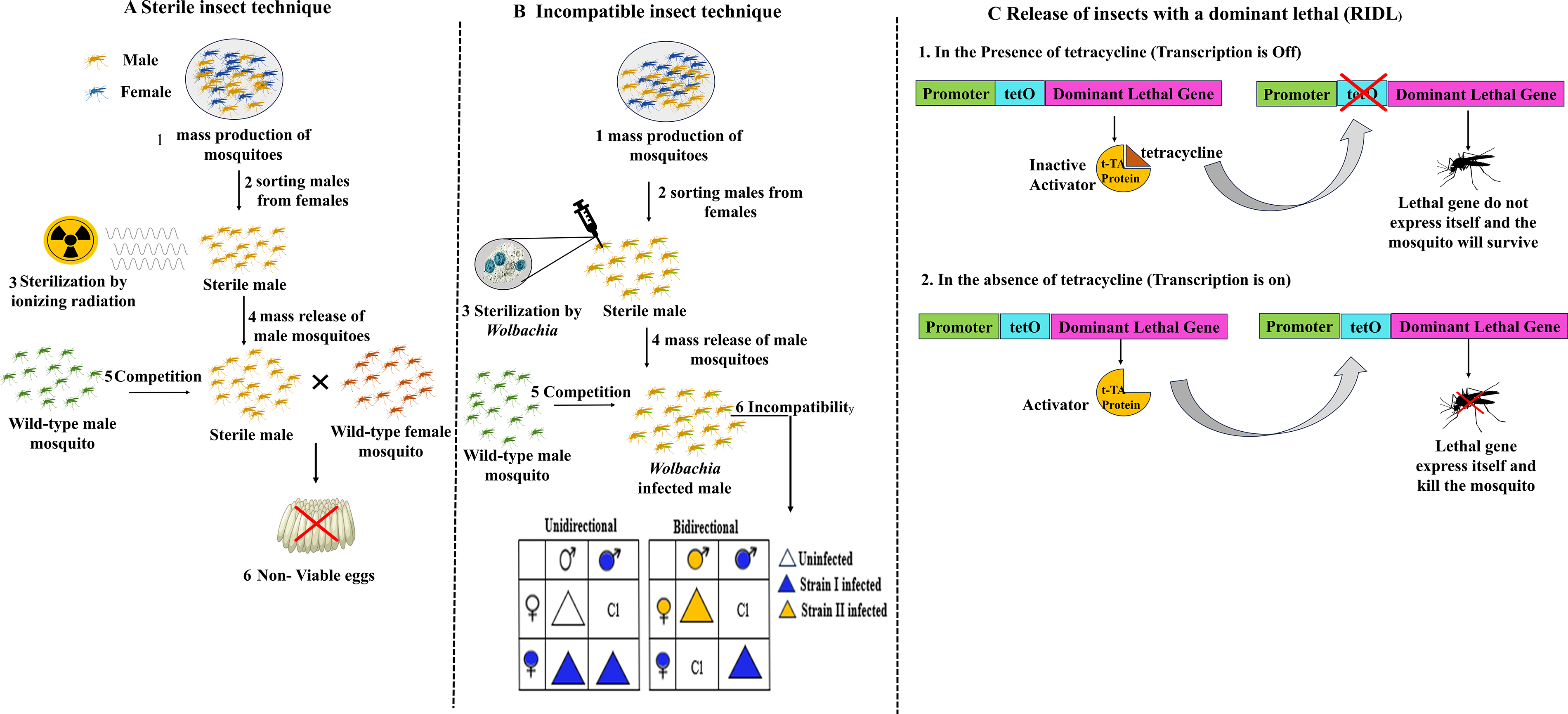
Different methods of population suppression approaches. (
Genetic Control Measures
Sterile insect technique
The SIT was originally developed in the 1950s to suppress agricultural pests, mainly the screwworm fly, and has since been established as one of the most effective methods of insect control (Lees et al., 2021). The main principle of this method involves mass-rearing target mosquitoes in a controlled environment, separating males from females, and sterilizing the males with ionizing radiation (mainly gamma rays), which damages their reproductive cells (sperm), rendering them incapable of fertilizing eggs during mating. When these sterile males are released into the wild to mate with wild females, the resulting eggs are either nonviable or have reduced viability. Ultimately, this leads to a significant reduction in the target population size over successive generations (Lees et al., 2021) (Fig. 2A). SIT is currently being utilized to control several medically important mosquito vectors, particularly species of the Aedes and Anopheles genera.
Over the past decade, SIT and related sterile male-based suppression strategies have advanced substantially from experimental frameworks to increasingly sophisticated operational deployments across multiple countries and ecological settings. In China, landmark large-scale combined IIT–SIT field trials targeting A. albopictus achieved >90% suppression of wild populations over multiple years, demonstrating robust feasibility for urban mosquito control while highlighting the advantages of integrating Wolbachia-based incompatibility with conventional sterilization (Zheng et al., 2019). In Italy, sustained releases of sterile A. albopictus males significantly reduced egg hatch rates and local mosquito population density, further validating SIT under European field conditions (Bellini et al., 2013). In Brazil, genetically based sterile male suppression strategies targeting A. aegypti, including transgenic and sterile male–based interventions, achieved local population reductions exceeding 80% under semi-operational conditions, reinforcing the applicability of sterile male–based mosquito control in endemic tropical regions (Macedo et al., 2026). In Greece, pilot sterile male release programs against A. albopictus further demonstrated the ecological adaptability of SIT under Mediterranean environmental conditions (Balatsos et al., 2024). In the United States, field deployment of self-limiting A. aegypti strains in Florida provided additional evidence for the translational potential of sterile male–based mosquito suppression technologies within highly regulated operational frameworks (Morreale et al., 2025). Together, these deployments across China, Italy, Brazil, Greece, and the United States demonstrate that SIT has evolved beyond its classical irradiation-based foundation into a broader and increasingly field-validated mosquito suppression platform. Recent innovations, particularly combined IIT–SIT frameworks, further enhance suppression efficiency, reduce risks associated with female contamination, and partially offset fitness costs linked to irradiation alone (Zheng et al., 2019; Bansal et al., 2024). Despite these advances, broader implementation remains constrained by major operational barriers, including cost-intensive mass rearing, precise sex separation, maintenance of sterile male mating competitiveness, regulatory oversight, and the substantial infrastructure required for sustained area-wide deployment (Lees et al., 2021). To address these limitations, innovations such as genetic sexing strains, automated sex-sorting technologies, and optimized artificial diets are increasingly being developed to improve scalability, cost-effectiveness, and long-term implementation feasibility (Lees et al., 2021). Moreover, the Food and Agriculture Organization of the United Nations (FAO) and the International Atomic Energy Agency (IAEA) has actively promoted SIT as part of integrated vector management programs in multiple countries, reinforcing its growing institutional relevance in sustainable mosquito control. Despite these successes, SIT has limitations, including difficulties in large-scale mass rearing, incomplete sex separation, potential reductions in mating competitiveness of irradiated males, and high operational costs (Lees et al., 2021). Advances in automation, artificial diets, and genetic sexing strains are currently being explored to overcome these barriers.
Release of insects carrying a dominant lethal
RIDL is a transgenic mosquito suppression strategy that employs genetically engineered male mosquitoes carrying a dominant, self-limiting lethal gene designed to suppress wild vector populations through inherited offspring mortality. RIDL mosquitoes are typically reared under laboratory conditions supplemented with tetracycline, which suppresses lethal gene expression and permits colony maintenance. The RIDL mechanism and population suppression strategy are illustrated in Fig. 2C. Once released into natural environments, engineered RIDL males mate with wild females; however, in the absence of tetracycline, the lethal transgene is activated in progeny, causing death before reproductive maturity and thereby progressively reducing local mosquito population density over successive generations (Alphey et al., 2010; 2014). Unlike classical SIT, RIDL does not depend on irradiation, potentially preserving male mating competitiveness while maintaining species-specific suppression.
Over the past decade, RIDL has become one of the most operationally advanced transgenic mosquito control strategies, particularly against A. aegypti. Early open-field trials in the Cayman Islands using Oxitec’s OX513A strain demonstrated ∼80% suppression of local A. aegypti populations (Harris et al., 2012). Subsequent sustained releases in Juazeiro, Brazil, reported population suppression frequently exceeding 80–95%, providing robust large-scale field validation under endemic tropical conditions (Carvalho et al., 2015). Additional operational deployments in Panama further supported RIDL’s ecological adaptability across geographically distinct transmission settings (Gorman et al., 2016). These multinational studies collectively established RIDL as a major proof-of-concept for scalable, genetically based mosquito suppression. RIDL is considered a self-limiting strategy because its effects depend on repeated releases of engineered males rather than permanent spread through wild populations. This feature may provide greater control compared with gene drive systems, since suppression can be adjusted or stopped if needed; however, it also means that long-term success depends on continuous operational investment. RIDL offers several advantages, including species-specific targeting, reduced reliance on chemical insecticides, and potential integration with other mosquito suppression approaches such as SIT or IIT. At the same time, important challenges remain. These include the need for repeated large-scale releases, high production and implementation costs, regulatory and biosafety requirements, and ongoing public concerns regarding the release of genetically modified mosquitoes. In many low-resource settings, these economic, infrastructural, and social barriers may limit large-scale sustainability. Therefore, although RIDL has become one of the most operationally advanced transgenic mosquito suppression strategies currently available, its wider adoption will depend not only on biological effectiveness but also on affordability, regulatory acceptance, and public trust. While RIDL provides a self-limiting transgenic suppression strategy, gene drive systems represent a more self-sustaining genetic approach aimed at spreading engineered traits through wild populations.
Gene drives
One of the most powerful and highly controversial genetic technologies for vector control is the gene drive. This method has a mechanism of inheritance that completely contrasts with the conventional Mendelian process. Gene drives increase the frequency of a gene within a population so rapidly that it facilitates the spread of the gene throughout the target population (Fig. 3). This technology is being explored to modify or control vector populations either by decreasing their competence to carry disease-causing agents or by altering their reproductive systems in ways that lead to infertility or poor survival (Hefferon and Herring, 2020). Homing Endonuclease Genes (HEGs), Zinc Finger Nucleases (ZFNs), Transcription Activator-Like Effector Nucleases (TALENs), and the CRISPR–Cas9 system are a few examples of gene drive mechanisms that have been extensively studied (Fig. 3). Among these, CRISPR-based gene drives have been especially attractive and have captured the most attention due to their ease of use, effectiveness, and programmable capability (Costa et al., 2017).
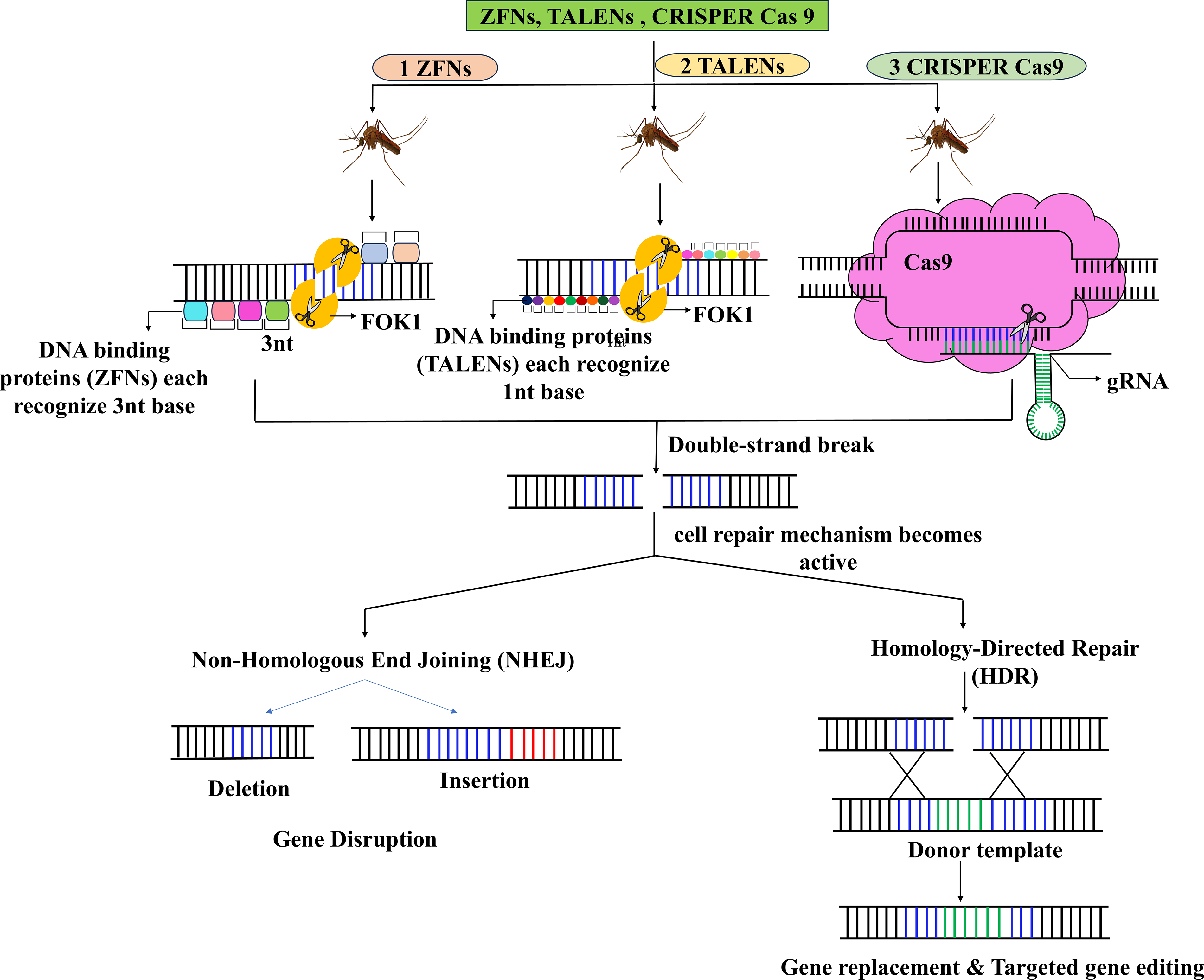
Different methods of population modification approaches using gene drive.
One of the most powerful and controversial genetic technologies for vector control is the gene drive (Burt, 2003; Hefferon and Herring, 2020). Unlike conventional Mendelian inheritance, gene drives bias gene transmission, allowing specific traits to spread rapidly through target populations (Fig. 3). These systems are being explored either to suppress mosquito populations by disrupting fertility or survival or to modify them by reducing pathogen transmission capacity (Hefferon and Herring, 2020). Several platforms, including HEGs, ZFNs, TALENs, and especially CRISPR–Cas9, have been investigated, with CRISPR-based systems attracting the greatest attention because of their efficiency and programmability (Costa et al., 2017). Experimental studies in A. gambiae have demonstrated remarkable suppression potential under controlled settings, including near-complete population collapse in caged populations (Kyrou et al., 2018). However, despite their transformative promise, gene drives remain largely experimental and have not yet reached broad field deployment comparable to SIT, IIT, or RIDL. Key concerns include resistance evolution, ecological uncertainty, unintended spread beyond target regions, and significant ethical and regulatory challenges (National Academies of Sciences, Engineering, and Medicine, 2016). Therefore, although gene drives represent a potentially powerful future tool for mosquito-borne disease control, their practical implementation currently remains limited by substantial biosafety, governance, and societal concerns.
Homing endonuclease genes
HEGs are naturally occurring autonomous genetic elements that can identify and cleave specific DNA sequences. Because of this ability, HEGs can cleave the homologous wild-type allele upon their introduction into mosquitoes. Later, this gene drive sequence can be replicated at the cleaved site through the natural replication process of mosquitoes in general, and by homology-directed repair in particular. As a result, nearly all offspring acquire this gene drive rather than the expected 50% (Oberhofer et al., 2018). Previously, HEG-based gene drives targeted genes involved in fertilization or sex determination. For example, HEGs directed against the dsx gene in A. gambiae led to female sterility and progressive population collapse (Bier, 2022). However, some limitations exist with HEGs, such as the requirement for strict sequence recognition and lower flexibility compared to CRISPR.
Zinc finger nucleases and transcription activator-like effector nucleases
In ZFNs and TALENs, specific engineered proteins are required to recognize and cleave DNA; therefore, their design and validation are required for each target site. The literature shows their use in mosquitoes for vector control; however, their application has been limited due to poor efficiency and highly complex protein designs (Gaj et al., 2013).
CRISPR–Cas9-mediated gene drive
CRISPR–Cas9 gene drives have become one of the most popular platforms for genetic control due to their versatility, which allows them to target almost any gene. Typically, two elements, the Cas9 enzyme, which functions like molecular scissors, and guide RNA (gRNA), which contains the genetic information required to target a DNA sequence, are inserted into the mosquito genome in the CRISPR-based drive system to make modifications. When these modified mosquitoes mate with unmodified (wild-type) ones, the Cas9/gRNA complex cleaves the targeted region in the gene on the wild-type chromosome. The drive-carrying chromosome then serves as a template for repair. The CRISPR–Cas9 system is sometimes referred to as a highly developed biological “search/replace” mechanism that cuts DNA and replaces the old code with a new one using gRNAs. This process of copying the drive segment is called “homing,” and it ensures that nearly 100% of offspring inherit the drive (Kistler et al., 2015). Various studies have demonstrated successful results of vector control in the laboratory using this system. For example, knocking out the fibrinogen-related protein 1 (FREP1) gene from A. gambiae mosquitoes using the CRISPR–Cas9 system may develop resistance to Plasmodium. In another study conducted in 2018, Hammond et al. reported intersex phenotypes or 100% sterilization in females of A. gambiae when the sex-differentiation controlling gene double-sex was targeted. Other successful studies on A. gambiae have also been documented, where the drive spread through a caged population within only 10 generations, leading to total population collapse (Kistler et al., 2015). Similar studies have been reported in A. aegypti to spread anti-pathogen effectors or female-specific lethal genes. Despite several promising features such as potential and specificity, gene drives are associated with significant ecological, ethical, and regulatory challenges: (1) mutations at the target site can hamper cleavage and lead to resistant alleles, ultimately blocking drive spread, (2) cleavage at nontargeted sites might result in unanticipated ecological or physiological consequences, (3) difficulty or irreversibility in recalling the drive once released, especially in highly mobile species, and (4) nontarget spread across species or unintended transport to other areas, which may threaten ecological balance (Collins, 2018). To overcome these concerns, various strategies are being explored. These include the use of molecular safeguards such as split drives, which separate Cas9 and gRNA components, and “threshold-dependent” or “daisy-chain” drives, which restrict persistence. Moreover, field trials are being designed with rigorous confinement, stakeholder participation, and phased testing frameworks. Furthermore, organizations such as WHO, the Convention on Biological Diversity, and national biosafety agencies are establishing international rules for gene drive research and deployment (Kelsey et al., 2020). However, no information is available about the release of CRISPR-based gene drives into open environments. Target Malaria, a nonprofit consortium developing gene drive technology, is pursuing laboratory research followed by limited field trials on A. gambiae in Africa and represents one of the most well-known gene drive initiatives. With rapid advancements, strong funding support, community engagement, transparent disclosure of risks, and cross-sector collaboration, CRISPR-based gene drives may eventually become feasible to implement (Kelsey et al., 2020).
Discussion
In this review, we attempted to cover past and ongoing research on emerging biological and genetic strategies to fight against mosquito-borne diseases. These approaches, ranging from symbiont-mediated pathogen prevention to sophisticated gene-editing technologies, offer both new opportunities for change and enormous challenges. There is a need to assess their impact on public health and ecology, their potential integration with ongoing vector control programs, and the ethical and legal frameworks defining their implementation (van den Berg et al., 2021). Conventional vector control strategies have mainly relied on chemical insecticides and environmental control methods. However, a shift toward holistic and sustainable methods is required because of the continuous development of insecticide resistance among mosquito species and the reduced effectiveness of pesticides. Aligning recently developed biological and genetic strategies with the WHO’s IVM principles may provide better alternatives to conventional vector control strategies (van den Berg et al., 2021). For example, in urban settings, especially where Aedes mosquitoes have very high densities, Wolbachia-based population replacement strategies can be successfully applied without changing habitats or using chemical spraying (Utarini et al., 2021). In peri-urban or rural settings with lower mosquito densities, SIT and IIT approaches are a perfect fit, with repetitive releases of male mosquitoes (Carvalho et al., 2015; Zeng et al., 2022). Furthermore, entomopathogenic fungi can be integrated with IRS and larval habitat management, while paratransgenic tools may provide various delivery platforms targeting different vector species (Gielen et al., 2024). With proper strategy, integration of these novel approaches with conventional methods could become a game-changer in controlling various VBDs. Mathematical models also support these strategies. For example, when SIT is combined with Wolbachia or gene drives with Attractive Toxic Sugar Baits (ATSBs), results show outcomes greater than the sum of applying these methods individually. Among all, Wolbachia-based vector control interventions, in which Wolbachia-infected mosquitoes were released in the field in countries like Australia, Indonesia, and Brazil, have shown significant reductions in dengue incidence (Utarini et al. 2021). Similar results were reported in pilot SIT and IIT programs, which showed significant suppression of Aedes and Anopheles mosquito populations (Zheng et al., 2019). Although these results provide initial success in controlling VBDs, real-world implementation faces various challenges. These include technical and financial barriers, the requirement for large infrastructure for mass mosquito production, quality control for irradiation or infection, and extensive community outreach (Benedict and Robinson, 2003). Furthermore, although gene drive systems are still in the experimental phase, they have the potential to be transformative for vector control. With field trials confirming laboratory success, this approach could provide solutions that are cost-effective and scalable (Burt, 2014). To successfully implement SIT, IIT, or paratransgenesis in the field requires extraordinary facilities, such as highly specialized infrastructure, systems for mass mosquito production, irradiation, sex-sorting technologies, and quality control protocols. These remain barriers, especially in rural and resource-limited settings (Lees et al., 2021). Moreover, accurate assessment of intervention outcomes becomes crucial when establishing these methods is already highly challenging. Therefore, reliable measures of vector competence, infection prevalence, and behavioral adaptations in natural settings are required. However, technical challenges remain due to a lack of standardized markers, reliable entomological surveillance systems, and real-time epidemiological data. Additionally, insufficient data on the long-term durability of interventions based on microbial symbionts or gene drives increases these challenges.
While these strategies show considerable potential, their effectiveness often depends on ecological context, scalability, and how easily they can be implemented in real-world settings. For example, Wolbachia-based approaches and SIT/IIT have already demonstrated strong field performance and practical applicability, whereas gene drive and paratransgenic strategies are still largely at the experimental stage and require further evaluation, particularly in terms of ecological safety. In addition, factors such as cost, infrastructure, and regulatory challenges can significantly influence their large-scale deployment. Therefore, no single strategy is universally optimal, and integrated, context-specific approaches are likely to be more effective. Ultimately, balancing efficacy, safety, and sustainability will be essential for the successful implementation of next-generation vector control tools.
Collectively, these emerging mosquito control strategies differ substantially in technological maturity, operational readiness, scalability, and biosafety complexity. Approaches such as SIT, IIT, RIDL, and Wolbachia-based interventions currently demonstrate the strongest field validation and near-term deployment potential, particularly when integrated into existing vector management frameworks. In contrast, gene drives and advanced paratransgenic systems offer transformative long-term possibilities but remain comparatively constrained by ecological uncertainty, regulatory complexity, and limited field validation. Consequently, the future of mosquito control will likely depend less on any single universal strategy and more on context-specific integration of complementary biological, genetic, and ecological interventions.
The use of these emerging genetic technologies is also heavily influenced by ethical concerns related to transparency and ecological risks, which must be addressed before field implementation (James et al., 2018). Since gene drives and paratransgenic approaches introduce inheritable changes into wild populations or their microbiomes, unlike chemical or mechanical approaches, there are concerns about global spread, biodiversity effects, nontarget species, and unforeseen consequences. As regulatory frameworks vary between countries, establishing unified regulatory standards is difficult, alongside the need for meaningful community engagement (Lavery et al., 2010). Although in agriculture, biopesticides such as fungi already have well-established regulatory channels, similar systems are needed for genetically modified organisms (GMOs). Some initiatives, like the WHO Guidance Framework for Testing Genetically Modified Mosquitoes, the Cartagena Protocol on Biosafety, and national biosafety laws, provide partial supervision. However, uniform global guidelines are still required. A comparative overview of major vector control strategies is presented in (Table 1) to highlight their effectiveness, operational feasibility, and current research gaps.
Comparative Overview Of Major Biological And Genetic Vector Control Strategies For Mosquito-Borne Diseases, Summarizing Their Effectiveness, Field Validation Status, Advantages, Limitations, And Key Research Gaps. Information Is Derived From Representative Experimental And Field Studies Cited In The Text

GMO, genetically modified organism.
Conclusions
Mosquito-borne diseases continue to pose a threat to global health, particularly in tropical and subtropical regions, which have limitations in the effectiveness, accessibility, and sustainability of vector control programs. Various interventions, including species-specific, environmentally friendly, and long-lasting mosquito control strategies, are emerging and have demonstrated encouraging outcomes, supported by several successful field trials, particularly for Wolbachia-based interventions and the SIT. In contrast, gene drive strategies remain largely in the experimental phase and require further validation through field studies and implementation frameworks. Despite these advances, implementation remains challenging due to factors such as counter-resistance, regulatory and operational constraints, scalability issues, ecological uncertainties, and the need for effective community engagement. These challenges may be mitigated through coordinated policy frameworks, interdisciplinary collaboration, and structured stakeholder engagement. Moreover, a proper strategy will be required in the future to integrate these newly developed approaches into IVM. Evidence-based deployment models, supported by adequate infrastructure and strong community partnerships, could facilitate the integration of these approaches into IVM. In conclusion, biological and genetic control methods, as potential alternatives to conventional methods, represent an important advance in vector management. Among currently available strategies, Wolbachia-based systems, SIT, IIT, and RIDL demonstrate the strongest operational maturity and field validation, whereas gene drives and advanced paratransgenic approaches remain more experimental despite substantial long-term transformative potential. Therefore, future progress in mosquito-borne disease control will likely depend on context-specific integration of complementary biological, genetic, and ecological strategies rather than reliance on any single universal intervention. With appropriate resourcing and planning, including risk assessments for integrating these methods, we can reduce the burden of mosquito-borne diseases and take one step closer to achieving health equity and global disease eradication.
Authors’ Contributions
N.K.: Conceptualization, writing, initial and final manuscript, visualization, and supervision (lead). M.J.: Writing—initial manuscript and figure preparation. N.K. and A.D.: Writing—reading and editing the final manuscript.
Footnotes
Author Disclosure Statement
No conflicting financial interests exist.
Funding Information
The authors received no financial support for this research, authorship, and/or publication of this article.